Article Text

Abstract
Objective To determine the potential clinical and pathological significance of altered expression of interleukin 6 (IL-6) in systemic sclerosis (SSc).
Methods Serum IL-6 and soluble IL-6 receptor levels were measured in patients with SSc (n=68) and healthy controls (n=15). Associations between serum IL-6 level and C reactive protein, platelet count and key clinical outcomes in SSc were explored. Expression of IL-6 in skin biopsies was also examined and western blot and reverse transcription PCRanalysis were performed using cultured dermal fibroblasts. The effect of IL-6 trans-signalling on production of extracellular matrix proteins was assessed and downstream signalling pathways were examined using pharmacological inhibitors.
Results Serum IL-6 level was frequently elevated in patients with SSc, particularly in those with diffuse cutaneous SSc (dcSSc) with thrombocytosis and elevated acute phase markers. Prominent expression in the skin was observed in dermal fibroblasts, mononuclear cells and endothelial cells in patients with early dcSSc. In vitro experiments supported a potent profibrotic effect of IL-6 trans-signalling via the JAK2/STAT3 and ERK pathways. High IL-6 expression early in dcSSc appears to be associated with more severe skin involvement at 3 years and worse long-term survival than in those without elevated IL-6 levels.
Conclusion Our results confirm the overexpression of IL-6 in dcSSc and support the potential of IL-6 as a surrogate marker for clinical outcome in this disease. The data also provide rationale for clinical studies targeting IL-6 trans-signalling as a potential antifibrotic therapy for SSc.
Statistics from Altmetric.com
Systemic sclerosis (scleroderma, SSc) is an autoimmune connective tissue disease with high mortality. Previous reports have associated several laboratory variables with increased mortality, including elevated erythrocyte sedimentation rate, anaemia, thrombocytosis and proteinuria.1 2 Although poorly understood, SSc pathogenesis appears to involve immunological abnormalities with autoantibody production, vascular alterations and increased accumulation of extracellular matrix (ECM) proteins in the skin and internal organs. Interaction of fibroblasts with other cell types, and cytokines that modulate key regulatory pathways controlling synthesis of matrix components, are likely to be central to clinical progression.
IL-6 is a pleiotropic cytokine that regulates cellular proliferation, activation and differentiation of numerous cell types by binding to the IL-6 receptor (IL-6R, gp80) on the cell surface in association with gp130. Neither IL-6R nor IL-6 binds to gp130 alone. The gp130 receptor is expressed on multiple cell types, but IL-6R is restricted to hepatocytes and leucocytes. However, soluble IL-6R (sIL-6R) together with IL-6 can stimulate cells lacking gp80, a process termed trans-signalling.3 As IL-6 interacts with its receptor gp80, it triggers the gp130 and IL-6R proteins to form a complex, thus activating the receptor. These complexes bring together the intracellular regions of gp130 to initiate a signal transduction cascade through Janus kinases (JAKs) and signal transducers and activators of transcription (STATs), but may also lead to an activation of MAP-kinase (MAPK) signalling cascades.4 IL-6 trans-signalling has now been implicated in a number of pathological processes including rheumatoid arthritis and Crohn's disease.
Numerous reports have shown that IL-6 levels are elevated in culture supernatants of dermal fibroblasts and serum from patients with SSc.5 6 The role of IL-6 in regulation of dermal fibrosis, however, is less clear. IL-6-deficient transgenic mice display delayed cutaneous wound healing and reduced collagen deposition.7 8 However, Mihara et al demonstrated that IL-6 trans-signalling induced proliferation of synovial fibroblast but has the opposite effect on dermal fibroblasts.9 10 In the present study, we explore expression of IL-6 in the serum and skin in defined subsets of SSc and explore a potential role in promoting ECM production and its association with clinical outcomes in SSc.
Patients and methods
Patients and controls
Thirty-nine patients with diffuse cutaneous systemic sclerosis (dcSSc), 29 patients with limited cutaneous systemic sclerosis (lcSSc) and 15 healthy controls were recruited for the study. Thrombocytosis was defined by platelet counts above the upper normal limit with a mean of 450×109/l. Twenty patients with persistent thrombocytosis on at least three occasions were identified during consecutive routine clinic attendances for their disease assessment between 2008 and 2010. Age- and sex-matched patients with dcSSc and lcSSc and healthy controls were selected during the same period. Patterns of organ involvement at the time of biopsy were assessed according to current clinical practice. Briefly, oesophageal involvement was determined by history, barium swallow or scintigraphy; lung fibrosis by high resolution CT scanning in the presence of a restrictive pattern of pulmonary function test abnormality; cardiac impaired ventricular function or significant pericardial effusion on echocardiography. Skeletal muscle involvement was defined by an increase of more than fourfold in the creatinine kinase level. Renal involvement was identified by previous scleroderma renal crisis or significant impairment of creatinine clearance. In addition, lesional dermal punch biopsy samples were obtained from the forearm of 10 patients with early dcSSc, 8 patients with established dcSSc with disease duration of greater than 36 months, 12 patients with lcSSc and 12 age- and site-matched healthy controls for histological study. The study was approved by our institutional local research ethics committee (Royal Free and Medical School Local Research Ethics Committee).
Measurement of serum IL-6 and sIL-6R levels
Serum IL-6 and sIL-6R levels were determined in SSc patients and healthy controls by a validated solid-phase sandwich enzyme immunoassay kit (Roche Diagnostics, Welwyn Garden City, UK). The IL-6 assay measures free IL-6 and the sIL-6R assay measures free sIL-6R and sIL-6R bound to IL-6. The minimum detection limit of the assay for IL-6 was 3.12 pg/ml. All standards and samples were run in triplicate.
Immunohistochemical staining of serial sections
Formalin fixed 3 μm paraffin sections were used for immunostaining. Endogenous peroxidase was blocked using 3% hydrogen peroxide. Optimally diluted anti-IL6 antibody (Novocastra, Newcastle, UK), gp130 (Santa Cruz, CA, USA) polyclonal antibody and anti-CD126 IL-6R α chain (Millipore, Abingdon, UK)were used in this series. Specificity of staining was confirmed in control sections incubated with an isotype-matched irrelevant control antibody. Sections were viewed and photographed using a Zeiss Axioscope brightfield microscope (Carl Zeiss, Germany) using Axiovision software. Staining intensity was assessed using semiquantitative assessment of epidermal, dermal and perivascular staining by an experienced observer, blinded to sample identity during scoring.
Fibroblast cultures and western analysis
Dermal fibroblasts were cultured from six early dcSSc patients with similar number of age- and site-matched control biopsies in DMEM, 10% fetal bovine serum (Invitrogen, Paisley, UK) as previously described.11 Cells (∼80% confluence) were serum-starved overnight, lysed in 2% SDS, and proteins were quantified (Pierce, Northumberland, UK) and subjected to western blot analysis as previously described.11 Antibodies used were: goat antihuman IL-6 antibody (R&D, Abingdon, UK), anti-α-smooth muscle actin (αSMA) (Sigma, Suffolk, UK), anti-GAPDH (glyceraldehyde-3-phosphate dehydrogenase) (Sigma), anti-CTGF (connective tissue growth factor) (Abcam, Cambridge, UK), antitype I collagen (Southern Biotechnology, Birmingham, Alabama, USA), monoclonal and polyclonal antibodies against phosphorylated-ERK1/2, ERK1/2, phosphorylated-JAK2, JAK2, phosphorylated-STAT3, STAT3 and gp130 (Cell Signalling Technology, Danvers, MA, USA). AG490 and S3I-301 were obtained from Calbiochem (San Diego, California, USA). ERK inhibitor, U0126 was obtained from Promega (Madison, Wisconsin, USA).
Quantitative reverse transcription PCR
Total RNA was extracted from confluent skin fibroblasts from patients with SSc and healthy controls using the RNeasy mini-kit (Qiagen, Crawley, UK) according to the manufacturer's instructions and quantified using the Nanodrop ND-8000 spectrophotometer (Thermo-Scientific, Wilmington, Delaware, USA). The minimum 260/280 ratio was 1.98. RNA integrity numbers ranged from 9.1 to 10, measured on an Agilent 2100 Bioanalyzer (Agilent Technologies UK Limited, Stockport, UK). One microgram of RNA was reverse transcribed using the Quantitect reverse transcription kit (Qiagen) and diluted fivefold with 0.2 ug/ml tRNA. The real-time quantitative reverse transcription PCR used 2 μl RNA in a 10 μl reaction volume using Sensimix SYBR (Quantace, London, UK) on a Rotorgene-6000 (Corbett Life Sciences, Sydney, Australia) under the following conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 57°C for 10 s, 72°C for 5 s. Specific products and absence of primer dimers were confirmed by melt curve analysis. Copy number and assay efficiency were derived from known copy number standard curves. Three stable reference genes: SDHA, RPL13 and HPRT1 were identified using geNorm and copy numbers were corrected using the computed normalisation factor.12 Primer sequences, written 5′–3′, are referenced where appropriate, assay efficiency and R2 are as follows: SDHA fwd tgggaacaagagggcatctg, rev ccaccactgcatcaaattcatg, 0.82, 0.999; RPL13 fwd cctggaggagaagaggaaagaga, rev ttgaggacctctgtgtatttgtcaa, 0.82, 0.998; HPRT1 fwd tgacactggcaaaacaatgca, rev ggtccttttcaccagcaagct, 0.83, 0.999; IL-6 fwd acagccactcacctcttcag, rev ccatctttttcagccatcttt, 0.96, 0.999.
Statistical methods
Fisher's exact test was used to compare proportions of subjects with high/low/undetectable IL-6 levels and Kruskal–Wallis test was used to compare IL-6R and C reactive protein (CRP) levels and modified Rodnan skin score (mRSS)in the different subgroups. Spearman's rank correlation coefficient was used to show correlation between IL-6/sIL-6R levels and acute phase reactants and mRSS. Log-rank test was used to compare Kaplan–Meier estimates of survival in the groups of high and low IL-6.
Results
Elevated serum IL-6 levels in patients with SSc
To examine if serum IL-6 and sIL-6R are upregulated in SSc subsets, we measured the levels in 20 patients with dcSSc and thrombocytosis, 19 patients with dcSSc and normal platelets, 29 patients with lcSSc and 15 healthy controls. Cohort characteristics are summarised in table 1.More than half of the patients with dcSSc harboured either anti-Scl70 or anti-RNA polymerase antibodies. Forty-nine per cent (n=19) of the patients with dcSSc developed lung fibrosis of which a majority (68%) harboured the anti-Scl70 antibody. Anticentromere antibody was detected only among patients with lcSSc, although two patients with lcSSc had the anti-Scl70 antibody and both had developed lung fibrosis. IL-6 levels varied between 3.23 and 51.3 pg/ml (mean 14.39, median 10.1, IQR 4.46–20.5 pg/ml). Thirty-five of the SSc patients and 13 of the controls had IL-6 levels below the lower level of detection. IL-6R levels varied between 18.6 and 75 pg/ml (mean 38.82, median 37.3, IQR 29.7–45.9 pg/ml) and CRP levels were between 4 and 80 (mean 11.3, median 7, IQR 5–12).
Demographic and clinical features of study cohort
We compared proportions of subjects with high (above median), low (below median and detectable) and undetectable levels of IL-6 in the SSc and control groups. As IL-6 has a potent stimulatory effect on thrombopoiesis via induction of thrombopoietin,13 we also explored whether a subgroup of patients with dcSSc with elevated platelet counts may have higher levels of IL-6. We found a statistically significant difference among the different subgroups, with high levels of IL-6 observed in 55% of the dcSSc cohort with thrombocytosis compared with 16% of the dcSSc cohort with normal platelets, 7% of the lcSSc subjects and 7% of the controls, while undetectable levels were seen in 20% of the dcSSc cohort with thrombocytosis compared with 47% of the dcSSc cohort with normal platelets, 76% of the lcSSc subjects and 87% of the controls (p<0.001). Figure 1A demonstrates median IL-6 levels in the different SSc subgroups and in control patients. There were no statistically significant differences in the median (IQR) levels of serum IL-6R across all SSc subgroups and controls: 35 (27–49) pg/ml for dcSSc with thrombocytosis, 35 (26–42) pg/ml for dcSSc with normal platelets, 40 (35–47) pg/ml for lcSSc and 38 (31–47) pg/ml in healthy controls (p=0.22).

Serum IL-6 levels in patients with systemic sclerosis (SSc) and healthy controls. (A) Serum samples were obtained from patients with diffuse cutaneous SSc (dcSSc; n=39 from which 20 patients have thrombocytosis), limited cutaneous SSc (lcSSc; n=29) and healthy controls (n=15). Serum IL-6 levels were measured by an enzyme immunoassay. For each subgroup, the median values are shown adjacent to the horizontal grey bars. The minimum detection limit of the assay for IL-6 was 3.12 pg/ml as indicated by a dotted line and patients who have undetectable levels of IL-6 with this assay were represented on this line. (B,C) Serum IL-6 levels were correlated with C reactive protein (CRP) and platelets in patients with SSc (n=68).
Correlation of serum IL-6 and sIL-6R levels with CRP and platelets
It is known that IL-6 is potent stimulant for synthesis of CRP as an acute phase reactant by the liver in response to various inflammatory processes. As expected, serum IL-6 levels positively correlated with CRP (Spearman's ρ=0.60, p<0.001, figure 1B). In addition, IL-6 levels correlated moderately well with platelet count (Spearman's ρ=0.38, p=0.002, figure 1C). There was no correlation between serum sIL-6R levels and CRP (Spearman's ρ=−0.128, p=0.251) or platelet count (Spearman's ρ=−0.18, p=0.142).
Immunostaining for IL-6 in SSc and healthy control skin
To further explore the potential pathological significance of IL-6 expression in SSc, a subset of the cases of SSc that were included in the study of serum levels were evaluated by staining for IL-6 expression in skin biopsies. Four distinct patterns of strong IL-6 ligand staining were observed (figure 2). Control isotype matched IgG antibody was used to confirm specificity of staining (data not shown). The most frequent pattern of localisation was strong epidermal staining specifically of keratinocytes, melanocytes and resident Langerhans cells in 37/42 subjects. This did not show any association with disease subset in 9/12 control, 10/12 lcSSc and all dcSSc samples. In 7/10 early dcSSc samples there was strong ligand expression of fibroblasts from the papillary dermis but not in the deeper dermis layers. Interestingly, this pattern was not observed in any other group (figure 2E). In the majority of skin sections from patients with early dcSSc (8/10), IL-6 was localised to perivascular inflammatory infiltrates consisting of lymphocytes and macrophages based on morphological criteria (figure 2F).Prominent endothelial staining for IL-6 was also detected in representative sections from each group but this was more frequent in cases of early dcSSc (8/10) and less often in established dcSSc (2/8) (figure 2F). It was, however, also observed in 4 of 12 healthy control biopsies and 6 of 12 lcSSc subjects. Similar to the constitutive epidermal expression for IL-6 ligand in all SSc and healthy controls, no differences were observed in the gp130 expression in the dermis (figure 2G,H). Preliminary results from histological analysis on IL-6R expression suggest that there was epidermal staining for both early dcSSc and healthy controls with no differences in the dermal expression (data not shown).
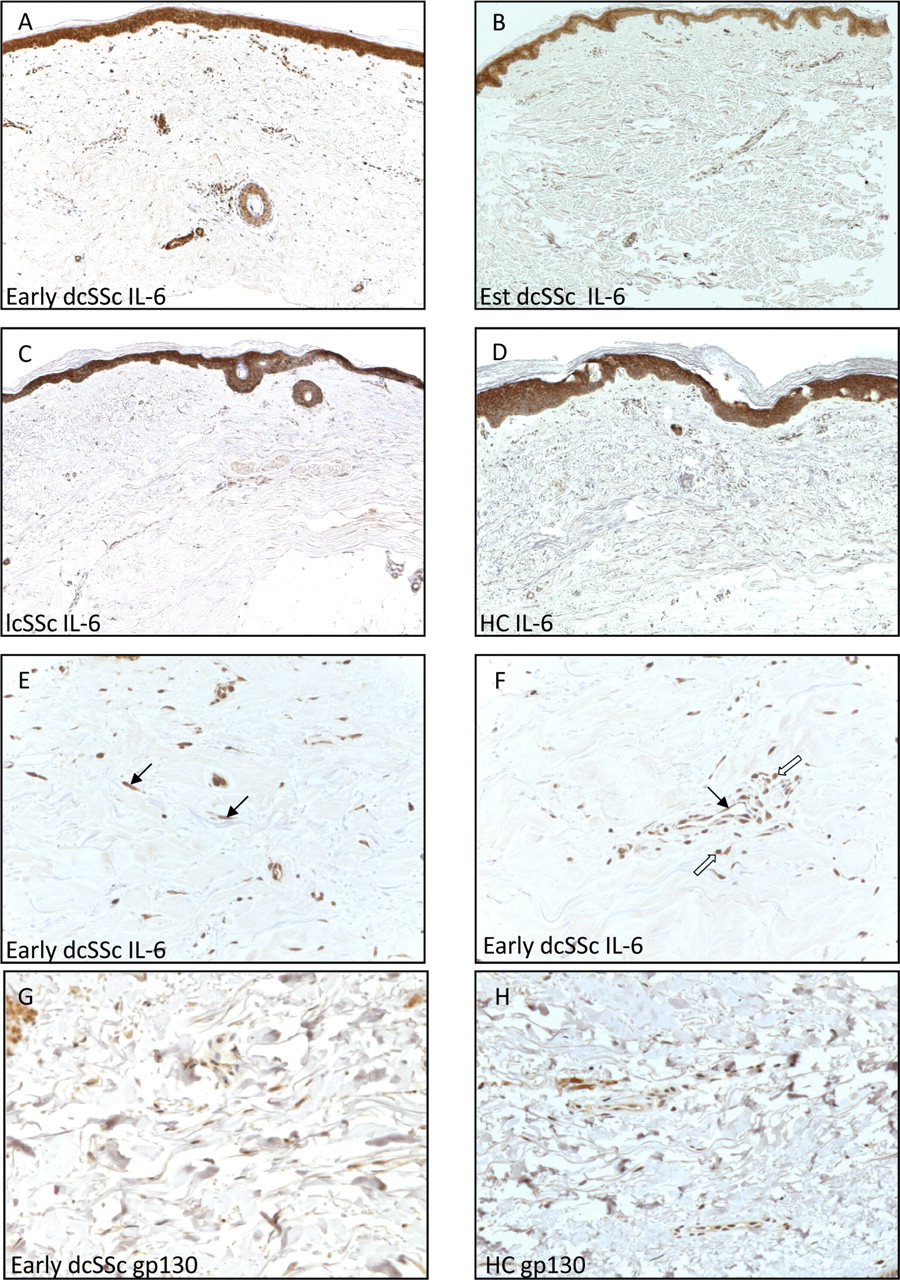
Increased dermal expression of IL-6 in systemic sclerosis (SSc). Expression of IL-6 in the skin sections from patients with early stage and established diffuse cutaneous SSc (dcSSc), limited cutaneous SSc (lcSSc) and healthy controls (HCs) (A–D). Paraffin sections of representative skin sections were immunostained with specific antibody for human IL-6. Original magnification ×50. (E,F) Increased abundance of IL-6 in skin sections from patients with early dcSSc. Distinct staining patterns for IL-6 were observed with spindle-shaped fibroblastic expression (arrows in E), endothelial localisation (arrow in F) with distribution of perivascular mononuclear inflammatory infiltrates (open arrows in F). (G,H) Immunostaining for gp130 protein in skin sections from patients with early dcSSc and control. Original magnification ×200. Est, established.
IL-6 mRNA and protein expression in dermal fibroblasts of SSc patients
To determine whether IL-6 is involved in the fibrotic response in SSc, we used reverse transcription PCR to show that IL-6 mRNA levels were increased in cultured SSc dermal fibroblasts compared with healthy controls (11.8±3.3×105 copies/ulin SSc fibroblasts compared with 1.7±0.06×105 copies/ulin healthy fibroblasts, p=0.002, n=3) (figure 3A). IL-6 protein was elevated in the supernatants from cultured SSc fibroblasts (n=6) (4.27±0.69 compared with 0.25±0.31 densitometric image unit (DIU) healthy fibroblasts, p<0.05) (figure 3B). This was associated with increased collagen production in SSc fibroblasts (19.41±3.96 compared with 2.58±1.36 DIU healthy fibroblasts, p<0.05).

Overexpression of IL-6 mRNA and extracellular matrix protein by lesional systemic sclerosis (SSc) skin fibroblasts and the effect of IL-6 trans-signalling on fibroblasts. (A) Reverse transcription PCR semiquantitative mRNA measurement of IL-6 in cultured SSc and normal fibroblasts. Data are representative of three independent experiments examining three patients and three healthy controls, respectively. Results are expressed as mean copy numbers±SEM, * p<0.05. (B) Fibroblast culture media from representative normal or SSc dermal fibroblasts cultures were analysed for protein expression using antibody to IL-6 and Col-1. GADPH staining of cell layer lysates was used as a loading control. Densitometric analysis summarising three independent experiments shows the mean protein levels for IL-6 and Col-1. *p<0.05. (C) Stimulation of cultured normal dermal fibroblasts with IL-6 and soluble IL-6 receptor (sIL-6R) demonstrate a dose-dependent increase in Col-1, α-smooth muscle actin and CTGF synthesis. Data represent mean±SEM of triplicate measurements in each group. * p<0.01 and †p<0.05. NF, normal fibroblast; SScF, scleroderma fibroblast.
Effect of IL-6 trans-signalling on fibroblasts
Previous studies have demonstrated that IL-6 may have a profibrotic effect but the role of IL-6/sIL-6R trans-signalling in ECM synthesis is unclear. As collagen was significantly upregulated in SSc fibroblasts (figure 3B), we explored whether the IL-6 ligand-receptor axis may promote ECM production in dermal fibroblasts. As shown in figure 3C, incubation of isolated dermal fibroblasts from healthy controls with either IL-6 (25–50 ng/ml) alone or sIL-6R (20 ng/ml) alone had no appreciable effect on collagen, αSMA and CTGF production. However, when variable concentrations of IL-6 were added with sIL-6R together to the cultured fibroblasts, there was a significant concentration-dependent increase in all three ECM proteins (figure 3C; p<0.05). There was maximal 3.5-fold elevation in collagen expression with 25 ng/ml IL-6 and 20 ng/ml sIL-6R. Similar induction of αSMA and CTGF by 12-fold and 15-fold, respectively, was observed in normal fibroblasts. At the highest concentration of IL-6 (50 ng/ml) with sIL-6R (20 ng/ml), there was a sustained enhanced αSMA expression but a reduced expression of collagen and CTGF in normal fibroblasts.
IL-6/sIL-6R enhances ECM synthesis through a JAK2/STAT3- and ERK-dependent mechanism
To delineate downstream signalling mechanism through which IL-6 trans-signalling promotes ECM synthesis in fibroblasts, we showed that IL-6 trans-signalling activated JAK2, STAT3 and ERK pathways in dermal fibroblasts (figure 4A). The induction of pJAK2 and pSTAT3 peaked at 45 min and this was diminished by 2 h in normal fibroblasts. Similar activation of ERK was observed in normal fibroblasts. Constitutive activation of both JAK2 and STAT3 pathways was observed in SSc fibroblasts and further activation by the addition of IL-6 and sIL-6R occurred at 15 min and this was sustained at 1 h. By contrast, addition of IL-6 and sIL-6R to SSc fibroblasts did not increase the constitutive activation of ERK pathway. Consistent with the immunohistochemical data on dermal gp130 expression (figure 2G,H), there was no difference in the expression of gp130 in cultured fibroblasts lysate from early dcSSc and healthy controls. To address the contribution of these pathways to the IL-6/sIL-6R induction of ECM proteins, a 45-minute preincubation with the JAK2 inhibitor, AG490 (25 µM), STAT3 inhibitor, S3I-301 (50 µM) and MEK/ERK inhibitor, UO126 (10 µM),14 15 prior to the addition of IL-6 and sIL-6R in normal dermal fibroblasts, abrogated the induction of collagen, αSMA and CTGF (figure 4B). While the addition of IL-6 with sIL-6R had no discernible effect on ECM production in SSc fibroblasts, AG490, S3I-301 and UO126 reduced the induction of ECM proteins in the SSc dermal fibroblasts. Thus, the IL-6 trans-signalling activation of ECM protein expression in normal dermal fibroblasts occurs through a JAK2/STAT3- and ERK-dependent mechanism and these signalling pathways appear to be relevant in the induction of key profibrotic elements in SSc fibroblasts.

Activation of extracellular matrix protein synthesis by IL-6 trans-signalling via JAK2/STAT3 and ERK signalling pathways in dermal fibroblasts. (A) Activation of JAK2/STAT3 and ERK by IL-6 and sIL-6R in dermal fibroblasts. Phosphorylation of JAK2/STAT3 and ERK in normal fibroblasts was determined by immunoblotting using antibodies specific for phosphorylated, activated forms of the above antibodies. Dermal fibroblasts were serum-starved overnight and treated with IL-6 (25 ng/ml) and soluble IL-6 receptor (sIL-6R) (20 ng/ml) for the indicated times. Antibodies against JAK2, STATs and ERK were also used to confirm that these protein concentrations were maintained. This is a representative of three independent experiments. (B) Cultured fibroblasts were stimulated overnight with recombinant IL-6 and sIL-6R in the presence of AG490 and S3I-201 that targets JAK2 and STAT3 pathways, respectively. U0126 was also added prior to overnight stimulation to block ERK signalling pathways in the cultured fibroblasts. Equal amounts of total cell lysates and supernatants were resolved in SDS-PAGE and subjected to western blot analyses with antibodies as indicated. Densitometric analysis from three independent experiments depicts the mean protein levels in each group. *p<0.05 and †p<0.01. JAK, Janus kinase; NF, normal fibroblast; SScF, scleroderma fibroblast; STAT, signal transducer and activator of transcription.
Relationship of serum IL-6 expression and clinical outcomes in dcSSc
We investigate the association between circulating levels of IL-6 and key clinical outcomes in SSc. Among patients with dcSSc with available mRSS at the time of presentation, serum IL-6 levels positively correlated with mRSS (Spearman's ρ=0.514, p=0.001, figure 5A). There was also a moderate correlation between peak skin score and serum IL-6 levels at the time of presentation (Spearman's ρ=0.374, p=0.027). We also explored whether serum IL-6 levels may predict the changes in mRSS over the first 3 years of disease. Serum IL-6 level at presentation positively correlated with mRSS at 36-month follow-up in a subgroup of dcSSc cases (Spearman's ρ=0.795, p<0.001, n=16, figure 5B). In addition, high serum level of IL-6 (≥10.1 pg/ml) at presentation of the disease appears to predict higher mortality in dcSSc with a 15-year survival of 30% among these patients compared with 93% in the group with low IL-6 levels (p=0.021) (figure 5C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Clinical significance of serum IL-6 levels in diffuse cutaneous systemic sclerosis. (A) Association of serum IL-6 levels with skin score at the time of disease presentation. (B) Correlation of serum IL-6 levels at baseline with skin score at 36 months. (C) Kaplan–Meier survival curves for 39 patients with systemic sclerosis, by serum IL-6 at entry into the study. Graphs are shown for those with high IL-6 (n=24) (continuous line) and low IL-6 (n=24) (dotted line) levels. Number of patients at risk listed below graphs at 5-year intervals. mRSS, modified Rodnan skin score.
Discussion
This study supports the role of IL-6 trans-signalling in induction of dermal sclerosis via direct activation of dermal fibroblasts. In addition, elevated serum IL-6 levels may identify a subgroup of dcSSc cases with poor outcome in terms of skin involvement at 3 years from disease onset and survival. Our results also confirm that thrombocytosis selectively identifies a cohort of dcSSc patients with high serum IL-6. Recent studies highlighted the role of platelet activation in SSc and thrombopoietin regulation by IL-6 in reactive thrombocytosis.13 16 17 Moreover, thrombocytosis at baseline was associated with high skin score at presentation in a large UK observational cohort of early dcSSc.14 Immunohistochemical analysis demonstrated that IL-6 was differentially expressed in affected tissues in early dcSSc with increased expression, based on morphological criteria, in the fibroblasts and mononuclear inflammatory infiltrates. This contrasts with early reports suggesting higher IL-6 in late stage disease but the small numbers of patients may account for the discrepancy.15 There was, however, no difference in gp130 expression between early active dcSSc and healthy controls as the ubiquitously expressed gp130 protein is believed to be relatively constant for all cells.
Previous studies have also shown that IL-6 induces ECM protein synthesis by fibroblasts.18 In addition, strategies to block the IL-6 response resulted in a significant reduction of procollagen type I in cultured SSc fibroblasts and myofibroblastic differentiation in dermal fibroblasts in bleomycin-induced model of dermal sclerosis.19 20 Interestingly, there was a selective downregulation of collagen and CTGF in response to the increased concentration of IL-6 (50 ng/ml) and sIL-6R (20 ng/ml). This may be related to activation of the negative feedback mechanism, SOCS3 in regulation of the fibrotic response.7 21
It has been suggested that IL-6 bioactivity depends upon trans-signalling mediated via JAK-STAT pathway. Barnes et al recently showed that SSc serum, in the presence of neutrophils as a possible source of IL-6 receptor, induced activation and apoptosis of endothelial cells in an IL-6 dependent manner, supporting the role of trans-signalling in SSc.22 Although elevated levels of sIL-6R have been reported in numerous inflammatory disease states,23 24 our study did not demonstrate increased serum level of sIL-6R in SSc. It is likely that the local source of sIL-6R may be relevant and may contribute to the trans-signalling that drives the local inflammatory and fibrotic response. Support for the enhanced local IL-6 trans-signalling response has been described in wound healing response.25 26 Similarly, activation of the JAK/STAT pathway is involved in several experimental models and human fibrotic diseases.27 In addition, we showed that IL-6 trans-signalling activates MAPK-related kinase ERK, which may partly explain constitutive ERK1/2 activation reported in SSc.11 28 29
There are some important limitations of the present study. Most patients with SSc even those in the early stage disease do not have thrombocytosis. In this study, 10 of the entire diffuse cohort had thrombocytosis with disease duration of less than 3 years. Conversely, thrombocytosis was observed in 10 of the 21 patients with established disease. This may therefore limit the generalisability of our results to other SSc patients. Moreover, the cohort is too small to evaluate the predictive power of IL-6 for SSc-specific organ complications including pulmonary arterial hypertension or renal crisis. Furthermore, serial change of IL-6 levels was not examined during follow-up period. It is possible that the prognostic potential of IL-6 may be enhanced by serial measurement of its levels. Recent study has shown that repeated assays of serum IL-6 levels can be used to examine the association between IL-6 and coronary heart disease.30 Further work is in current progress to evaluate expression of signalling intermediates including receptor expression in lesional tissues. Our study did not specifically address the impact of soluble form of gp130 (sgp130) that selectively inhibits IL-6 trans-signalling without affecting the classical pathway. However, it remains unclear if targeting the trans-signalling with this approach offers an advantage to the current therapies against IL-6R.
Serum IL-6 levels have been shown to correlate with severity of inflammatory diseases such as rheumatoid arthritis and it is associated with adverse prognosis in cardiovascular diseases and sepsis.31,–,34 In our study, we observed that high IL-6 levels at the time of disease presentation are associated with high skin score not only at baseline but also at 36-month follow-up in a subgroup of patients with early dcSSc. Moreover, high levels of IL-6 portend a poor survival. Recent studies revealed that serum IL-6 levels may predict risk of relapse in inflammatory bowel diseases and cardiovascular events.31 32 Our data also support the observation of Shand et al that patients with worse skin trajectory with high baseline mRSS with no improvement in mRSS had poor survival.35 It is also established that severe skin disease and elevated acute phase proteins are associated with high mortality.36
In conclusion, our findings extend earlier work on IL-6 in SSc and define a subset of cases in which IL-6 expression and function may be especially relevant. Since there are now specific inhibitors of IL-6 receptor such as tocilizumab, these may be promising potential therapeutic agents to test in prospective randomised clinical trials in SSc.
References
Footnotes
-
Funding This study was funded by grants provided by Raynaud's & Scleroderma Association and the EULAR Orphan Diseases Programme (ODP) on Systemic Sclerosis.
-
Competing interests None.
-
Ethics approval This study was conducted with the approval of the Royal Free and Medical School Local Research Ethics Committee.
-
Provenance and peer review Not commissioned; externally peer reviewed.