Article Text

Abstract
Background Dermal fibroblasts from patients with systemic sclerosis (SSc) release excessive amounts of collagen resulting in tissue fibrosis. The molecular mechanisms underlying this pathological activation are incompletely understood.
Objective To investigate whether Notch signalling contributes to the uncontrolled activation of fibroblasts in SSc.
Methods Activation of the Notch pathway was assessed by immunohistochemistry or Western blot for the Notch intracellular domain and the Notch ligand Jagged-1 (Jag-1) and real-time PCR for the target gene hes-1. Differentiation of resting dermal fibroblasts into myofibroblasts was assessed by staining for α-smooth muscle actin. The synthesis of collagen was quantified by real-time PCR and Sircol assays.
Results Notch signalling was activated in lesional skin of patients with SSc. The activation persisted in cultured dermal SSc fibroblasts. Stimulation of healthy dermal fibroblasts with recombinant human Jag-1-Fc chimera resulted in an SSc-like phenotype with increased release of collagen and differentiation of resting fibroblasts into myofibroblasts. Consistent with the selective activation of the Notch pathway in dermal SSc fibroblasts, DAPT or siRNA against Notch strongly reduced the basal collagen expression in SSc fibroblasts, but not in fibroblasts from healthy volunteers.
Conclusion It was shown that Notch signalling is activated in SSc and plays an important role in fibroblast activation and collagen release. Inhibition of Notch signalling might be an effective strategy to selectively prevent the aberrant activation of SSc fibroblasts.
Statistics from Altmetric.com
Introduction
The major histopathological hallmark of systemic sclerosis (SSc) is an excessive accumulation of extracellular matrix proteins.1 2 The molecular mechanisms underlying the pathological activation of SSc fibroblasts have only partially been elucidated.
Notch signalling, first discovered in Drosophila, is essential for cell fate decision and development.3 4 Activation of the Notch pathway by binding of ligands such as Jagged-1 (Jag-1) results in cleavage of Notch receptors by the γ-secretase complex with subsequent release of the active Notch intracellular domain (NICD).3 Two families of canonical Notch ligands have been described (Jagged and Delta-like), which seem to have distinct functions.5 6 As described for T-helper cell differentiation, Jagged induces Th2 response, whereas Delta-like, mainly Dll-4, is responsible for Th1 differentiation.5 These ligands can interact with Notch receptors on both the surface of another cell (trans-interaction) and of the same cell (cis-interaction).6 In contrast to the trans-interaction, cis-interactions seem to inhibit Notch signalling. The mechanisms and effects of cis-interactions, however, are still unkown.6 In addition to the canonical ligands, several non-canonical ligands such as CCN3, MAGP-1 and MAGP-2 have been described.6 Translocation of the NICD into the nucleus activates the transcription of target genes such as the Hairy/Enhancer of Split (Hes).7 Alterations in Notch signalling are implicated in the pathogenesis of several human diseases such as T-cell acute lymphoblastic leukaemia, melanoma and Alagille syndrome.8,–,11 Clinical trials with γ-secretase inhibitors in patients with T-cell acute lymphoblastic leukaemia have already yielded promising results.12
This study aimed to investigate whether Notch signalling may promote fibroblast activation and persistently increase release of collagen in SSc.
Material and methods
Fibroblast cultures
Dermal fibroblast cultures were obtained from skin biopsy samples of patients with SSc. All patients fulfilled the criteria for SSc as suggested by LeRoy and Medsger.13 Biopsy specimens from patients with SSc (n = 10) were taken from affected skin of the forearm (online supplementary table 1). Control dermal fibroblasts (n = 10) were obtained from forearm skin biopsy specimens of healthy age- and sex-matched volunteers. Dermal fibroblasts from passages 4–8 were used for the experiments. All patients and controls signed a consent form approved by the local institutional review boards.
Stimulation of dermal fibroblasts with Jag-1 and inhibition of Notch signalling
Confluent dermal fibroblasts were cultured in Dulbecco's modified Eagle's medium containing 0.1% fetal calf serum for 24 h before the experiments. Dermal fibroblasts were stimulated with recombinant human Jag-1-Fc chimera (R&D Systems, Minneapolis, Minnesota, USA) dissolved in phosphate-buffered saline in concentrations of 0.04–5.0 µg/ml for 24 h.
To assess the effects of Notch signalling inhibition, dermal fibroblasts were incubated with the γ-secretase inhibitor N-[N-(3,5-difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT) (Sigma-Aldrich, Steinheim, Germany) at concentrations of 1.0 and 10 µmol/l for 24 h. Jag-1 was added 1 h after DAPT at a concentration of 5.0 µg/ml.
In addition to DAPT, dermal SSc fibroblasts were transfected with 1.5 µg siRNA against Notch-1 using the human dermal fibroblast nucleofector kit (Amaxa, Cologne, Germany) (supplementary table 2).14 Fibroblasts transfected with control siRNA (Ambion, Darmstadt, Germany) served as controls. Cells were harvested after 48 h.
mRNA-stability assay
To investigate whether Jag-1 stabilises collagen mRNA, an mRNA-stability assay was performed as previously described.15
Luciferase reporter assay
The −353 col 1a2/luciferase construct was kindly provided by M Trojanowska.15 HEK293T cells were transfected in serum-free medium with 0.5 µg reporter construct mixed with polyethyleneimine. A common lacZ reporter vector was used as control. Cells were stimulated with Jag-1 directly after the transfection. The luciferase activity was analysed after 24 h.
Quantitative real time PCR
Total RNA was isolated with the NucleoSpin RNA II extraction system (Macherey-Nagel, Düren, Germany) and converted into cDNA as described previously.16 In skin samples, the epidermal and dermal layers were separated for RNA isolation. The primer pairs used for real-time PCR are given in supplementary table 2. A predeveloped β-actin assay (Applied Biosystems, Darmstadt, Germany) was used to normalise for the amounts of loaded cDNA. All samples were measured in duplicate.
Quantification of soluble collagen protein
Total soluble collagen in cell culture supernatants was quantified using the Sircol collagen assay (Biocolor, Belfast, Northern Ireland) as described.17
Immunohistochemistry for Jag-1, CD3 and the NICD
Fresh frozen sections of forearm skin from patients with SSc (n = 13; supplementary table 3) and healthy individuals (n = 10) were used for immunohistochemical analyses (see online supplementary methods).
Immunofluorescence for α-smooth muscle actin (αSMA) and the NICD
Fibroblasts stimulated with Jag-1-Fc (5 µg/ml) were incubated with monoclonal anti-αSMA antibodies (clone 1A4; Sigma-Aldrich) and polyclonal rabbit-anti-human NICD (Abcam, Cambridge, UK). TRITC-conjugated goat anti-mouse antibodies and FITC-conjugated goat anti-rabbit antibodies (Invitrogen, Karlsruhe, Germany) served as secondary antibodies. Counterstaining was performed with 4',6-diamidino-2-phenylindole (DAPI; Santa Cruz Biotechnologies, Santa Cruz, California, USA) for 10 min at room temperature.
Western blot
Whole-cell lysates were prepared as previously described.18 Polyvinylidene fluoride membranes were incubated with polyclonal rabbit-anti Notch-1 (Abcam) or polyclonal rabbit-anti activated Notch-1 (Abcam). Horseradish peroxidase-conjugated polyclonal goat anti-rabbit antibodies (DAKO, Hamburg, Germany) were used as secondary antibodies. Equal loading of proteins was confirmed by visualisation of α-tubulin (Invitrogen).
Statistics
Data are expressed as mean ± SEM. The Wilcoxon signed rank test for related samples and the Mann–Whitney U test for non-related samples were used for statistical analyses. A p value <0.05 was considered statistically significant.
Results
Notch signalling is activated in lesional skin of patients with SSc
We first analysed the expression of the four Notch receptors in dermal fibroblasts. The levels of Notch-1 mRNA were significantly increased in SSc fibroblasts as compared with controls (online supplementary figure 1). Notch-2 was detectable, but in contrast to Notch-1, its levels did not differ between SSc and controls. Notch-3 and Notch-4 were not detectable by real-time PCR in dermal fibroblasts as these receptors are predominantly expressed in vascular smooth muscle cells (VSMCs) and vascular endothelial cells.19
To investigate whether Notch signalling is activated in lesional skin of patients with SSc, we stained for the NICD, which is only released upon ligand binding to Notch and therefore is a marker for activation of the Notch pathway.3 The NICD was almost undetectable in healthy individuals (figure 1A). In contrast, we observed an intensive staining in all patients with SSc, particularly in fibroblasts of the papillary dermis (figure 1A). Staining for the NICD, however, was not restricted to fibroblasts and was also prominent in keratinocytes, endothelial cells, pericytes, VSMCs and T cells as described before (figure 1A).20 Consistent with increased Notch signalling, the mRNA levels of the Notch target gene hes-1 were elevated by 392 ± 33% in skin biopsy samples of patients with SSc as compared with similar samples from healthy individuals (p < 0.05) (figure 1B). To investigate the mechanisms that activate Notch signalling in SSc, we analysed the expression of Notch ligands and observed an upregulation of Jag-1 mRNA levels by 608 ± 86% in SSc skin compared with control skin samples (p < 0.05) (figure 1C). Expression of Jag-1 protein was detected in epidermis and dermis of all patients with SSc (figure 1D). A prominent expression of Jag-1 was seen in inflammatory infiltrates in fibrotic skin. Double staining for Jag-1 and CD3 showed elevated Jag-1 expression in T cells (figure 1E). In contrast to patients with SSc, no significant expression of Jag-1 was detected in healthy individuals.

Notch signalling is activated in lesional skin of patients with systemic sclerosis (SSc). (A) An intensive staining for the Notch intracellular domain (NICD) was observed in fibroblasts in skin sections of patients with SSc, whereas the NICD was almost undetectable in skin sections of healthy individuals. (B) The Notch target gene hes-1 is overexpressed in lesional skin of patients with SSc. (C) Increased mRNA levels of the Notch ligand Jag-1 in skin of patients with SSc. (D) Jag-1 is overexpressed in lesional skin of patients with SSc. (E) Jag-1 expression is elevated in T cells of inflammatory infiltrates of fibrotic skin. n = 13 for SSc samples and n = 10 for healthy skin samples. Representative skin sections are shown at 200-fold (A and D) or 400-fold (E) magnification, fibroblasts are shown at 1000-fold magnification. Asterisks indicate statistical significant differences compared with skin samples from healthy controls.
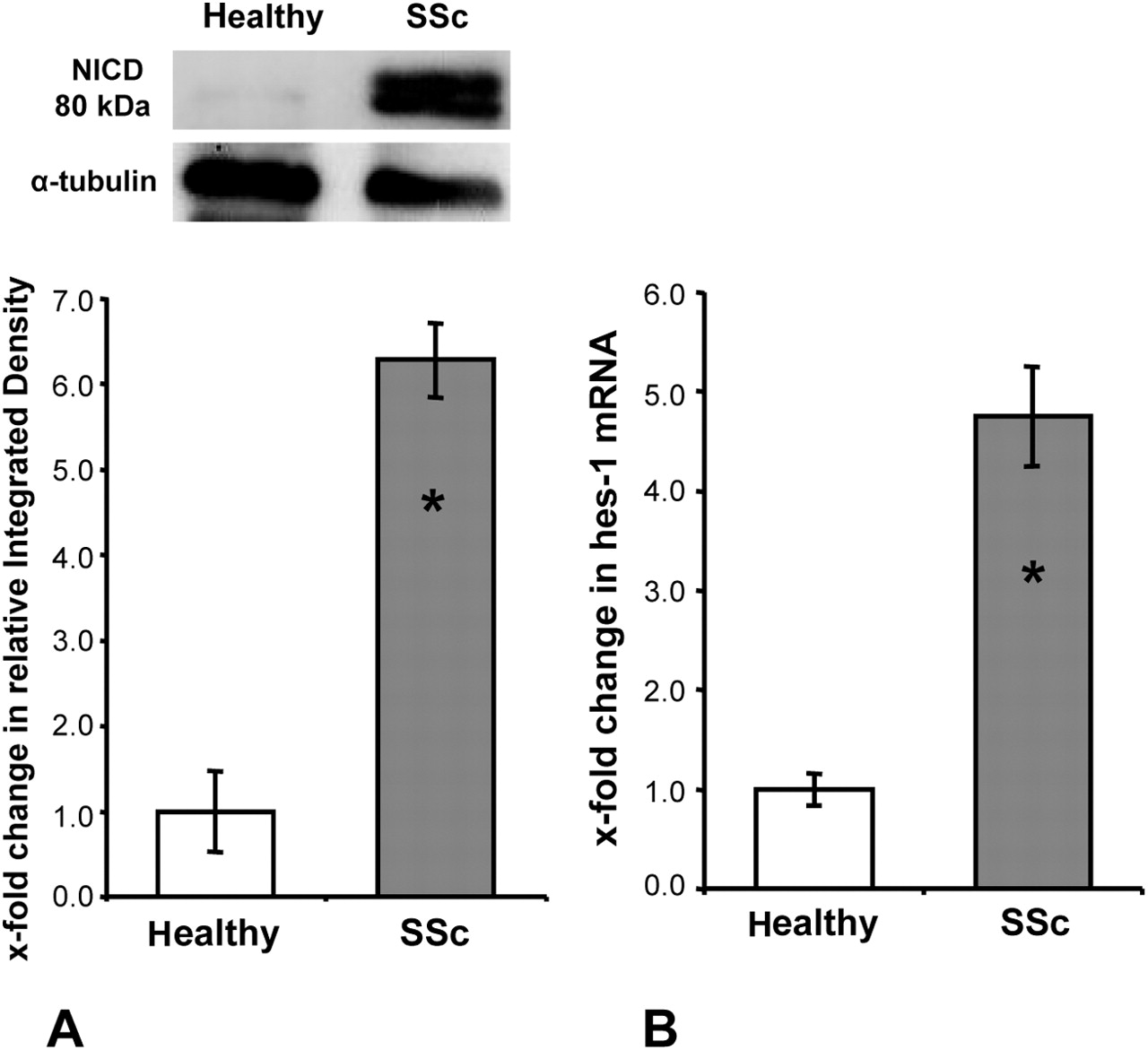
Persistent activation of the Notch pathway in cultured SSc fibroblasts
Of particular interest, the increased activation of the Notch pathway in dermal SSc fibroblasts persisted in vitro. A significant accumulation of the NICD occurred in SSc fibroblasts, whereas the NICD was barely detectable in fibroblasts from healthy individuals (figure 2A). Consistently, the mRNA levels of hes-1 were upregulated in cultured SSc fibroblasts in passages 4–8 with increases of 375 ± 50% compared with fibroblasts from healthy individuals (p < 0.05) (figure 2B).

Persistent activation of the Notch pathway in systemic sclerosis (SSc) fibroblasts after long-term culture. (A) Elevated levels of the Notch intracellular domain (NICD) in SSc fibroblasts as determined by Western blot (n = 3 each). (B) Increased mRNA levels of the target gene hes-1 (n = 8 for healthy fibroblasts, n = 6 for SSc fibroblasts). Asterisks indicate statistical significant differences compared with healthy dermal fibroblasts.
Jag-1 stimulates the release of extracellular matrix
To investigate whether the overexpression of Jag-1 and the increased activation of the Notch pathway contribute to the aberrant production of extracellular matrix, healthy dermal fibroblasts were stimulated with recombinant Jag-1.21 22 Activation of the Notch pathway upon stimulation with Jag-1-Fc was confirmed by a dose-dependent increase in hes-1 mRNA levels (figure 3A). Jag-1, reaching a maximum at concentrations of 5 µg/ml, upregulated dose-dependently the expression of extracellular matrix components (figure 3B–D). Stimulation with Jag-1-Fc increased the mRNA levels of col 1a1 and col 1a2 to 294 ± 37% and 217 ± 28%, respectively (p < 0.05) (figure 3B,C). Consistent with the results on the mRNA level, collagen protein in cell culture supernatants increased to 259 ± 32% upon stimulation with Jag-1-Fc as assessed by the Sircol assay (p < 0.05) (figure 3D).

Jag-1 stimulates the synthesis of collagen in fibroblasts. Stimulation of healthy dermal fibroblasts with Jag-1 at concentrations from 0.04 to 5 µg/ml increased the mRNA levels of hes-1 (A), col 1a1 (B) and col 1a2 (C) in a dose-dependent manner. Consistently, Jag-1 stimulated the release of collagen protein (D) (n = 8 each). Asterisks indicate statistical significant differences compared with unstimulated fibroblasts.
To discriminate between increased gene transcription and altered mRNA stability in response to stimulation with Jag-1-Fc, we performed mRNA stability and reporter assays. The relative changes in mRNA levels of extracellular matrix proteins did not differ between Jag-1-Fc-stimulated fibroblasts and controls over time upon incubation with actinomycin D, suggesting that Jag-1-Fc does not increase mRNA stability (supplementary figure 2A). In contrast, Jag-1-Fc stimulated collagen transcription by 173 ± 12% as analysed by a luciferase expression vector under the control of the col 1a2 promoter (p < 0.05) (supplementary figure 2B).
Jag-1 induces differentiation of resting fibroblasts into myofibroblasts
Myofibroblasts play an important role in the pathogenesis of fibrotic diseases. They are identified by the expression of contractile proteins such as αSMA. Upon stimulation with Jag-1-Fc, the mRNA levels of αSMA increased dose-dependently by 239 ± 37% (p < 0.05) in normal dermal fibroblasts (figure 4A). Intensive staining for αSMA protein and for the NICD was observed in Jag-1-Fc-stimulated healthy fibroblasts as analysed by immunofluorescence (figure 4B). The percentage of double-positive cells increased from 19% in unstimulated control cells to 82% in Jag-1-Fc-stimulated fibroblasts, whereas only 3% of the cells were positive for the NICD but negative for αSMA, and 6% of the cells were negative for the NICD but positive for αSMA. Together, these data suggested that Jag-1-Fc stimulates differentiation of resting fibroblasts into myofibroblasts.

Jag-1 induces differentiation of resting fibroblasts into myofibroblasts. (A) Jag-1 increased the mRNA levels of α-smooth muscle actin (αSMA) in a dose-dependent manner (n = 8). Asterisks indicate statistical significant differences compared with unstimulated fibroblasts. (B) Jag-1 induced the release of the Notch intracellular domain (NICD) and the expression of αSMA protein in resting fibroblasts (n = 5). Representative examples are shown at 200-fold magnification. DAPI, 4',6-diamidino-2-phenylindole.
DAPT effectively reduces the stimulatory effects of Jag-1
The activity of γ-secretase is essential for the activation of Notch signalling. To evaluate the effects of the γ-secretase inhibitor DAPT on the release of extracellular matrix, we incubated Jag-1-stimulated healthy dermal fibroblasts with DAPT at pharmacologically relevant concentrations.23 24 DAPT inhibited the mRNA expression of col 1a1 in healthy dermal fibroblasts stimulated with Jag-1-Fc dose-dependently by up to 99 ± 23% (p < 0.05) (supplementary figure 3A). Similar effects were obtained for col 1a2 (p < 0.05) (supplementary figure 3B). Consistent with reduced mRNA levels, the release of collagen protein was reduced by 94 ± 34% upon pretreatment with DAPT (p < 0.05) (supplementary figure 3C).
Inhibition of Notch signalling reduces the basal collagen synthesis specifically in SSc fibroblasts
We demonstrated a persistent activation of the Notch pathway in cultured fibroblasts from patients with SSc. To assess whether inhibition of the Notch pathway can inhibit the basal release of collagen, SSc fibroblasts were incubated with DAPT in the absence of Jag-1-Fc. Treatment of unstimulated SSc fibroblasts with DAPT reduced the mRNA levels of col 1a1 and col 1a2 by up to 49 ± 9% and 41 ± 6%, respectively (p < 0.05 for both) (figure 5A,B). The collagen content in the supernatants was reduced by up to 59 ± 11% upon incubation with DAPT (p < 0.05) (figure 5C). Consistent with the persistent activation of the Notch pathway in SSc fibroblasts, no effects of DAPT on collagen expression were detected in cultured fibroblasts from healthy individuals in the absence of exogenous Jag-1 (figure 5A–C).

The γ-secretase inhibitor DAPT reduces the basal synthesis of extracellular matrix only in systemic sclerosis (SSc) fibroblasts. DAPT reduced the basal mRNA levels of col 1a1 (A) and col 1a2 (B) and decreased the release of collagen protein (C) in SSc fibroblasts (n = 7). In contrast, no effect of DAPT on the basal collagen expression was seen in healthy fibroblasts in the absence of exogenous Jag-1 (A–C) (n = 6). Asterisks indicate statistical significant differences compared with untreated fibroblasts.
To exclude off-target effects, we confirmed the antifibrotic effects of DAPT signalling by transfection of SSc fibroblasts with siRNA against Notch-1, which resulted in significantly decreased mRNA and protein levels of Notch-1 (p < 0.05) (figure 6A). Knockdown of Notch-1 efficiently reduced mRNA levels of col 1a1 and col 1a2 by 58 ± 11% and 48 ± 9% compared with mock transfected SSc fibroblasts (figure 6B,C). Consistently, the release of collagen protein was reduced by 42 ± 8% (p < 0.05) upon silencing of Notch-1 (figure 6D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Knockdown of Notch-1 by siRNA decreases collagen expression in systemic sclerosis (SSc) fibroblasts. Nucleofection of SSc fibroblasts with 1.5 µg of siRNA against Notch-1 efficiently reduced Notch-1 mRNA and protein (A). Knockdown of Notch-1 decreased the mRNA levels of col 1a1 (B) and col 1a2 (C) and reduced the release of collagen (D) (n = 6 each). Asterisks indicate statistical significant differences compared with mock transfected fibroblasts.
Discussion
We have shown in this study that Notch signalling is activated in SSc with a prominent expression of the ligand Jag-1 in infiltrating T cells and active Notch signalling with accumulation of the NICD in fibroblasts. We further showed that the activation of Notch signalling has major implications on the activation of fibroblasts. The expression patterns of Jag-1 and the NICD suggest that infiltrating T cells expressing Jag-1 might activate Notch signalling in resident fibroblasts. Jag-1 is prominently expressed in T cells in fibrotic skin. Direct cell–cell contact between T cells expressing the ligand Jag-1 and fibroblasts expressing high levels of Notch-1 may be the major mechanism for the activation of Notch signalling in SSc fibroblasts, with accumulation of the NICD and increased transcription of target genes such as hes-1 in fibroblasts. Considering the potent stimulatory effects of Jag-1-Fc on cultured fibroblasts, T-cell-mediated activation of Notch signalling in fibroblasts might be an important mechanism for the aberrant activation of fibroblasts in SSc. However, further studies are needed to confirm this hypothesis and to further elucidate the molecular mechanism of this activation. Consistent with our findings of an increased activation of Notch signalling in SSc, Kavian et al recently demonstrated an activation of Notch signalling in SSc as well as in mouse models of dermal fibrosis.25
We have shown that activation of Notch signalling induces a profibrotic phenotype in resting dermal fibroblasts. Jag-1-Fc stimulates the release of collagen protein and induces differentiation of resting dermal fibroblasts into myofibroblasts. Consistent with the increased expression of contractile proteins seen in our study, Notch may also regulate migration of fibroblasts.23 Thus, pathological activation of Notch signalling stimulates resting dermal fibroblasts to express contractile proteins, differentiate into myofibroblasts and release excessive amounts of extracellular matrix proteins.
Although evidence for the important role of Notch signalling is growing, it remains unclear how Notch signalling induces myofibroblast differentiation and stimulates the release of collagen in fibrotic diseases on a molecular level. In this context, it is still not clear whether hes-1 induction is essential for these stimulatory effects. Notch signalling may also be induced under hypoxic conditions or by transforming growth factor β.26,–,28 Consistently, we have preliminary evidence that Notch-1 and hes-1 are increased in dermal fibroblasts upon stimulation with transforming growth factor β or under hypoxia. In addition, NICD and pSmad3 seem to be co-expressed in skin of patients with SSc. Moreover, recent work has indicated that local production of reactive oxygen species might induce ADAM-17, a protease involved in the activation of Notch signalling.25
Finally, we demonstrated that inhibition of the Notch pathway by either pharmacological blockade of the γ-secretase complex or siRNA against Notch-1 exerted potent antifibrotic effects selectively in dermal SSc fibroblasts, whereas the basal collagen synthesis in healthy dermal fibroblasts was not affected by inhibition of Notch signalling. Thus, targeting Notch signalling might have the potential to selectively reduce the activation of dermal SSc fibroblasts without affecting the collagen synthesis in resting normal dermal fibroblasts. In addition, pharmacological blockade of the γ-secretase complex might decrease the proliferation of SSc fibroblasts. Our in vitro findings are further extended by recent work by Kavian et al.25 Treatment with the γ-secretase inhibitor DAPT prevented development of experimental dermal and pulmonary fibrosis. In mice, DAPT significantly ameliorated the development of dermal and pulmonary fibrosis, decreased proliferation of fibroblasts, reduced the production of H2O2 and abrogated the production of anti-DNA topoisomerase-1 antibodies.25 These findings might have translational implications as different inhibitors of the γ-secretase complex are in clinical trials for different types of cancer (http://www.clinicaltrials.gov). However, given the activation of Notch signalling in endothelial cells in our study and the role of Notch signalling in vascular homoeostasis (reviewed by Gridley 29), inhibition of Notch signalling might be hindered by antiangiogenic effects in patients with SSc who have extensive small-vessel vasculopathy with capillary rarifications. By contrast, inhibitors of the γ-secretase complex might be beneficial in patients with proliferative vasculopathies such as pulmonary arterial hypertension (PAH), because Notch-3 is activated in VSMCs from PAH tissue and contributes to the increased proliferation of VSMCs.30
In summary, Notch signalling is activated in lesional skin of patients with SSc. Considering its potent stimulatory effects on dermal fibroblasts, Notch signalling might contribute to the pathological activation of SSc fibroblasts.
Acknowledgments
The authors would like to thank Maria Halter for excellent technical support.
References
Supplementary materials
Web Only Data
Files in this Data Supplement:
Footnotes
-
Funding The study was supported by the Deutsche Forschungsgesellschaft (DI 1537/2-1), grants A20 and A40 of the Interdisciplinary Center of Clinical Research (IZKF) in Erlangen, by the Career Support Award of Medicine of the Ernst Jung Foundation and by support of CMH Research Projects No 00000023728.
-
Competing interests None.
-
Ethics approval This study was conducted with the approval of the ethics committee of the University of Erlangen.
-
Provenance and peer review Not commissioned; externally peer reviewed.