Article Text

Abstract
Background Histone deimination regulates gene function and contributes to antimicrobial response, allowing the formation of neutrophil extracellular traps (NETs). Deiminated proteins are target of anti-citrullinated peptides antibodies (ACPA) in rheumatoid arthritis (RA).
Objective The objective of this paper is to test the hypothesis that RA sera react with deiminated histones contained in NETs.
Methods Neutrophils from peripheral blood were stimulated with A23187 and acid treated; NETosis was induced by phorbol myristate acetate, and NET proteins were isolated. Sera were tested by immunoblot on acid extracted proteins from neutrophils and from NETs, and by ELISA on deiminated histone H4 or H4-derived peptides. Bands reactive with RA sera were excised from gels, digested with trypsin and subjected to matrix-assisted laser desorption/ionisation time of flight (MALDI-TOF) analysis, before and after derivatisation to detect citrullinated peptides.
Results RA sera reacted with a deiminated antigen of 11 KDa from activated neutrophils, recognised also by anti-H4 and antideiminated H4 antibodies. A similar reactivity was observed with NET proteins. The antigen from neutrophils or NETs was identified as citrullinated H4 by MALDI-TOF analysis. By ELISA, RA sera bound in vitro citrullinated H4. Citrullinated H4 14–34 and 31–50 peptides detected antibodies in 67% and 63% of RA sera and in less than 5% of controls; antibody titre was correlated with anti-CCP2.
Conclusions Citrullinated H4 from activated neutrophils and NETs is a target of antibodies in RA, and synthetic citrullinated H4-derived peptides are a new substrate for ACPA detection. As NETosis can generate antigens for ACPA, these data suggest a novel connection between innate and adaptive immunity in RA.
- Ant-CCP
- Autoantibodies
- Rheumatoid Arthritis
Statistics from Altmetric.com
Deimination, a post-translational modification converting protein arginine residues in the non-standard amino acid citrulline, has the potential to alter the structure, antigenicity and function of proteins.1 Deimination affects proteins in the normal development of tissues and organs such as skin or brain, as well as in inflammation; it is catalysed by a family of calcium-binding enzymes, called peptydilarginine deiminases (PAD) (enzyme classification (EC) 3.5.3.15), with different tissue distribution. PAD4 is highly expressed in peripheral blood mononuclear cells and, among PAD, is the only one that has a classical nuclear localisation signal.2 Thus, although predominantly localised in the cytoplasm, PAD4 can move to the nucleus upon cell activation.3 The nuclear substrates of PAD4 are nucleophosmin and histones, namely H2A, H3 and H4.4
The deimination of histones is a crucial event in cell biology. On the one hand it regulates gene function, antagonising arginine methylation, which is linked to active transcription.5 On the other, it represents an integral part of the response to infections,6 causing the unfolding of chromatin during neutrophil extracellular traps (NET) formation7 In neutrophils (but also in eosinophils and in mast cells), different inflammatory stimuli, probably acting on PAD4, induce histone deimination, chromatin decondensation and NET formation.8 ,9 Core histones are the most abundant proteins of these web-like structures that trap and kill microbes and represent an essential component of innate immunity responses.10
While deimination is a physiological process amplified in inflammatory conditions, only individuals carrying genetic predisposition to develop rheumatoid arthritis (RA) make antibodies to deiminated proteins.1 These antibodies, collectively identified as ACPA, react with different deiminated proteins11 and display partially overlapping specificities.12 ,13 Because of their strict association with RA and their presence in the early phases of the disease (and even before disease onset) ACPA are now included in the classification criteria for RA.14
The aim of this paper is to investigate the presence in RA sera of a new subset of ACPA antibodies reactive with deiminated histones.
Patients and methods
Patients
One hundred and fifty-two consecutive patients with RA (119 women and 33 men; mean age 56.2 years, range 19–81; mean disease duration 9 years, range 9 months—40 years) diagnosed according to the American College of Rheumatology/European League Against Rheumatism (ACR/EULAR) classification criteria14, were enrolled in this study.
One hundred and forty-one control subjects were also studied, comprising normal healthy subjects (NHS—n=50; blood donors and laboratory personnel) and patients with: ankylosing spondylitis (AS—n=12); psoriatic arthritis (PsA—n=12); systemic sclerosis (SSc—n=7); mixed cryoglobulinaemia (MC—n=12); systemic lupus erythematosus (SLE—n=48). AS was diagnosed according to the revised New York criteria15 and PsA according to the criteria of Vasey and Espinoza.16 The diagnoses of SLE17 and SSc18 were made based on the ACR criteria; MC was diagnosed in the presence of Meltzer's triad (purpura, weakness and arthralgia) and cryoglobulins in the sera.19
Written consent of all study participants was obtained in accordance with the Declaration of Helsinki, and the study was approved by the local ethics committee.
Serum samples were stored at −20°C until tested.
Neutrophils isolation and stimulation
Neutrophils were obtained from buffy coat of healthy blood donors and isolated using discontinuous gradient centrifugation according to English et al.20
Purified neutrophils were resuspended at 2×106 cell/ml in Locke's solution (NaCl 150 mM, Hepes-HCl 10 mM pH 7.3, CaCl2 2 mM, Glucose 0.1%) and incubated at 37°C for 15 min with or without A23187 4 μM (Sigma-Aldrich, Saint Louis, Missouri, USA), followed by a further incubation of 1 h in serum-free Roswell Park Memorial Institute medium (RPMI) at 37°C.
NET preparation
Neutrophils were seeded in Petri dishes in RPMI at 2×106 cells/w and activated with 100 nM phorbol myristate acetate (PMA) for 4 h at 37°C.21 After removing the medium, the wells were washed 2×10 min with Dulbecco-modified phosphate buffer saline (D-PBS) and incubated for 20 min at 37°C with 10 U/ml DNase I (Sigma) in RPMI. DNase activity was stopped by adding EDTA 5 mM (final concentration). The samples were then centrifuged at 3000 g to remove intact cells and intact nuclei; the supernatants containing NET proteins were processed as described below.
Histone extraction
Stimulated and unstimulated neutrophils and NETs were incubated overnight in H2SO4 0.2 M at 4°C with agitation. Acid extracted proteins were then precipitated with 33% trichloroacetic acid (TCA) for 2 h at 4°C, washed twice with acetone and suspended in ddH2O.22 Protein concentrations were determined using bicinchoninic acid (BCA) Protein Assay (Pierce, Rockford, Illinois, USA).
SDS-PAGE and immunoblotting
Acid extracted proteins from stimulated and unstimulated neutrophils and from NETs were resolved in a 16.5% Tris-Tricine-sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS PAGE) (Bio-Rad, Hercules, California, USA) under non-reducing conditions and blotted onto polyvinyliden fluoride (PVDF) (Millipore, Billerica, Massachusetts, USA). The membrane strips were saturated for 30 min at room temperature in tris buffer saline (TBS) containing 5% bovine serum albumine (BSA) and 0.05% Tween-20, and incubated overnight at 4°C with human sera diluted 1 : 200, anti-Histone H4 (Upstate, Millipore), and anti-histone H4 (citrulline 3) (Upstate, Millipore) rabbit antisera diluited 1 : 500.
Anti-Citrulline (modified) detection kit (Upstate, Millipore) was used to detect deiminated proteins, following the manufacturer's instructions.
Peroxidase activity was visualised by means of enhanced chemiluminescence using Luminata Western HRP Substrate (Millipore). Images were acquired and analysed using the VersaDoc Imaging System and QuantityOne analysis software (Bio-Rad).
In-gel digestion
The gel bands of interest were chopped into small pieces, washed with water and shrunk with acetonitrile. Reduction and alkylation were performed in a solution of 10 mM dithiothreitol (DTT) (56°C, 45 min) and iodoacetamide 55 mM (room temperature, 30 min, in the dark), respectively. Shrinking and rehydratation were carried out with acetonitrile and 100 mM ammonium bicarbonate in order to completely remove Coomassie staining. Proteins were digested by adding a solution of 12 ng/µl of trypsin (type IX-S, from porcine pancreas, Sigma Aldrich) in 10 mM ammonium bicarbonate (37°C, overnight). Digestions were stopped with 10% trifluoroacetic acid (TFA), and the supernatants were recovered for subsequent mass spectrometry analyses.
Chemical modification
Derivatisation of citrulline residues was performed as described.23 ,24 Briefly, 10 µl of proteolytic digest was added to 10 µl of 50 mM antipyrine in presence of 20 µl of 10% TFA; 10 µl of freshly prepared 2,3-butanedione 50 mM was finally added to start the reaction. After 2 h of incubation at 37°C, in the dark, the samples were SpeedVac-concentrated up to 20 µl and desalted by Zip-Tip C18 pipette tips (Millipore).
MALDI-TOF analyses
A matrix-assisted laser desorption/ionisation- time of flight MALDI-TOF/TOF spectrometer Ultraflex III (Bruker Daltonics, Bremen, Germany) was set up in positive Reflectron mode, and the spectra were acquired in the mass range of 860–4000 m/z.
For the analyses of the proteolytic digests, the dried droplet deposition method was applied using a solution of α-cyano-4-hydroxycinnamic acid spotted in an AnchorChip (Bruker Daltonics) target plate. Peptide mass fingerprint data were obtained for each sample, and mass-spectrometry-mass spectrometry (MS-MS) analyses were performed on selected precursors. Mascot searches were done against the Swiss-Prot database calculating two maximum missed cleavages for trypsin, carbamidomethylation of cysteine as fixed modification, oxidation of methionine as variable modification, 30 ppm and 0.6 Da as precursors and fragments tolerances, respectively. For the analyses of the derivatised digests, the same instrumental conditions were applied.
H4-derived synthetic peptides
Linear peptides I–XI of 20 aa (table 1) corresponding to the whole sequence of the human histone H4 (P62805—H4_human) and with a 5-mer overlapping were synthesised by Toscana Biomarkers (Siena, Italy). Peptides II and IV were also synthesised as multiple antigen peptides (MAP) bearing four identical sequences on a lysine scaffold25 giving products XII and XIII (Histone Citrullinated Peptide HCP1 and HCP2, respectively). Moreover, peptides II, III and IV were also synthesised in the non-citrullinated form IX, X and XI.
Sequences of H4-derived synthetic peptides I–XIII
Peptides were prepared on an automatic (APEX, AAPPTec) or on a microwave-assisted (Discover, CEM) synthesiser following the Fmoc/tBu solid-phase peptide strategy.26 ,27 Peptides were purified by high performance liquid chromatography (HPLC) (purity >95%) and characterised by electrospray ionisation-mass spectrometry (ESI-MS).
The sequences of products I–XIII are shown in table 1.
Deimination of histone H4
Purified PAD from rabbit skeletal muscle was purchased from Sigma. Human recombinant histone H4 (Millipore) was incubated at 1 mg/ml with 7.5 units/ml of PAD in 0.1 M Tris HCl (pH 7.4), 10 mM CaCl2, and 5 mM dithiothreitol for 2 h at 50°C. H4, diluted in the deimination buffer and kept at 50°C for 2 h without PAD, was used as the control.
Purification of anti-HCP1 and anti-HCP2 antibodies
HCP1 (XII) and HCP2 (XIII) were conjugated to CNBr-activated Sepharose (Sigma) according to standard procedures. Total immunoglobulins from RA sera containing anticitrulline antibodies were precipitated with 50% saturated ammonium sulfate; the precipitates were dissolved in phosphate buffer (pH 7.4) and dialysed overnight against phosphate buffered saline (PBS). Enriched immunoglobulin preparations were applied to the column, and the flowthrough was collected for subsequent analysis. The column was extensively washed with 20 mM Na2HPO4, 150 mM NaCl (pH 7.2), and the antibodies bound to the column were eluted by 0.1 M glycine buffer (pH 2.8) (0.5 ml/fraction), immediately neutralised with 50 μl Tris 1 M (pH 8.0), and dialysed overnight against PBS. The antipeptide antibody content in the eluates and flowthrough was tested by ELISA.
Direct ELISA and inhibition assays
In the direct binding assay, ELISA polystyrene plates (Nunc MaxiSorp F96; Nunc, Roskilde, Denmark) were coated with linear peptides I–XI (10 μg/ml) or MAPs XII and XIII (5 μg/ml) or histone H4 (5 μg/ml) in 50 mM sodium carbonate/bicarbonate buffer pH 9.6 and incubated overnight at 4°C. Saturation was carried out with PBS containing 1% porcine gelatin (Sigma Aldrich) for 45 min at room temperature. Sera diluted 1 : 200 in PBS, 0.5% porcine gelatin and 0.05% Tween-20 were incubated on the plates for 3 h at room temperature.
In ELISA employing histone H4 or deiminated histone H4, sera were diluted in PBS, 0.5% porcine gelatin, 0.05% Tween-20, containing also NaCl 0.5 M, to reduce non-specific binding.
After washings with PBS, 1% Tween-20 and PBS, alkaline phosphatase-conjugated antihuman IgG (Sigma) diluted 1 : 3.000 was added to the wells, and the plates were incubated for 2 h at room temperature.
Alkaline phosphatase activity was revealed with p-nitrophenyl phosphate in sodium carbonate/bicarbonate buffer.
Anti-cyclic citrullinated peptide (CCP2) antibodies were detected using a commercial kit (QUANTA Lite CCP, Inova Diagnostics, San Diego, California, USA), according to the manufacturer's instructions.
For the inhibition assays, sera from patients with RA were tested on deiminated H4 at different dilutions. The dilution corresponding to 50% of the maximum binding of each serum was preincubated with deiminated and non-deiminated H4-derived peptides for 30 min at 37°C before being transferred to the deiminated H4-coated plates. Thereafter, ELISAs were carried out as for direct binding assay. Inhibition assays employing affinity purified anti-HCPs antibodies were carried on by the same protocol.
Results
RA sera react with an antigen expressed in nuclei of activated neutrophils
Neutrophils were activated by A23187 stimulation for 15 min at 37°C, lysed directly in H2SO4, and acid extracted proteins were precipitated in TCA.
Acid precipitated proteins were separated on acrylamide gels, transferred to PVDF and probed with RA sera or antisera specific for histones. As shown in Figure 1A, RA sera and antideiminated H4 reacted with a band of identical molecular weight, decorated also by antimodified citrulline antiserum.
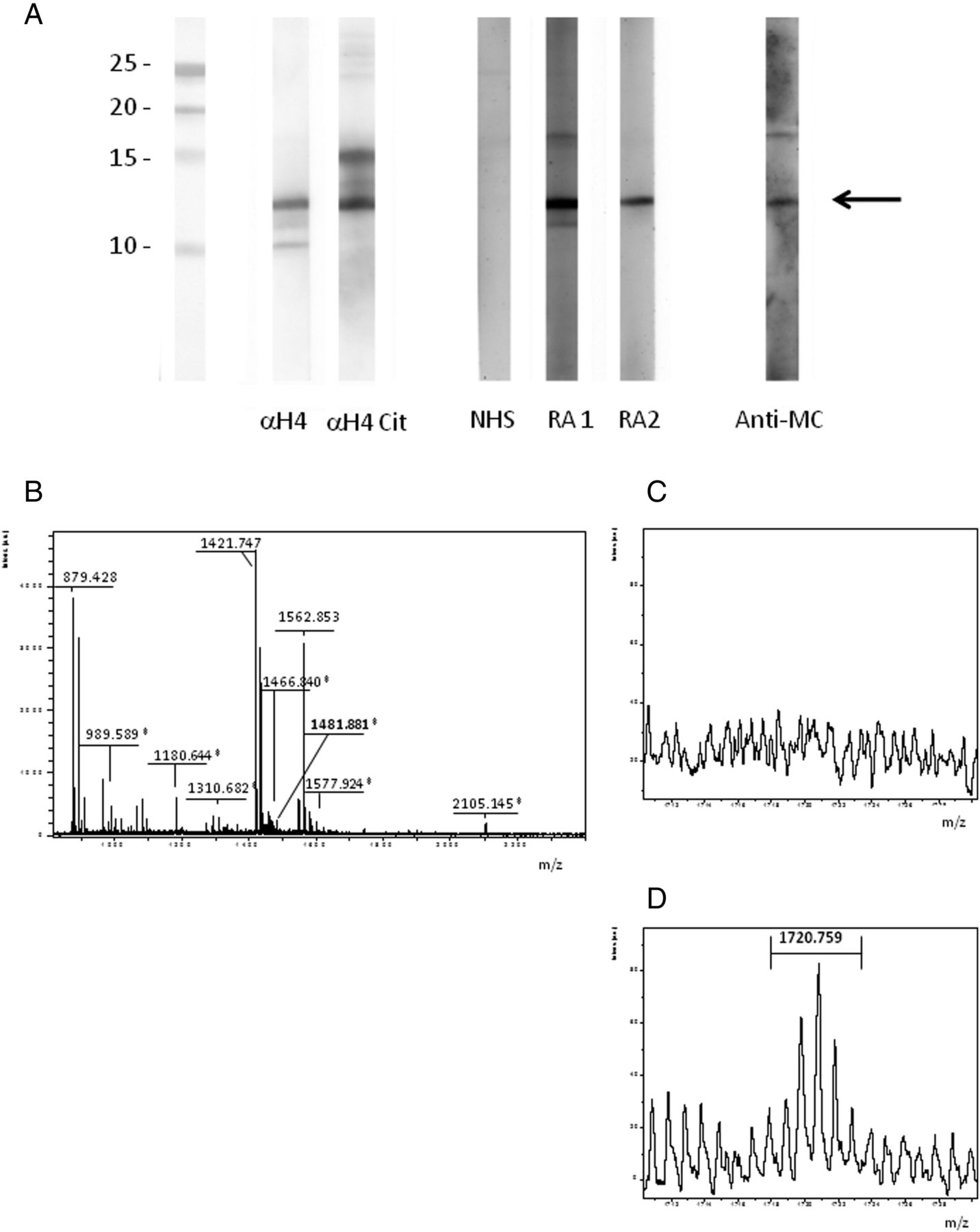
RA sera bind to citrullinated H4 from stimulated neutrophils. (A) Acid extracted proteins from A23187-stimulated neutrophils were subjected to western blotting and probed with NHS or RA sera, with anti-H4 (α-H4) or anticitrullinated H4 (Anti-H4cit) polyclonal antibody and with antimodified citrulline (Anti-MC) antibody. A protein of 11 KDa is bound by RA sera, by anti-H4cit and by anti-MC. (B) MALDI-TOF spectrum obtained from the excised band (→) after tryptic digestion; peptide sequencing shows that H4 (*) and S100A8 are contained in the band. (C), (D) MALDI-TOF spectrum of tryptic digest before (C) and after (D) derivatisation with 2,3 butanedione and antipyrine; MS analysis shows the presence of the mass of the unmodified sequence at 1481.8 m/z, the citrullyl sequence at 1482.8 m/z, and the derivatised citrullyl sequence at 1720.8 m/z displaying the expected isotopic pattern (D).
The band at 11 kDa was excised from the acrylamide gel, treated with DTT and iodoacetamide, and then digested with trypsin. The peptides obtained by trypsin digestion were subjected to MALDI-TOF analysis (Figure 1B). By MASCOT search on Swissprot database, two proteins were identified: histone H4 (P62805; p=0.0052) and S100A8 (P05109, p=0.00072). The sequences identified are reported in online supplementary table S1.
Histone H4 and S100A8 contain several arginine and lysine residues liable to deimination (causing a 0.984 Da mass increase) and carbamylation (causing a 43.006 Da mass increase), respectively.
The tryptic digest was thus treated with antipyrine and 2,3-butanedione. The comparison of the mass profiles before and after the treatment with antipyrine and 2,3-butanedione showed one citrulline but no homocitrulline in the histone H4 (Figure 1C, D). The citrulline is located in the tryptic sequence 23–35 of H4, and more specifically at position 23 (sequence: CitDNIQGITKPAIR) as confirmed by MALDI-TOF/TOF analysis (see online supplementary figure S1).
Taken together, these data suggest that RA sera and antideiminated H4 antibody recognise deiminated H4 within nuclear extracts from activated neutrophils.
RA sera react with purified H4 and with H4-derived deiminated peptides
Human purified H4 was deiminated by PAD4 in vitro and the presence of citrullyl residues at positions 23, 35, 36 and 45, was confirmed by MALDI-TOF/TOF analysis (see online supplementary table S2). To analyse the reactivity of RA sera, deiminated H4 was used as an antigen to measure IgG antibodies by ELISA assay. The binding of RA sera to deiminated H4 was significantly higher than the binding of NHS (p<0.0001). On the contrary, no difference was observed with unmodified H4 (Figure 2).

Binding of NHS and RA sera to native and in vitro deiminated H4. Anti-H4 and antideiminated H4 antibodies were measured by ELISA. Results are expressed as absorbance at 405 nm. RA sera specifically bound the deiminated form of H4 (p<0.0001); the binding was significantly higher than the one observed with NHS (p<0.0001). Setting the cut-off value at the 97.5° percentile of the NHS population, 17% of RA sera are positive on deiminated H4 as compared with 5% on native H4.
Epitope scanning of H4 was performed by means of synthetic peptides. Overlapping synthetic peptides covering the whole H4 sequence were synthesised, substituting arginine with citrulline, and used as antigens on the solid phase (table 1: peptides I-XI). IgG binding to peptides II and IV were detected in 10% and 13% of RA sera, respectively (data not shown). Less than 5% of RA sera reacted with the other H4-derived citrullinated peptides. On the whole, increasing the number of citrullyl residues in the peptide, result in a higher number of reactive RA sera (data not shown).
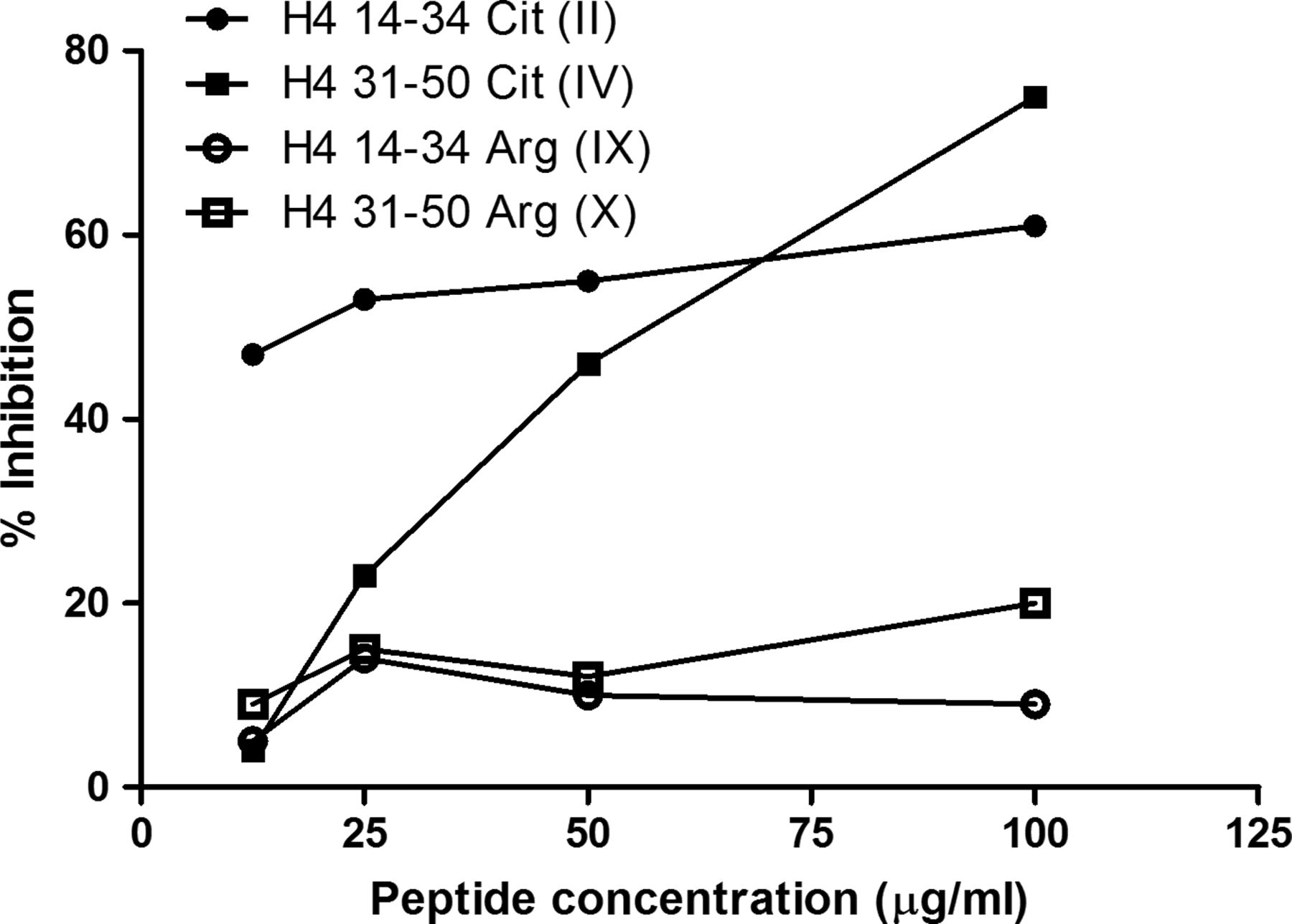
Inhibition assays were employed to test whether synthetic peptides represent epitopes which are recognised by RA sera in the entire protein. As shown in figure 3, preincubation of RA sera with peptides II and IV inhibited the binding to in vitro deiminated H4.

Inhibition of serum binding to deiminated H4. RA sera were preincubated with arginine-containing or citrulline-containing H4 14–34 and H4 31–50 peptides (II, IV, IX, X—table 1) and transferred to deiminated H4-coated plates. Citrullinated form of both peptides inhibited the binding to deiminated H4, while no inhibition were obtained with the arginine containing peptides. A representative example is shown in the graph. The results are expressed as the percentage inhibition of binding: 100−(100×optical density (antibody+peptide)/optical density (antibody+buffer)).
H4-derived deiminated peptides as tool to detect ACPA
Peptides II and IV were synthesised as multiple antigen peptides (HCP1—XII; and HCP2—XIII, see table 1), and used in ELISA. Setting the threshold of normality at 97.5 percentile of a normal population, IgG anti-HCP1 were detected in 67% and anti-HCP2 in 63% of RA sera (Figure 4A and B) and in less than 5% of disease controls (p<0.0001). The majority of RA sera (75%) react with HCP1 and HCP2, 15% with HCP1 only and 10% with HCP2 only.

Levels of anti-HCP1 and anti-HCP2 antibodies in RA and disease control patients, correlation with anti-CCP2 and fine specificity. (A and B) Anti-HCP1 and anti-HCP2 antibodies were measured by ELISA in NHS (normal healthy subjects), ankylosing spondylitis, psoriatic arthritis (PsA), systemic sclerosis (SSc), systemic lupus erythematosus and mixed cryoglobulinemia. Results are expressed as arbitrary units/ml (AU/ml). Horizontal bars represent median values. Anti-HCP1 and anti-HCP2 are present in 67% and 63%, respectively, and in less than 5% than controls (p<0.0001). (C and D). Anti-HCP1 and anti-HCP2 levels are highly correlated with levels of anti-CCP2 antibodies (p<0.0001). (E and F). Fine specificity of anti-HCP1 (E) and anti-HCP2 (F) antibodies. Affinity-purified anti-HCP1 and anti-HCP2 immunoglobulin (Ig)G antibodies from two representative patients with RA were preincubated with different amounts of HCP1, HCP2, VCP1, VCP2, and control peptides before being transferred to HCP1- and HCP2-coated plates, respectively. The results are expressed as the percentage inhibition of binding: 100-(100 × optical density (antibody+peptide)/optical density (antibody+buffer)).
Anti-HCP1 and anti-HCP2 titres were significantly correlated with anti-CCP2 antibodies (Figure 4C and D; p<0.0001).
As already observed with antibodies to viral citrullinated peptides from EBNA1 (VCP1) and EBNA2 (VCP2),13 anti-HCP antibodies can bind different citrullinated substrates. We analysed the cross-reactivity of anti-HCP1 and anti-HCP2 antibodies from two different patients with RA by liquid-phase inhibition assays. The binding of anti-HCP2 antibodies is inhibited by preincubation with VCP1 and VCP2, whereas anti-HCP1 purified antibodies from another patient with RA display limited cross-reactivity (Figure 4E and F).
The data so far obtained indicate that RA sera react with deiminated H4.
As deiminated H4 is a constituent of NETs, we analysed the reactivity of RA sera with proteins contained in NETs.
RA sera react with deiminated H4 contained in NETs
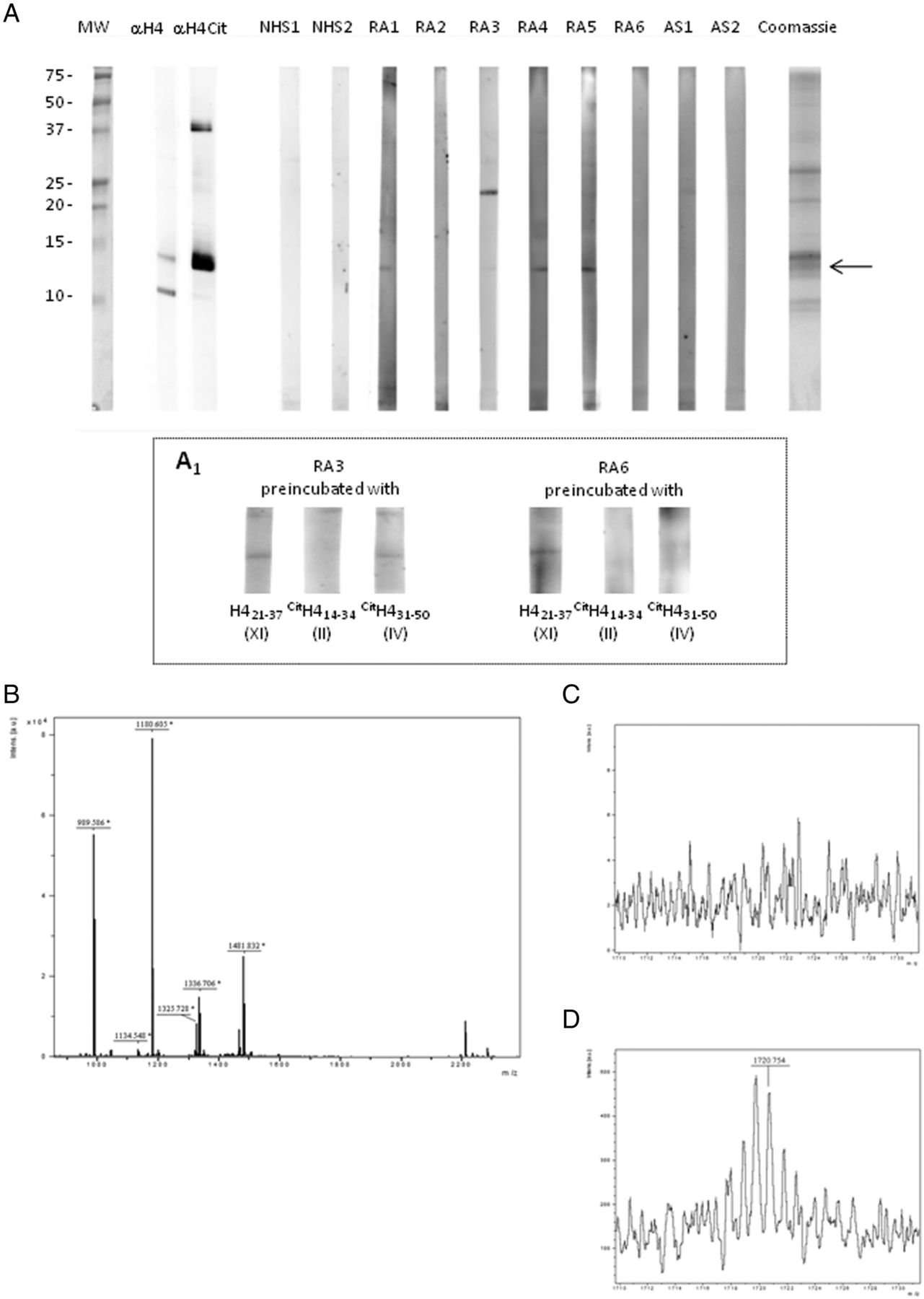
Granulocytes were in vitro stimulated with PMA to induce NETosis; NET proteins were isolated by DNase I digestion, acid precipitated, fractionated on Tris- Tricine SDS-PAGE and transferred to PVDF. Strips were incubated with NHS, RA and disease control sera. As shown in figure 5A, RA sera, but not disease control sera, decorated a band that reacted also with antideiminated H4 antibodies. The binding can be inhibited by preincubation of sera with citrullinated H4-derived peptides II and IV. (figure 5 A1)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
RA sera bind to citrullinated H4 from neutrophil extracellular traps (NET). NET proteins from phorbol myristate acetate-stimulated neutrophils were subjected to western blotting and probed with NHS, RA and ankylosing spondylitis sera and with anticitrullinated H4 (Anti-H4cit) polyclonal antibody. RA sera and by anti-H4cit recognise a band of identical MW. The binding can be inhibited by preincubation of sera with citrullinated H4-derived peptides II and IV (A1). (A) MALDI-TOF spectrum obtained from the excised band (→) after tryptic digestion; peptide sequencing shows that H4 is contained in the band. (B), (D) MALDI-TOF spectrum of tryptic digest before (C) and after (D) derivatisation with 1,3 butanedione and antipyrine; MS analysis shows the presence of a citrullinated H4-derived peptide (m/z 1720.759) displaying the expected isotopic pattern (D).
The band of interest was excised and, after in-gel digestion with trypsin, proteolytic peptides were submitted to MALDI-TOF analysis leading to the identification of histone H4. The derivatisation technique with antipyrine and 2,3-butanedione allowed to identify the sequence H423–35 with citrulline at position 23 (figure 5C and D; online supplementary figure S2).
These data suggest that RA sera target deiminated H4 from NETs proteins.
Discussion
RA sera contain a variety of autoantibodies, but those specific for citrullinated antigens are of utmost interest, being highly specific for the disease, probably endowed with pathogenic potential, and possibly related to basic mechanisms of the disease.11 It is now accepted that ACPA represent a family of antibodies with overlapping specificities that are directed to a variety of citrullinated substrates of exogenous or endogenous origin.12 ,13 In this paper, we identify citrullinated H4 as a novel substrate for ACPA and show that NETosis can be the source of this antigen, thus establishing a novel connection between innate and adaptive immunity in RA. Moreover, we describe synthetic citrullinated H4-derived peptides as a new substrate for ACPA detection.
Several lines of evidence indicate that histone deimination is the result of PAD4 activity. It has been shown that PAD4 is localised in nuclei,2 and can convert multiple arginine sites of H3 and H4 into citrulline.28 PAD4 can, in fact, deiminate Arg2, Arg8, Arg17 and Arg26 on histone H35 and Arg3 on the N-terminal sequence shared by H2A and H4.4
PAD4 activation and histone deimination is triggered in HL60 cells and blood granulocytes by a variety of stimuli, such as Ca2 + ionophore, lipopolysaccharide (LPS), TNF, chemoattractant peptides and H2O2.6
Under these stimuli, neutrophils, eosinophils and mast cells extrude NETs, undergoing a novel form of cell death that requires reactive oxygen species produced by NAPDH-oxidase.
In neutrophils stimulated with Ca2+ ionophore, the qualitative and quantitative NET composition has been analysed by a proteomic approach.10 Core histones are the most abundant proteins, accounting for approximately 70% of all NET-associated proteins. H3 and H4 are present in lower amounts than H2B and H2A, and their molecular weight is reduced when compared with nuclear histones, suggesting post-translational modifications. Indeed, by confocal microscopy, deiminated H3 was detected in NETs.6 The crucial role of deimination is outlined by two sets of observations: PAD4 inhibition blocks NET formation,7 and PAD4 deficient neutrophils lose the capacity to form NETs in vivo and in vitro.9
In this paper, we identify deiminated H4 as a target of antibodies in RA; moreover, we show that RA sera bind deiminated H4 contained in NET.
In fact, by immunoblot, we detected a reactivity of RA sera with a band whose identity with H4 was suggested by specific antibody recognition and demonstrated by MALDI-TOF.
Citrullination of H4 was suggested by reactivity with anticitrulline antibodies and proved by MALDI-TOF/TOF analysis after peptide derivatisation. The specific reactivity of RA sera with H4 was further confirmed by ELISA assays on in vitro deiminated human H4 and on citrullinated synthetic peptides corresponding to H4 sequences. Furthermore, we showed RA sera reactivity with isolated NET proteins; again, the band was identified as deiminated H4 using specific antibodies and, subsequently, by MALDI-TOF/TOF analysis of tryptic peptides from the digested band before and after derivatisation.
As already reported by Urban et al,10 commercial antisera specific for histones display cross-reactivities that impair the correct identification of molecular targets. Protein identification was thus carried out by mass spectrometry. Moreover, due to the specific interest on deiminated proteins, a chemical modification with antipyrine and 2,3-butanedione was applied to the proteolytic digest in order to identify citrullines. These reagents, employed in the antimodified citrulline detection kit, recognise also carbamylated proteins.29 ,30 The reaction gives rise to an adduct with a mass increase of 239.09 Da for deiminated arginine and 281.11 for carbamylated lysine, with respect to their unmodified counterpart; moreover, MS spectra of the modified citrullinated peptides gain the characteristic isotopic pattern described by Holm et al.23 that we actually found in the derivatised digest.
By these tools, H4 and S100A8 were identified in the band recognised by RA sera in nuclear extracts from activated granulocytes. The calgranulin, S100A8, is a calcium-binding protein endowed with proinflammatory and regulatory properties, highly abundant in cytosol of neutrophils and monocytes.31 Although its levels are increased in RA sera, it has never been described as a target of autoantibodies.32 Even if we did not detect citrullinated S100A8-derived peptides after derivatisation, the protein contains two arginines susceptible of deimination. Experiments are in progress to test the reactivity of RA sera with native or citrullinated S100A8.
The presence of autoantibodies reactive with deiminated histones has been previously reported in patients with RA developing Felty syndrome.33 Antibodies displaying a preferential binding to in vitro deiminated calf thymus histones have been detected in a high percentage of patients affected by Felty syndrome, but only in low frequency and amounts in RA or SLE sera. By immunoblot, a preferential binding to deiminated H3 was observed, and ELISA confirmed the reactivity of sera on recombinant H3.
At variance with the data reported by Dwivedi et al,33 we found a preferential reactivity of RA sera with deiminated H4. Several differences in the experimental protocols may explain the different results: in this paper, sera reactivity was tested on in vivo deiminated histones from activated granulocytes, and confirmed by ELISA on in vitro deiminated purified H4. A different format of the ELISA assay (different solid phase, diluting buffer containing NaCl 0.5 M) may have permitted the detection of a preferential binding of RA sera to deiminated H4. High ionic strength buffer reduced the non-specific binding of normal sera to deiminated and non-deiminated H4, allowing the detection of significant binding in 9/67 (13%) of RA sera. The presence in normal sera of antibodies reactive with non-deiminated sequences of histones may increase the background and hamper the detection of antibodies that target citrullinated histones. In fact, synthetic peptides corresponding to deiminated sequences of H4 showed low reactivity with normal sera. Performing an epitope mapping of citrullinated H4, we detected two sequences (aminoacids 14–34 and 31–50) recognised by a high percentage of RA sera.
The ELISA assay employing the two peptides as MAPs allows the detection of antibodies in a high percentage of RA sera (67% and 63%, respectively) and in a very low number of normal or disease control sera (<5%). This ability to discriminate RA from controls, together with the high correlation with well established assays like CCP2, indicates that citrullinated H4 14–34 and 31–50 are suitable substrates for the detection of ACPA. The description of autoantibodies reactive with citrullinated histones raises important questions on the mechanisms leading to their production. Considering this, it is of utmost interest that both studies showing the reactivity of RA or Felty syndrome sera with deiminated histones detect binding to NET components. In fact, Dwivedi et al33 using immunofluorescence showed that immunoglobulins from patients with Felty syndrome decorated NET fibers; using immunoblot we showed the reactivity of RA sera with extracted NET proteins that were further characterised by sequencing.
A role of NET components in the induction of autoantibodies has already been proposed in SLE34 and ANCA-positive vasculitis.35 A subset of SLE patients are characterised by a reduced clearance of NETs; this defect may be caused in some patients by DNase inhibitors in sera and in others by anti-NET antibodies that protect NETs from degradation. Moreover, SLE neutrophils and, in particular, a low-density subset, displayed significantly enhanced NET formation, and netting neutrophils have been found to infiltrate lupus skin and kidneys.36 Similar data have been reported in ANCA-positive vasculitis. Kessenbrock et al35 identified cathelicidin or LL37, an antimicrobial peptide produced by epithelial cells and granulocytes, as a NET component. In extracellular sites, LL37 can form complexes with DNA or RNA, protecting nucleic acids from degradation and delivering them to endosomal compartments of myeloid or plasmocytoid dendritic cells. Interacting with endosomal toll-like receptors, these complexes induce the production of IFN-α from dendritic cells (DC) and trigger maturation and release of proinflammatory cytokines in myeloid DC.37 Indeed, supernatants from SLE netting neutrophils induced plasmocytoid dendritic cells to synthesise more IFN-α.34 ,36
Summarising these observations, the prevailing idea is that neutrophils by NETosis expose autoantigens to immune cells in the presence of danger signals, but whether this is sufficient for the induction of autoantibodies in a normal or autoimmune background is still to be settled. It has been reported that neutrophils from patients with RA contain an increased amount of deiminated H3 as compared with normal subjects,33 and experiments are in progress to evaluate H4 deimination in RA as compared with controls. As neutrophils infiltrate synovial tissue and are the most abundant cells in synovial fluid, NETosis may create targets for ACPA in rheumatoid synovia, where these autoantibodies are produced.38
The analysis of NET composition in healthy subjects and in patients with RA, and the study of their immunogenic properties, may help to unravel the relationship between NETosis, citrullinated histones and induction of ACPA in patients with RA.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
Handling editor Tore K Kvien
-
Contributors All authors were involved in drafting the article or revising it critically for important intellectual content, and all authors approved the final version to be published. PM and FP had full access to all the data in the study and take responsibility for the integrity of the data and the accuracy of the data analysis. Study conception and design: FP, ID, MCA, PR, PM. Acquisition of data: FP, ID, CT, MCA, IP, FB, FB, FP, IP, PR, PM. Analysis and interpretation of data: FP, ID, MCA, FP, PR, PM.
-
Funding This work was supported by the Italian Ministry of Education, University and Research (MIUR) and by the Fondazione Ente Cassa di Risparmio di Firenze.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval This study was conducted with the approval of the University of Pisa ethics committee.
-
Provenance and peer review Not commissioned; externally peer reviewed.