Article Text

Abstract
Objective Variants in CLEC16A have conferred susceptibility to autoimmune diseases in genome-wide association studies. The present work aimed to investigate the locus' involvements in juvenile idiopathic arthritis (JIA) and further explore the association with rheumatoid arthritis (RA), type 1 diabetes (T1D) and Addison's disease (AD) in the Norwegian population.
Methods Three single nucleotide polymorphisms (SNPs) were genotyped in patients with RA (n=809), JIA (n=509), T1D (n=1211) and AD (n=414) and in healthy controls (n=2149).
Results All diseases were associated with CLEC16A, but with different SNPs. The intron 22 SNP, rs6498169, was associated with RA (p=0.006) and JIA (p=0.016) and the intron 19 SNPs, rs12708716/rs12917716, with T1D (p=1×10−5) and AD (p=2×10−4). The RA association was confined to the anti-cyclic citrullinated peptide antibody (anti-CCP) negative subgroup (p=2×10−4).
Conclusion This is the first report of a CLEC16A association with JIA and a split of the RA association according to anti-CCP status. Different causative variants underlie the rheumatic versus the organ specific diseases.
This paper is freely available online under the BMJ Journals unlocked scheme, see http://ard.bmj.com/info/unlocked.dtl
Statistics from Altmetric.com
Introduction
Genome-wide association studies (GWAS) have recently identified single nucleotide polymorphisms (SNPs) in C-type lectin domain family 16, member A (CLEC16A) to be associated with the autoimmune diseases; type 1 diabetes (T1D)1 2 and multiple sclerosis (MS).3 To date, the function of CLEC16A remains unknown. A role in immunity is however likely since it is almost exclusively expressed in immune cells, such as dendritic cells, B lymphocytes and natural killer cells (http://symatlas.gnf.org/SymAtlas/). The gene is classified as a C-type lectin based on bioinformatic analyses, although atypical, as it lacks crucial domains in carbohydrate recognition.4
We have earlier reported a positive association between polymorphisms in CLEC16A and Addison's disease (AD),5 and such evidence was also recently reported in rheumatoid arthritis (RA).6 Different polymorphisms in CLEC16A have been reported to be disease associated in different autoimmune diseases, with a possible explanation that studies have not analysed the same SNP sets. However, there is extensive linkage disequilibrium (LD) between the most strongly associated SNPs, and a representative subset of these SNPs was analysed in this study to attempt to ascertain if these associations could point to a common SNP, or if the associations in fact rely on different SNPs.
Our aim was to provide further support for CLEC16A as an autoimmune risk locus and in particular to address the potential role in susceptibility to juvenile idiopathic arthritis (JIA), a disease not previously studied in this context, as well as to further explore its putative role in RA.
Patients and methods
Patients and controls
The panel of autoimmune diseases consisted of 809 patients with RA, 509 with JIA, 1211 with T1D and 414 with AD (see supplementary material). Controls were recruited from 2 independent cohorts: 1029 from set 1 and 1120 from set 2. All individuals were Norwegian Caucasians and informed consent was given by all participants in compliance with the Helsinki Declaration.
SNP selection
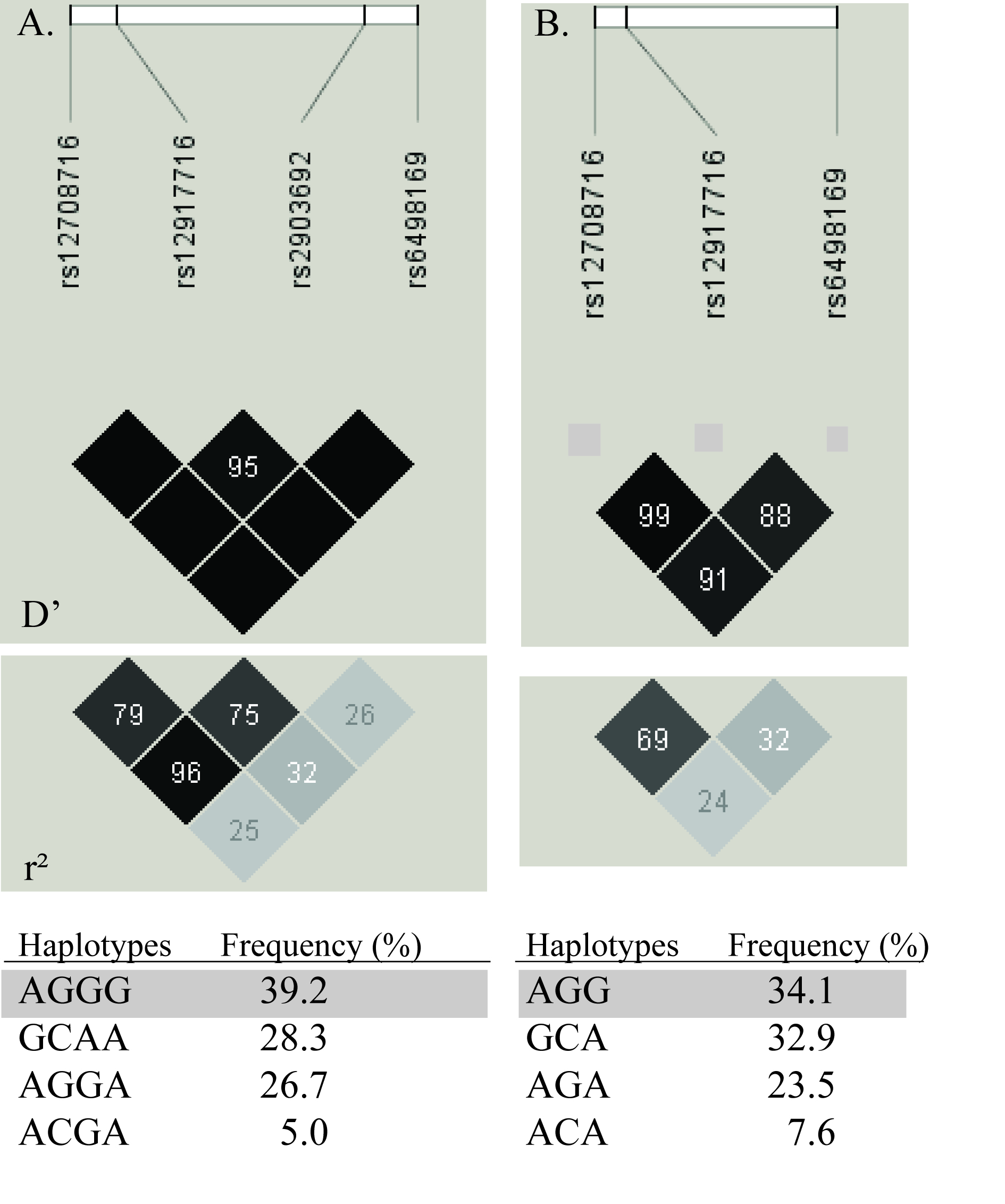
First, based on the dense tagging of SNPs analysed in our previous study on AD,5 we decided to genotype the strongest associated SNP, rs12917716, in the other disease sample sets and an enlarged AD sample set. Second, based on the LD pattern and haplotypes between the SNPs that have shown association with other autoimmune diseases (supplementary figure 1A),1,–,3 together with the knowledge that RA and T1D have shown association with different SNPs in CLEC16A,6 we also genotyped rs12708716 and rs6498169 in the RA, JIA and T1D samples. Rs12708716 was earlier analysed in 332 AD samples and rs6498169 was then tagged by rs27838 (r2=0.97).5
Genotyping
Genotyping of rs12917716, rs12708716 and rs6498169 was performed using predesigned TaqMan SNP Genotyping Assays (Applied Biosystems, Foster City, California, USA), except rs12917716, rs12708716 and rs27838 in the previously published AD samples and control set 1,5 which were analysed with SNPlex Genotyping System (Applied Biosystems).
Serology
An ELISA kit assay (INOVA Diagnostics, San Diego, California, USA) was used to measure anti-cyclic citrullinated peptide antibody (anti-CCP) concentrations in the RA samples, with a positivity cut-off defined as levels >25 U/ml.
Statistical analyses
Power calculations (α=0.05) were performed using R V.2.6.1,7 assuming an OR of 1.25 and an allele frequency of 40%; fairly representative for the three analysed SNPs. Estimated power was for RA 97%, JIA 89%, T1D 99% and AD 83%. Allele and genotype analyses were performed in Plink V.1.058 based on χ2 contingency tables. Reported p values are two sided and not adjusted for multiple testing because of the strong correlation between the genotyped SNPs. Haplotype constructions and regression analyses to estimate haplotype risks were performed using Unphased V.3.1.1,9 choosing the haplotype formed by the protective alleles as baseline reference.
Results
All four autoimmune diseases showed associations with SNPs in CLEC16A (table 1). RA and JIA were significantly associated with the intron 22 SNP, rs6498169. In contrast, T1D and AD proved to be strongly associated with the two intron 19 SNPs, rs12708716 and rs12917716. There was strong LD between rs12708716 and rs12917716 in the Norwegian population with D′=0.99 and r2=0.69 (supplementary figure 1B). Both were also in partial LD with rs6498169 with D′>0.88 and r2>0.24. The associations in RA, T1D and AD fitted an additive model, while the significance in the JIA cohort was highest in a recessive model (p=0.003).
Association analyses of single nucleotide polymorphisms (SNPs) in the CLEC16A gene
These findings raised the question whether there appear to be different causal variants in RA and JIA compared with T1D and AD. Hence we investigated haplotypes to explore if different risk haplotypes emerged, or if one common haplotype could explain these different association patterns (supplementary table 1). When constructing haplotypes of rs12917716–rs6498169, the rs6498169 risk allele G appeared only on haplotype IV, which was significantly associated with RA, JIA and T1D. The risk for this haplotype was more or less the same as the single SNP analysis of rs6498196 in JIA and RA, concluding that this SNP alone mark their association within the CLEC16A locus and that adding the T1D/AD associated SNP rs12917716 in a haplotype did not result in a better marker for the RA and JIA association.
Haplotype construction in T1D revealed that haplotypes III and IV with different rs6498196 allele had similar risk estimates, suggesting that this SNP cannot explain the T1D association, in contrast with what was observed in RA and JIA. AD was in line with T1D, suggesting that the rs12708716–rs12917716 risk haplotypes (III and IV) pointed to an untyped SNP, in strong LD with both, to be the causative variant.
The anti-CCP status has been shown to divide patients with RA into two groups clinically and genetically and we therefore considered it relevant to stratify the patients with RA based on anti-CCP status. This revealed that the association with rs6498196 followed the patients who were anti-CCP negative as allele G was significantly increased among patients who were anti-CCP negative (44.0%) compared to patients who were anti-CCP positive (37.7%) or controls (35.9%), OR=1.40 (95% CI 1.18 to 1.68), p=2×10−4 (table 2). There was no evidence of significantly different allele frequencies among JIA subtypes (p=0.2) (supplementary table 2).
Association of rs6498169 stratified on anti-CCP status in patients with RA
Discussion
CLEC16A polymorphisms were initially detected as susceptibility markers for T1D and MS in GWAS;1,–,3 associations, which have thereafter shown their resemblance to other populations,4 6 10,–,13 now also with T1D in Norwegians. Our results confirm the recently reported association with RA in a Spanish population,6 but demonstrated that this association is restricted to patients who were anti-CCP negative. Further, an association with JIA was demonstrated for the first time.
Interestingly, our results indicate that different causative SNPs underlie the systemic autoimmune diseases, RA and JIA and the organ specific diseases, T1D and AD, included in this study. The rheumatic disorders RA and JIA are caused by inflammation of the joints, even though their clinical features differ significantly. However, T1D and AD are anticipated to share some genetic background as almost 10% of the patients with AD have concomitant T1D. Studies in MS are inconclusive regarding which SNP in CLEC16A is the most strongly associated. Rs12708716 did not initially pass the quality check in the GWA reporting on rs6498169 as the strongest association,3 but has subsequently been reported with equivalent risk estimates in the same sample set.14 The other initially reported T1D SNP, rs2903692,1 is located in the same intron as rs6498169, but in stronger LD with rs12708716 than rs6498169 (supplementary figure 1A). Also the risk estimates for rs2903692 were indistinguishable from the rs6498169 risk in the MS sample reported by Martínez et al.6 However, rs2903692, and not rs6498169, showed evidence of association in a subgroup of patients with Crohn's disease.15 Rs2903692 was not genotyped in our study due to the strong correlation with rs12708716 and rs12917716.
The identification of clinical subsets of heterogeneous diseases is important for the understanding of their genetic basis. This has been clearly demonstrated in RA where different human leucocyte antigen (HLA) DRB1 associations in the context of anti-CCP antibody status have been reported; DRB1 shared epitope (SE) association being restricted to patients who were anti-CCP positive, and the DRB1*03 haplotype associated with RA solely in patients who were anti-CCP negative.16 Also non-HLA associations have been dependent on subclassification of anti-CCP, for example, CTLA4 and peptidyl arginine deiminase type IV (PADI4) are associated with patients who are anti-CCP positive.17 Conversely, the CLEC16A association with RA in the present study was restricted to the anti-CCP negative subgroup, as has also been reported for interferon regulatory factor 5 (IRF5).18 It should be noted that Martínez et al did not detect any difference in CLEC16A association when stratifying for anti-CCP, but their statistical power was also lower.6
Our study highly supports variants within CLEC16A to tag a common autoimmunity predisposing locus. Further effort is needed to understand the function of CLEC16A and the splicing variants within this LD block, and also to identify the causative variants, which seem to differ between the autoimmune diseases.
Acknowledgments
The Norwegian Bone Marrow Donor Registry is acknowledged for contributing DNA from healthy controls. The authors thank Anne-Marit Selvaag for collecting JIA samples and Gry Namløs Nordang for excellent administration of the RA sample set (Oslo University Hospital, Rikshospitalet).
References
Supplementary materials
Web Only Data ard.2009.114934
Files in this Data Supplement:
{kind=link}
Footnotes
-
Funding This research was supported by grants from EU FP7, Grant number 201167, Euradrenal and the South-Eastern Regional Health Authorities.
-
Competing interests J WJ Bijlsma was the handling editor for this manuscript.
-
Ethics approval This study was conducted with the approval of the Regional Ethical Committees of Western and South Eastern Norway and the Data inspectorate of Norway.
-
Provenance and peer review Not commissioned; externally peer reviewed.