Article Text

Abstract
Objective To study the characteristics and phenotype of anticitrullinated protein antibody (ACPA)-specific B cells in peripheral blood of patients with rheumatoid arthritis (RA).
Methods Peripheral blood B cells from ACPA-positive patients with RA were cultured with or without stimulating factors. Following culture, supernatants were assessed for the presence of ACPA-IgG and non-specific total IgG by ELISA.
Results Following stimulation, ACPA were detectable in up to 100% of culture wells. Of interest, ACPA were also produced spontaneously by unstimulated peripheral blood mononuclear cells. In both cases, the average ACPA titre per culture well correlated with ACPA serum titres. No ACPA production was detectable in B cell cultures from ACPA-negative patients with RA or healthy controls. Importantly, FACS-sorting experiments located spontaneous ACPA production to the CD20 negative B cell population corresponding to circulating plasmablasts/cells.
Conclusions ACPA-specific peripheral blood B cells are not confined to the CD20 positive memory pool, as circulating plasmablasts/cells spontaneously producing ACPA are also readily detectable. The latter points to an ongoing B cell immune response against citrullinated proteins and contrasts conventional immune responses against, for example, vaccines, where antigen-specific plasmablasts appear in peripheral blood only shortly after vaccination. These circulating, ACPA-specific plasmablasts/cells might represent targets for novel therapeutic interventions.
- Rheumatoid Arthritis
- B cells
- Ant-CCP
Statistics from Altmetric.com
Introduction
In many autoimmune diseases, including rheumatoid arthritis (RA), autoantibodies are believed to be the causative agent that drives disease pathogenesis. Therefore, novel therapeutic approaches that target autoantibody-producing B cells require a thorough understanding of their nature, phenotype and disease-specific localisation.
Several autoantibodies have been described in RA. Among those, anticitrullinated protein antibodies (ACPA) exhibit the highest specificity for the disease, predict disease onset and severity, and identify a subgroup of patients eligible for more aggressive treatment.1–3 Citrullinated antigens are present in the inflamed joints of patients with RA, supporting the concept of ACPA-specific effects at the site of inflammation.4 Indeed, recent evidence suggests a role for ACPA in disease pathogenesis.5–8
So far, little is known on the characteristics of ACPA-producing B cells. A single study detected ACPA production in cultures of synovial fluid mononuclear cells.9 Another study identified ACPA production by peripheral blood B cells upon stimulation.10 However, it remained unclear whether ACPA detected in this latter study originated primarily from stimulated, otherwise resting memory B cells, or from circulating, differentiated plasmablasts/cells (PB/PC).
In antivaccine immune responses, antigen-specific PB transiently appear in peripheral blood in the week after vaccination. These cells rapidly disappear from the circulation and are thought to home to the bone marrow to become long-lived PC that provide high antibody serum titres for years.11 ,12 Like antibody titres against recall antigens, ACPA serum titres in patients with RA are relatively stable and can persist at high titre. Nevertheless, it has been postulated that these high ACPA serum titres in patients with RA are due to a persistently active immune response against citrullinated proteins, as suggested by the continuous presence of IgM ACPA in some ACPA-positive patients.13 If this would be correct, one could predict the presence of ACPA-secreting B cells in peripheral blood. Here, we set out to define ACPA-producing B cells in more detail to increase our understanding of anticitrullinated protein immunity.
Patients and methods
Patients and healthy individuals
Peripheral blood and serum samples were obtained from patients with ACPA-positive and ACPA-negative RA visiting the outpatient clinic of the Department of Rheumatology at Leiden University Medical Centre, Leiden, The Netherlands. Patients fulfilled the 1987 criteria for RA at the time of diagnosis and gave written informed consent for sample acquisition. Treatment included disease-modifying antirheumatic drugs, biological agents, and glucocorticoids. Permission for conduct of the study was obtained from the ethical review board of Leiden University Medical Centre.
Isolation of PBMC, B cells and B cell subsets
Mononuclear cells were isolated from peripheral blood using Ficoll–Paque gradient centrifugation (LUMC pharmacy). CD19-positive B cells were isolated by magnetic bead-based positive selection using Dynabeads and DETACHaBEAD CD19 (Invitrogen) according to the manufacturer's instructions.
For selected experiments, B cells were sorted into subsets on a BD FACS Aria flow cytometer. Peripheral blood mononuclear cells (PBMC) were surface stained with CD3 Alexa Fluor 700 (clone UCHT1), CD14 Pacific Blue (clone M5E2), CD19 APC-Cy7 (clone SJ25C1), CD20 PerCP (clone L27), and CD27 PECy7 (clone M-T271, all BD Biosciences). Subpopulations were defined as CD3−CD14−CD19+CD20−CD27++ PB/PC, CD3−CD14−CD19+CD20+CD27− naïve and CD3−CD14−CD19+CD20+CD27+ memory B cells (figure 3A).14 ,15
Culture conditions
B cells were cultured in Iscove's Modified Dulbecco's Medium (IMDM) supplemented with 10% heat-inactivated fetal calf serum (FCS), penicillin/streptomycin (100 U/ml), and 2 mM Glutamax in 96-wells flat bottom plates at a density of 2×104 cells/well on a layer of irradiated (7000 RAD) mouse fibroblast cells stably transfected with human CD40 ligand (CD40L, 5×103 cells/well) in the presence of B cell activating factor (BAFF) (Miltenyi, 100 ng/ml), IL21 (Invitrogen, 50 ng/ml) and anti-IgM F(ab′)2-fragments (JacksonImmunoresearch Laboratories, 5 µg/ml). Supernatants were harvested after 6–7 days.
To assess spontaneous ACPA production, PBMC were cultured at a density of 2×105 cells/well in IMDM supplemented with 10% heat-inactivated FCS, penicillin/streptomycin (100 U/ml), and 2 mM Glutamax with no additional stimuli. FACS-sorted B cell subsets were cultured under similar conditions, with only variation in cell density (2×104 cells/well for memory/naïve populations; 1500–8000 cells/well for PB/PC) and culture plate (96-well round-bottom plates). Supernatants were collected at day 6 and 13.
Measurement of IgG-ACPA and total IgG
Serum and culture supernatants were assessed for the presence of IgG-ACPA by ELISA based on reactivity against the CCP2-peptide (Euro Diagnostica). Serum samples were tested at a 1:50 dilution or higher according to the manufacturer's instructions, culture supernatants were tested undiluted. The commercial standard was diluted to assess the lowest concentration at which the standard curve was consistently linear (0.125 U/ml (see online supplementary figure S1)). This value takes into account that the concentration of the commercial standard relates to a 1:50 dilution of serum samples, requiring a division by 50 of the manufacturer's units. A representative set of patients (25%) was tested, and found negative, for reactivity against the arginine-containing control peptide.
The presence of total IgG in culture supernatants was assessed by standard ELISA (Bethyl Laboratories).
Statistical analysis
Analysis was performed using GraphPad Prism V.5.01. Correlations were assessed as non-parametric correlation. p Values <0.05 were considered significant.
Results
ACPA production upon in vitro B cell stimulation
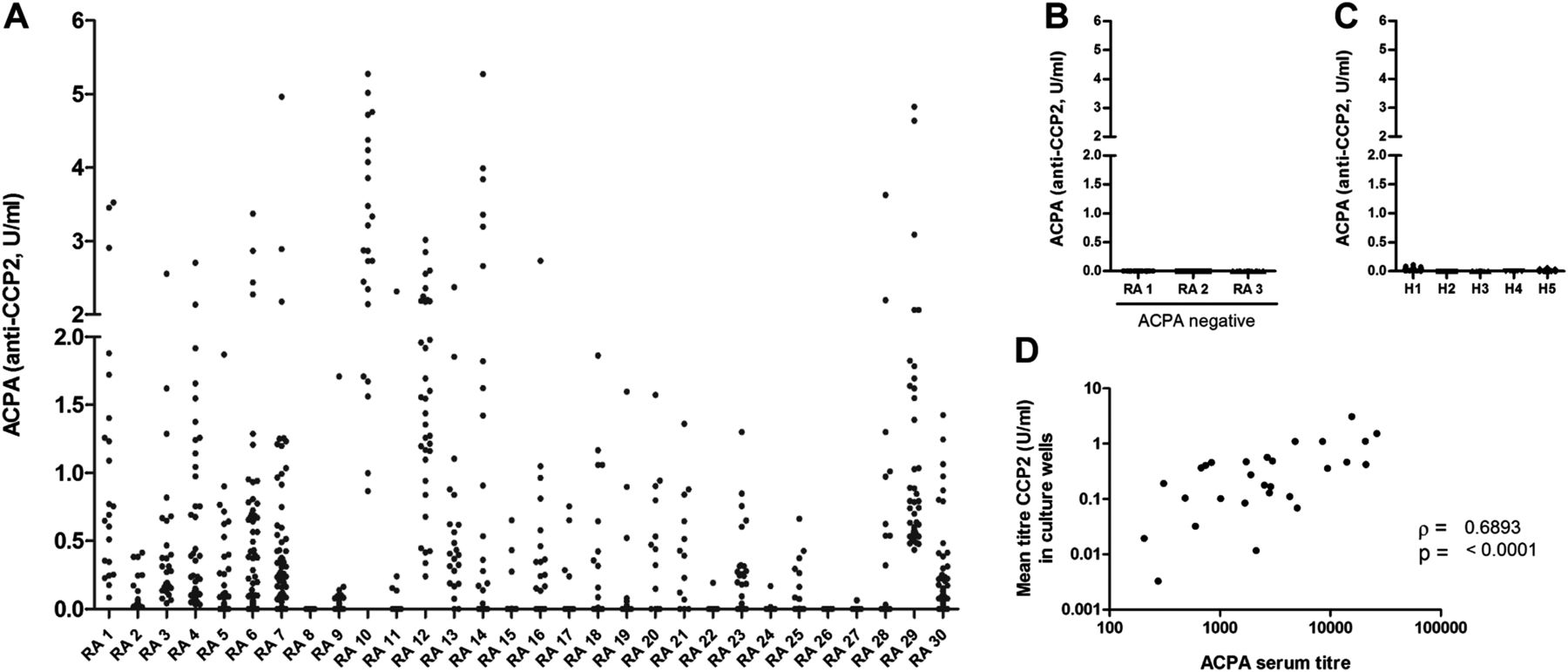
To assess the general presence of ACPA-producing B cells in peripheral blood, we stimulated freshly isolated CD19+ B cells of ACPA-positive patients with RA (n=30) as described previously.16 ,17 After 6–7 days, ACPA were detected in 80% of the cultures and, in some patients, in up to 100% of the culture wells (figure 1A). No ACPA were detected using B cells isolated from healthy individuals and ACPA-negative patients with RA (figure 1B,C), although total IgG production was comparable (not shown). Of interest, mean ACPA titres obtained in the cultures correlated with ACPA serum titres (figure 1D). Based on the number of B cells per culture well, we estimate that, in some patients, at least one in every 2×104 B cells can produce ACPA.
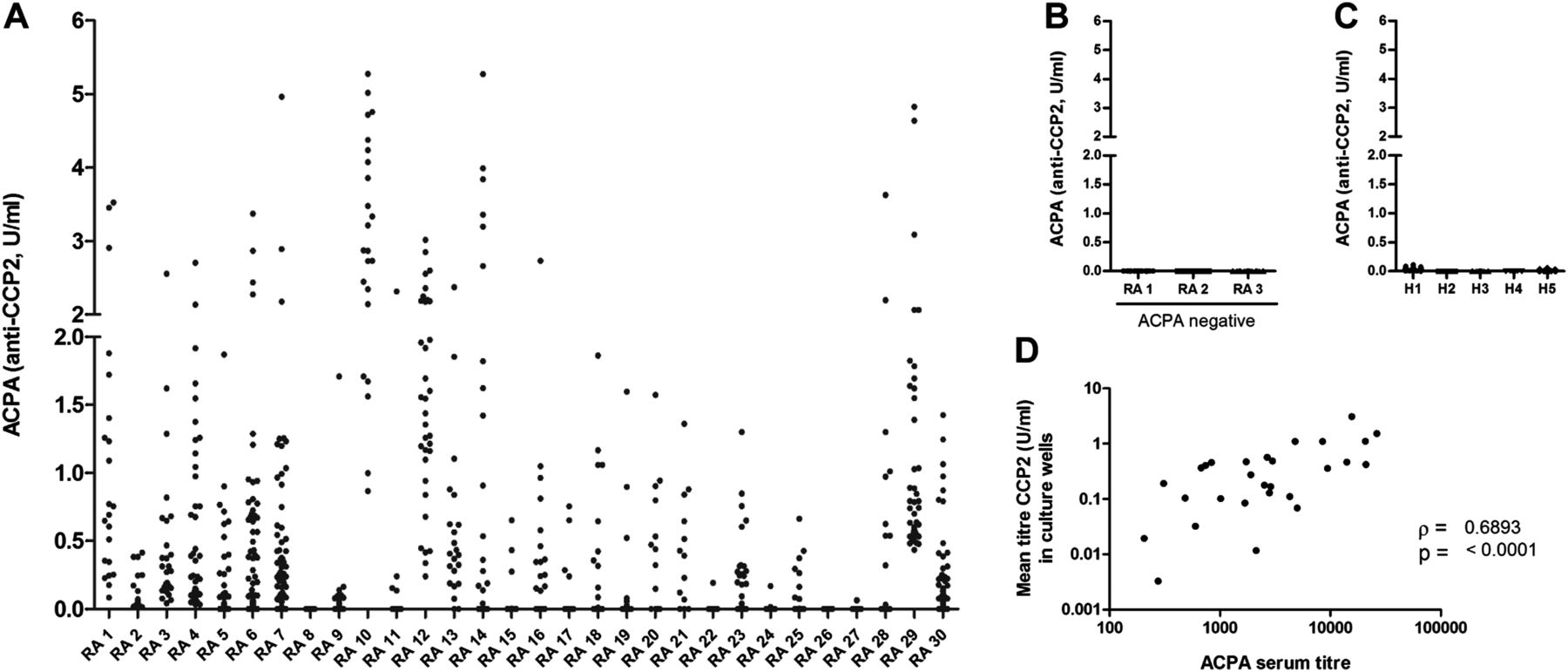
Anticitrullinated protein antibody (ACPA) production upon in vitro B cell stimulation. CD19-positive B cells were isolated and cultured for 6–7 days. Supernatants were assessed for the presence of IgG-ACPA by ELISA. Each dot represents a culture well, results are grouped per patient. Values below linearity of the standard (0.125 U/ml) were calculated by extrapolating the standard curve linear through zero (see online supplementary figure S1). Displayed are results from (A) ACPA-positive rheumatoid arthritis (RA) patients. On average, we obtained 38 (6–170) wells per patient, with a mean ACPA titre of 0.45 U/ml (0.0–3.1)) at the end of the cultures, (B) ACPA-negative patients with RA, n=3; average of 45(16–96) wells per experiment, (C) Healthy donors, n=5; average of 51 (32–73) wells per experiment. (D) Shows the correlation between ACPA serum titres and mean ACPA titres produced in culture.
Spontaneous ACPA production
If the ACPA-response would result from an ongoing immune response, we reasoned that naïve or memory B cells should be continuously activated resulting in the presence of IgG-ACPA producing PB/PC in peripheral blood. Therefore, we analysed whether B cells spontaneously producing ACPA could be observed. To this end, PBMC cells were cultured without exogenous stimuli. Interestingly, comparable amounts of ACPA-positive culture wells were detected in most patients when comparing stimulated versus non-stimulated PBMC per patient (figure 2A), whereas non-specific IgG production was strongly increased upon stimulation (up to 100-fold, data not shown). Importantly, also mean titres of spontaneous ex-vivo ACPA production correlated with ACPA serum titres (figure 2B). This suggests that ACPA detected in the culture wells originate primarily from circulating PB/PC.

Spontaneous versus stimulated anticitrullinated protein antibody (ACPA) production. (A) peripheral blood mononuclear of ACPA-positive rheumatoid arthritis patients were split in two and cultured either with or without stimulation at a density of 2×105 cells/well. Supernatants were assessed for the content of IgG-ACPA at day 13 of culture. (B) Shows the correlation between ACPA serum titres and mean ACPA titres spontaneously produced in culture.
Circulating PB/PC as primary source of ACPA ex vivo
To confirm the presence of ACPA-producing PB/PC in peripheral blood, and to exclude the possibility of endogenous stimulation of memory B cells by antigen-presenting cells within the PBMC population, we next isolated PB/PC by FACS sorting (see figure 3A for gating strategy). ACPA production by purified PB/PC was compared with ACPA production of total PBMC, PBMC depleted of PB/PC, and with PBMC depleted of the CD20 positive B cell fraction (figure 3B,C). In line with our hypothesis, isolated PB/PC readily produced ACPA ex-vivo. The total PBMC fraction and PBMC depleted of CD20-positive cells both showed comparable ACPA production, whereas ACPA production by PBMC depleted of PB/PC was considerably diminished.

{kind=link}
{kind=link}
{kind=link}
Circulating plasmablasts/cells (PB/PC) as primary source of ex-vivo anticitrullinated protein antibody (ACPA) production. CD19-positive B cell subpopulations were FACS-sorted based on the expression of surface markers CD20 and CD27 into CD20−CD27++ PB/PC, CD20+CD27+ memory B cells and CD20+CD27− naïve B cells (A). In addition, the markers were used to deplete peripheral blood mononuclear cells (PBMC) of the respective B cell subset. Sorted B cell populations and PBMC depleted of subsets were cultured without stimulation. (B) Representative data of one patient. Each dot corresponds to one culture well. (C) Combined data of five patients. Each dot represents one donor. The lowest point of linearity of the standard was used as cut-off for ACPA positivity. (D) Naïve and memory B cells were FACS-sorted as described above for four patients. Both populations were stimulated as described in the Methods section. Supernatants were harvested after 6 days of culture and assessed for ACPA by ELISA. Data are depicted as units corrected for culture volume.
ACPA were not produced by unstimulated, FACS-sorted memory B cells (not shown), but were detectable upon stimulation. This indicates the additional presence of CD20-positive, ACPA-specific memory cells in the circulation (figure 3D). ACPA were hardly detected in wells containing naïve B cells, irrespective of stimulation.
Together, these data highlight the presence of circulating, ACPA-producing PB/PC in peripheral blood of patients with RA, and indicate that depletion of the CD20-positive B cell compartment does not eliminate these cells.
Discussion
The immune response against citrullinated antigens in RA is incompletely understood. Specifically, characteristics of ACPA-producing B cells have scarcely been studied. Here, we show that ACPA-producing PB/PC circulate in peripheral blood of patients with RA.
In the steady state, most circulating PB/PC in healthy individuals are thought to originate from mucosal immune responses.11 Upon vaccination, antigen-specific PB appear in the circulation only transiently. Therefore, our findings provide evidence for the recent and continuous activation of ACPA-producing B cells in RA patients. This observation is relevant, as recently activated PB/PC are phenotypically different from fully differentiated, long-lived plasma cells or memory B cells. Indeed, depletion of the CD20-positive B cell compartment did not eliminate the spontaneous ACPA production observed.
Next to circulating PB/PC, we identified ACPA-specific memory B cells that produced ACPA upon stimulation. The latter observation could be taken as a surprise as it is conceivable that citrullinated protein-directed memory B cells are constantly driven to the PB/PC population as a consequence of the persistent and systemic presence of citrullinated antigens. Novel therapeutic approaches aimed at inhibiting ACPA production should thus target the CD20-positive B cell subset, but need to also include the PB/PC compartment.
Finally, we found an association between ACPA production in culture and ACPA serum titres. This suggests that ACPA serum titres originate partly from tissue resident, long-lived plasma cells producing rather stable ACPA ‘background’ titres, and partly from recently generated, circulating PB/PC. The latter reflect the active part of the immune response and are thus more variable in their contribution to the ACPA serum titre, as suggested by our study. In line with this, intensive treatment was previously shown to be able to decrease ACPA serum titres by up to 50%,18–20 suggesting indeed a fluctuation in ACPA titres due to an ongoing immune response.
In conclusion, we here identify ACPA-specific PB/PC circulating in peripheral blood of ACPA-positive patients with RA. This population, which is not targeted by CD20 depletion, reflects the active part of the ACPA-specific B cell response and could represent a relevant target for therapeutic intervention.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figures
Footnotes
-
Contributors PFK performed and designed experiments, analysed data and drafted the article. YR designed experiments and critically revised the manuscript for important intellectual content. EIHvdV performed and designed experiments and analysed data. LAT interpreted data, designed experiments and critically revised the manuscript for important intellectual content. TWJH interpreted data, drafted the article and critically revised the manuscript for important intellectual content. REMT interpreted data, drafted the article, designed experiments and critically revised the manuscript for important intellectual content. HUS designed experiments, analysed and interpreted data and drafted the article. All authors approved the final version of the manuscript.
-
Funding This work was supported by grants from the Dutch Arthritis Foundation, the European Union (Seventh Framework Programme integrated project Masterswitch, grant number: 223404) and the IMI JU funded project BeTheCure (contract no 115142–2). LAT received a VIDI grant and REMT a VICI grant from NWO-ZonMW, The Netherlands.
-
Competing interests None.
-
Ethics approval Ethical Review Board of Leiden University Medical Centre.
-
Provenance and peer review Not commissioned; externally peer reviewed.
-
Data sharing statement Additional unpublished data are available to the authors as mentioned in the manuscript text.