Article Text

Abstract
Background: Variants in the growth differentiation factor 5 (GDF5) and in the double von Willebrand factor A (DVWA) have recently been reported to be associated with osteoarthritis (OA) in Asian populations.
Objective: To assess the role of such variants in OA susceptibility in two independent UK samples of Caucasian origin.
Methods: Polymorphisms rs11718863 and rs7639618 (DVWA) and rs143383 (GDF5) were genotyped in 999 patients with knee OA, 843 patients with hip OA and 1166 controls from two UK studies from Nottingham and Chingford.
Results: In agreement with previous reports, the major allele at rs143383 (GDF5) was associated with a higher risk of knee OA in our samples (ORMH = 1.29, 95% CI 1.14 to 1.47 p = 8×10−5). Conversely, the major allele at the DVWA SNP rs7639618, which increased risk in Asians, was not associated with a risk of knee OA, (ORMH = 0.88, 95% CI 0.74 to 1.04; p = 0.12). A meta-analysis of Asian and UK knee OA data indicated highly significant heterogeneity (I2 = 92%, Q = 48.5, p = 7×10−10) and no significant association with knee OA using a random effects meta-analysis (ORDL = 1.18, 95% CI 0.86 to 1.63; p = 0.309).
Conclusions: These data confirm that the GDF5 variant is consistently associated with the risk of knee OA. Considerable ethnic variation in allele frequencies at the DVWA gene was found and no significant association was found in UK samples or by combining UK and Asian samples. The results suggest that the effect of DVWA amino acid changes on tubulin binding is unlikely to influence the risk of OA in Caucasians.
Statistics from Altmetric.com
Osteoarthritis (OA) is the most prevalent form of arthritis in the elderly. Primary OA is an idiopathic phenomenon, occurring in previously intact joints, with no apparent initiating factor such as joint injury or developmental abnormalities. Several lines of work have shown a role of genetics in OA including familial aggregation studies,1 2 twin studies, linkage analysis and genetic association studies.3 4
Recent studies in Japanese and Chinese populations have reported associations of various loci with knee or hip OA. The growth and differentiation factor 5 (GDF5) is known to be involved in the development and maintenance of bone and cartilage.5 In addition, genetic variation at this locus has been shown to influence height in humans.6 The common, T allele of the rs143383 single nucleotide polymorphism (SNP) is associated with higher risk of knee and hip OA in Asian populations (OR = 1.55, p<1 ×10−6), and luciferase reporter assays have shown that the same allele mediates a significant reduction in the activity of the GDF5 promoter.7 Replication in European samples of the GDF5 association showed a weaker effect of this variant than the one reported in Asians, statistically significant for knee OA (OR = 1.13) but weaker for hip OA (OR = 1.04, NS).8 However, other variants, such as an intronic polymorphism in the CALM1 gene which showed a strong association in the Japanese population,9 have been found not to have any effect on risk of OA in European studies.10 11
More recently a locus which had until then been considered only a hypothetical gene (LOC344875 gene ID = 344875) theoretically encoding a 1504 amino acid protein was found to be a real gene which actually encodes a 276 amino acid peptide and it was called by the authors double von Willebrand factor A (DVWA).12 The common variants at two non-synonymous polymorphisms in the DVWA gene, rs11718863 and rs7639618, were reported in Japanese and Chinese patients to be very strongly associated with risk of knee OA (OR = 1.43 p<7 ×10−11) and to influence the binding of the DVWA protein to β-tubulin. The authors concluded that the weaker tubulin binding by the wild-type protein was the likely molecular mechanism by which genetic variation in this gene affected the risk of knee OA and that therefore tubulins and microtubules might be protective factors in the pathogenesis of OA.
In this study we investigated if the above-mentioned genetic variants were associated with OA of the knee and the hip in two independent United Kingdom studies.
Subjects and methods
Study subjects
Nottingham case-control study
Patients with knee or hip OA were recruited in Nottingham both from families with a history of OA and from clinic populations. All research participants gave written informed consent to take part. Approval for recruitment of index patients with knee and hip OA and siblings of these patients was obtained from the research ethics committees of Nottingham City Hospital and North Nottinghamshire.
Patients with hip and knee OA were recruited from hospital orthopaedic surgery lists (current and for the previous 5 years). All had been referred to hospital with symptomatic, clinically severe hip or knee OA and the majority had undergone unilateral or bilateral total hip replacement (THR) or total knee replacement (TKR) within the previous 5 years. Subjects were excluded from the study if they had other major arthropathy (eg, rheumatoid arthritis, ankylosing spondylitis); Paget’s disease of bone affecting the pelvis or femur; overt childhood hip disease (eg, Legg–Calvé–Perthes disease, slipped femoral epiphysis, severe acetabular dysplasia); THR due to trauma or avascular necrosis of the femoral head; or terminal illness. We only included patients if they had a diagnostic code of primary hip OA, and we excluded patients aged >90 or those who had had surgery at <40 years of age. Patients were further characterised by inquiry, examination and investigation. Height and weight were measured to calculate the body mass index (BMI). Controls were age-matched subjects from the same hospital catchment area. Unaffected siblings of probands who had had a joint replacement, free from radiographic OA and over the age of 55 were also used as controls. At most one affected and one unaffected sibling per family were included and before including siblings in the genetic association analyses we confirmed that there were no differences in allele frequencies between unrelated controls and sibling controls. For the GDF5 SNP the minor allele frequencies (MAFs) were 37.1% in unaffected siblings and 38.4% in unrelated controls, χ2 (1 df) = 0.23, p<0.63 NS. For the DVWA SNPs the MAF in unrelated controls was 14.9% and 15.4% in sibling controls (χ2 (1 df) = 0.05 p<0.82). Hence we concluded that allele frequencies at the polymorphisms studied in unaffected siblings were equivalent to those in unrelated controls.
Preoperative knee or pelvis radiographs of index patients with knee or hip OA were examined to confirm the diagnosis and to grade for changes of OA. For siblings of index patients with hip OA, new anteroposterior pelvis radiographs were obtained. Siblings of patients with knee OA were assessed for radiographic knee OA. All pelvis and knee radiographs were scored for features of OA by a single observer using the Kellgren and Lawrence (KL) grade for the tibiofemoral compartments of each knee and each hip.13 Self-reported ethnicity was assessed by a nurse-administered questionnaire and only people of European descent were included in the genetic study. Additional details on these samples can be found in Neame et al1 and Lanyon et al.2
The Chingford study
The Chingford study is a prospective population-based longitudinal cohort of Caucasian women (assessed by questionnaire) derived from the age/sex register of a large general practice in north London, representative of the general UK population for weight, height and smoking characteristics. The study design and rationale have been described elsewhere in detail.14 The Guys and St Thomas’ Trust and Waltham Forest Trust ethics committees approved the study protocol. After study procedures were explained to participants, they gave written consent. The KL grade was scored for the tibiofemoral compartment of each knee.13 Radiographic knee OA was defined as a KL score ⩾2 of one or both joints. Radiographic hip OA was defined as joint space narrowing and a KL score ⩾2 at either the right or left hip.
Laboratory methods
For both Nottingham and Chingford study participants, genomic DNA was extracted from peripheral blood leukocytes of affected patients and controls using standard protocols. Genotyping was carried out by Kbioscience, Hertfordshire, UK. SNPs were genotyped using the KASPar chemistry, which is a competitive allele-specific PCR SNP genotyping system using FRET quencher cassette oligos. Genotyping accuracy, as determined from the genotype concordance between duplicate samples, was 99.6%. All three polymorphisms were in Hardy–Weinberg equilibrium in controls (p>0.05).
Statistical analysis
Allele odds ratios were calculated by comparing the number of alleles among cases and controls and the p value was computed using a Pearson χ2 test. Only an additive model was tested for alleles. For the DVWA SNPs only the allelic tests were carried out as this is the only model reported. Because a previous meta-analysis reported both additive and dominant models for the GDF5 polymorphism, we also tested different genetic models. A logistic regression was carried out with disease status being the dependent variable, genotype status as the independent variable 0 = CC, 1 = CT+TT in the dominant model and 0 = CC+CT, 1 = TT in the recessive model (including age, BMI and sex as covariates in the model). Only the best fitting model is reported.
Statistical power
The statistical power given the MAFs of the GDF5 and DVWA variants in our European samples given a specified odds ratio, for hip and knee OA was calculated using a log-additive (co-dominant) genetic model with an α = 0.05. The Quanto Software v 1.2. (University of Southern California, California, USA; http://hydra.usc.edu/gxe, accessed 14 July 2009) was used.
Fixed effect meta-analyses
In the absence of interstudy heterogeneity within samples we constructed a Mantel–Haenszel meta-analysis of data from the samples to assess the overall evidence of association. The Mantel–Haenszel χ2 test and the Mantel–Haenszel estimate of the odds ratio (ORMH)15 were used to provide a summary test and odds ratio.
Random effects meta-analysis
In the DerSimonian and Laird method, studies are considered as a random sample from a population of studies. The random effects model incorporates the heterogeneity of the studies. The overall treatment effect is estimated by a weighted average of the individual effects with weights inversely proportional to the variance of the observed effects. We tested the assumption of heterogeneity for each planned analysis using the method of DerSimonian and Laird based on work first presented by Cochran.15 The statistical significance of the combined effect for the DerSimonian–Laird odds ratio (ORDL) was estimated using the Z statistic, which is the ratio of the point estimate to its standard error.
Results
Table 1 presents the descriptive characteristics of cases and controls in the Chingford and Nottingham samples.
Descriptive characteristics of the study cohorts
We computed the statistical power available to us and assessed that given the sample size available combining both studies and the allele frequencies in controls we had 80% statistical power to detect a minimum odds ratio of 1.21 for either knee OA or hip OA for the GDF5 risk allele using a co-dominant (additive) genetic model, α = 0.05. For the DVWA variants, the major allele frequency is 85% in the UK controls, so the combined studies had 90.87% statistical power to detect an association with knee OA with an odds ratio of 1.34 (α = 0.05) for the major allele, which was the average odds ratio reported by Miyamoto and coworkers12 in their replication samples (Japan set C + Chinese). No previous data exist on hip OA and DVWA, but the current sample size would allow us to detect an odds ratio of ⩾1.30 for the major allele with 80% power and α = 0.05. Hence, this study is sufficiently powered to detect the size of genetic effects previously reported with knee OA for DVWA in replication studies.
We found that the major allele at the GDF5 variant was significantly associated with knee OA both in the Chingford and Nottingham studies (table 2). A combined analysis of the T allele using an additive model on knee OA resulted in an odds ratio ORMH = 1.29 (95% CI 1.14 to 1.47, p = 8×10−5) and of ORMH = 1.12 (95% CI 0.97 to 1.29 p<0.12) for hip OA. There was no evidence of heterogeneity between the two studies (I2 = 0% for both knee OA and hip OA). The genetic model found to give the smallest p value in these samples was a recessive one—that is, the TT genotype versus the CC +CT genotypes, and the GDF5 genotype was significantly associated with knee OA in both studies and also with THR in the Nottingham study after adjustment for sex, age and BMI (table 2).
Association between the GDF5 single nucleotide polymorphism rs143383 and osteoarthritis of the knee and the hip
Figure 1A shows that combining the present hip OA data with the European hip OA data from Chapman et al8 did not yield a significant result overall (OR = 1.07, 95% CI 0.98 to 1.16, p = 0.13). For knee OA data, on the other hand, a statistically significant odds ratio was seen combining the European samples reported by Chapman et al8 with our data (fig 1B). Therefore, our data increase the support in the literature for a role of the GDF5 promoter variant in the risk for knee OA.

Forest plot of study-specific estimates and fixed effects summary odds ratio (OR) and 95% confidence interval (CI) for the association between rs143383 and (A) hip osteoarthritis (OA), (B) knee OA.
The two SNPs in the DVWA gene reported to be associated with knee OA and tubulin binding in Asian populations are rs11718863 and rs7639618. The SNP rs11718863 encodes a Tyr to Asn substitution and rs7639618 encodes a Cys to Tyr change. The MAFs for these SNPs are much lower in European than in Asian samples as also seen in public domain data (http://www.hapmap.org/cgi-perl/gbrowse/hapmap_B36/, accessed 14 July 2009). We found that in UK controls from this study the MAFs of these SNPs were 15.33% for rs7639618 and 15.37% for rs11718863, yet were 48.3% and 48.0%, respectively, in the Japanese samples and >50% in the Chinese samples studied by Miyamoto and coworkers.12
In the UK samples studied here these two SNPs are in almost complete linkage disequilibrium (genotypes are identical for all except one of the subjects genotyped, resulting in r2 = 0.988 p<1×10−250 in controls). Hence we only show the association data for rs7639618 (table 3).
Association between DVWA rs7639618 (rs11718863)* single nucleotide polymorphism and osteoarthritis of the knee and the hip
The DVWA rs7639618 major allele, associated with increased risk of knee OA in Asians, was significantly associated with lower risk of TKR and not with radiographic knee OA nor with hip OA (table 3). Combining the two studies using TKR in the Nottingham cohort and radiographic OA in the Chingford we obtained a summary Mantel–Haenszel ORMH = 0.88 (95% CI 0.74 to 1.04, p<0.12) for the association of allele G with knee OA and heterogeneity between the two UK studies did not achieve statistical significance (I2 = 37%, p<0.21). For hip OA the combined analysis resulted in ORMH = 0.88 (95% CI 0.72 to 1.05, p<0.14).
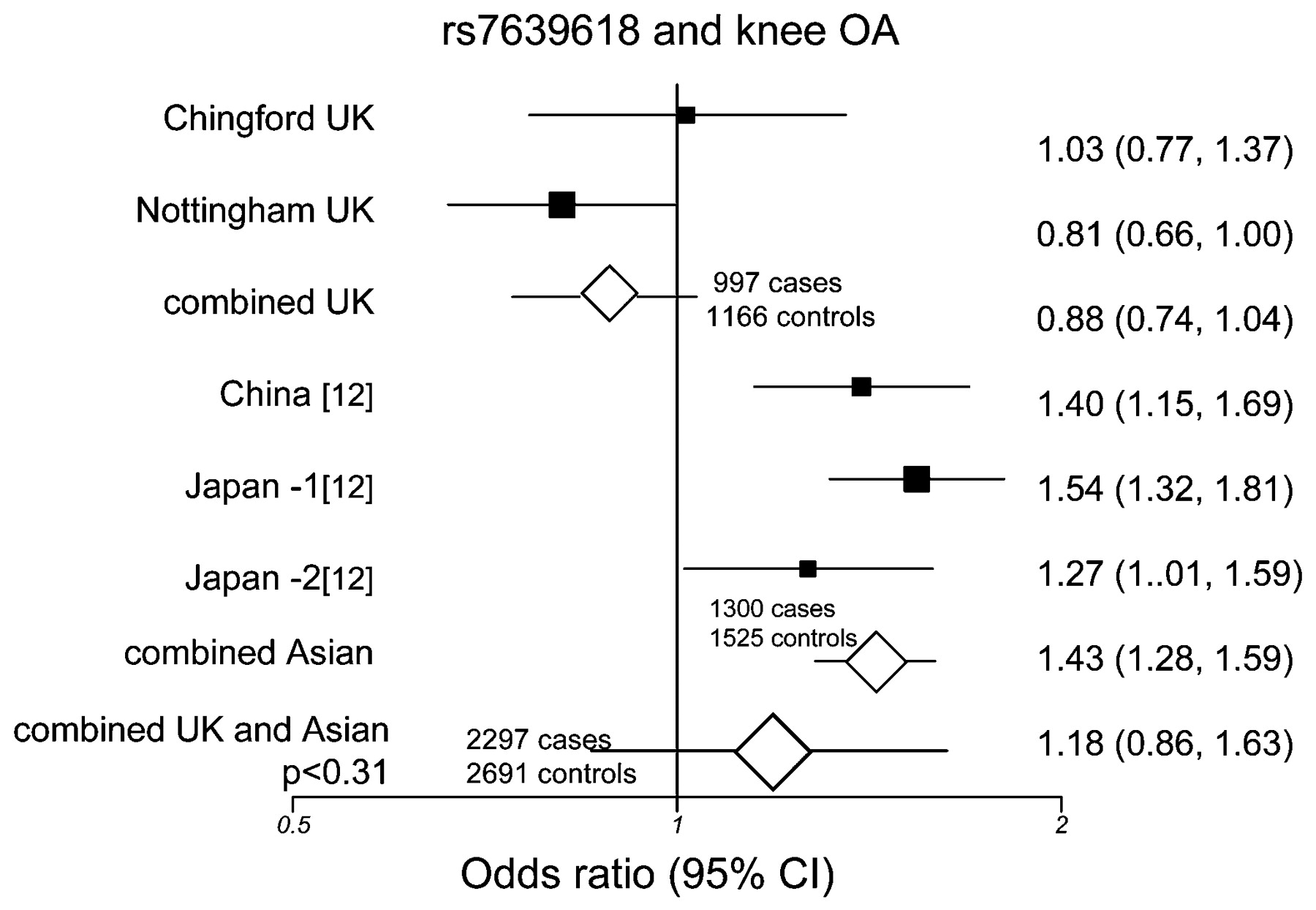
For comparison, the allele counts and frequencies in the Japanese and Chinese case–control studies previously reported12 are also shown (table 3). When we combined the current UK data with the published Japanese and Chinese data for rs7639618 we found very strong evidence of heterogeneity between studies (I2 = 91.8%, p<7 ×10−10) therefore we used a random effects DerSimonian–Laird model. Figure 2 summarises the odds ratios of individual studies and the meta-analysis for knee OA and the DVWA variant. The random effects model resulted in ORDL = 1.18, 95% CI 0.86 to 1.63, p = 0.309 for knee OA for the combined Asian and European studies, indicating no support for the involvement of this variant in the risk of knee OA overall.

{kind=link}
{kind=link}
Forest plot of study-specific estimates and random effects summary OR and 95% confidence interval (CI) for the association between rs7639618 and knee osteoarthritis in UK and Asian samples.
Discussion
In this study we have confirmed a role for the GDF5 promoter variant in genetic susceptibility to knee OA in people of European descent from the UK. Consistent with previous data, the association is stronger for knee OA than for hip OA, and we find that this association with knee OA in Europeans is found regardless of the genetic model used.
We also found that the major allele of non-synonymous polymorphisms in the DVWA gene, previously reported to be involved in tubulin binding and strongly associated with higher risk of knee OA in Chinese and Japanese patients, was, on the contrary, associated with a lower risk of TKR in a UK study. This variant was not associated with the milder radiographic form of the disease nor with hip OA in UK samples, even though our study was sufficiently powered to detect genetic associations of the same magnitude or smaller than those originally reported for variants in the DVWA gene. We must note, however, that our control samples consisted both of unrelated controls and of age-matched unaffected siblings of patients with OA, which might potentially decrease our power to detect an association. Nevertheless, because the allele frequencies in unaffected siblings and in unrelated controls were extremely similar to each other this is unlikely to have affected our final conclusions.
The major difference between our data in UK samples and the study in Asians is that the rs7639618 G allele which was associated with higher risk of knee OA in the original report is associated with lower or no risk in our study. When Asian and UK data were combined we did not find any evidence for association between knee OA and this variant.
There are at least two different explanations for this discrepancy. First, both our TKR result and those in the original report in Asians might be false positives and the meta-analysis result would appear to favour that hypothesis. However, a second explanation might be that DVWA is really implicated in the pathogenesis of OA but that rs7639618 and the linked rs11718863 are not the true functional variants in the DVWA region involved in risk of OA. In such a hypothetical scenario it would be possible that a different SNP could be truly implicated in the pathogenesis of knee OA and such a theoretical SNP would need to be in positive linkage disequilibrium (LD) in the UK cohorts studied here with the “A” allele in rs7639618 (encoding a Tyr) but in negative LD with the “A” allele and in positive LD with the “G” allele (encoding a Cys) reported in the Asian cases.12
The minor allele frequency of rs7639618 is three times lower among Caucasian than among Asian samples both in public domain data and in our study. Further, some notable differences in LD patterns can be seen for some of the SNPs in the same LD block studied by Miyamoto and coworkers.12 For example, rs353093 is an intronic SNP in DVWA which was also associated with knee OA in that study with p<6 ×10−4. LD between rs7639618 and rs353093 is r2 = 0.53 in Asians from HapMap and >0.7 in the Japanese samples studied by Miyamoto et al.12 In Caucasians on the other hand (CEPH data from HapMap release B36), LD is almost absent with an r2 = 0.028. Hence, differences in LD in these two populations might account for the observed discrepancy.
The data at hand cannot definitively exclude the possibility that the discrepancies we find are due to LD and we must also note that a lack of reproducibility among Caucasian patients has been described of other very strong genetic associations with OA previously reported in Asians. A recent meta-analysis of the EDG2 gene16 found no evidence for association of a promoter variant with OA in subjects of European descent, although it had been previously reported to be strongly associated with knee OA in Japanese patients. Similarly, the association of the CALM1 promoter variant was not replicated in any of the Caucasian samples tested to date.8 9 Such lack of reproducibility of Asian associations in Caucasian patients may have a number of explanations, including genetic or even environmental differences between Asians and Europeans in their risk of OA.
The results from this study indicate that the differences in tubulin binding by the DVWA protein due to the amino acid changes encoded by rs7639618 and rs11718863 are unlikely to be responsible for any functional involvement of this gene in OA in Caucasians.
Acknowledgments
This work was supported by the European Commission Framework 7 (grant number 200800 TREAT-OA) and by the Arthritis Research Campaign (grant numbers 16170, L0529, L0510 and 17489).
REFERENCES
Footnotes
Competing interests None.
Ethics approval Approval from Nottingham City Hospital and North Nottinghamshire, Guys and St Thomas’ Trust and Waltham Forest Trust ethics committees.