Article Text

Abstract
Objectives Erdheim-Chester disease (ECD) is a rare form of histiocytosis characterised by uncontrolled chronic inflammation. The oncogenic BRAFV600E mutation has been reported in biopsies in 19 out of 37 patients with ECD from the largest published cohort, but never found in the patients’ peripheral blood. Also, the role of the mutation in the pathogenesis of the disease has not been elucidated yet. BRAFV600E has been associated with oncogene-induced senescence (OIS), a protective mechanism against oncogenic events, characterised by the induction of proinflammatory pathways.
Methods We verified the BRAF status in biopsies and peripheral blood from 18 patients with ECD from our cohort and matched controls by means of immunohistochemistry and of an ultrasensitive assay, based on the combination of a locked nucleic acid PCR and pyrosequencing. Droplet digital PCR was used to confirm the findings. We also evaluated the presence of senescence markers in ECD histiocytes.
Results BRAFV600E mutation was present in all the biopsy and peripheral blood samples from patients with ECD and in none of the controls. ECD histiocytes and a fraction of circulating monocytes from patients with ECD showed signs of a constitutive activation of the MAPK pathway. Moreover, BRAF-mutated histiocytes expressed markers of OIS.
Conclusions The oncogenic BRAFV600E mutation is present in biopsies and in the peripheral blood from all patients with ECD who were evaluated and is associated with OIS. These findings have significant implications for the pathogenesis, diagnosis and treatment of ECD.
- Inflammation
- Cytokines
- Gene Polymorphism
Statistics from Altmetric.com
Introduction
Erdheim-Chester disease (ECD) is an extremely rare form of non-Langerhans’ cell histiocytosis (LCH), characterised by infiltration of tissues by ‘foamy’ lipid-laden macrophages.1 To date, about 500 cases have been described worldwide.2 Since histiocytes can infiltrate and accumulate in virtually every tissue and system, the disease leads to protean manifestations.1 ,3 ,4 The sites of organ involvement have a major impact on the clinical course and prognosis of the disease, which can range from asymptomatic/oligosymptomatic to multisystemic forms associated with a poor prognosis, especially when the central nervous and the cardiovascular system are affected.5
The pathogenesis of ECD is not clearly understood. In the past, some authors have suggested that ECD might be a clonal disease, but this observation has not been univocally confirmed.6–8 Subsequently, our group and others have demonstrated the occurrence in patients with ECD of a local and systemic proinflammatory cytokine-chemokine network, which may be responsible for the recruitment and activation of histiocytes into ECD lesions.9–12 These observations led to the hypothesis that cytokine blockade might be a possible therapeutic strategy for ECD, and clinical observations on small cohorts of patients have demonstrated that interleukin 1 and tumour necrosis factor α blockade can be beneficial.13 ,14
More recently, it has been shown that infiltrating histiocytes from a significant proportion of patients with ECD bear a mutation in the proto-oncogene BRAF.15 ,16 In particular, the analysis of the largest ECD cohort indicated, by means of direct pyrosequencing techniques, that 19 out of 37 patients harboured a mutation that determines the amino acid substitution of glutamic acid for valine at position 600 of the BRAF protein (V600E).17 Intriguingly, the same mutation was recently found in up to 57% of studied specimens of LCH.18–21 The hypothesis of a crucial relevance of BRAFV600E mutation in ECD pathogenesis is also supported by the dramatic response to BRAFV600E pharmacological inhibition observed in three patients with ECD who harboured the mutation.22 BRAF is a serine/threonine protein kinase that contributes to the RAS-RAF-MEK-ERK protein kinase pathway. This signalling pathway regulates cell proliferation and survival and is typically activated by extracellular growth factors binding to receptor tyrosine kinases.23 BRAFV600E is a mutational hot spot in melanomas, papillary thyroid cancers and hairy-cell leukaemia, and its oncogenic activity in these cancers has been extensively documented.24–26 Of interest, BRAFV600E mutation has been also associated with oncogene-induced senescence (OIS), a major protective response against oncogenic events aimed to restrain tumour growth and characterised by stable proliferative arrest and by an increased expression of tumour suppressor proteins, such as p16Ink4a and p21.27–29 Senescent cells have been shown to implement a complex proinflammatory response, known as senescence-associated secretory phenotype (SASP).30 Intriguingly, this inflammatory phenotype shares many similarities with the one described in ECD.9–11 ,14
With this study, we aimed to evaluate by means of ultrasensitive molecular biology techniques the presence of the BRAFV600E mutation in all the histological specimens from our cohort of patients, and to investigate whether the same mutation is detectable in the peripheral blood mononuclear cells (PBMCs) of patients with ECD. We finally wanted to evaluate if a role of OIS and SASP in ECD pathogenesis can be envisaged.
Methods
Patients and controls
We evaluated all the available stored tissue samples from 18 patients with ECD followed at San Raffaele Hospital, Milan. ECD diagnosis relied on clinical and imaging evaluation by dedicated physicians and was always histologically confirmed by at least two independent pathologists. Specimens from five histological samples from non-ECD xanthogranulomatous lesions (two patients with retro-orbital inflammatory pseudotumour, one with inflammatory pseudotumour of the lung, one with sclerosing mesenteritis and one with idiopathic retroperitoneal fibrosis) served as negative controls. We also obtained peripheral blood samples from 16 of those 18 patients and from 16 age-matched and sex-matched healthy donors. A written informed consent was obtained from all patients and controls.
PMBC collection and monocyte-derived macrophages (MDM) cultures
PBMCs were isolated from EDTA-anticoagulated venous blood obtained from patients and matched controls through Ficoll-Paque gradient centrifugation. MDM cultures were generated from freshly isolated monocytes, obtained from PBMC upon adhesion on glass coverslips (Prestige, Italy). After 1 h of incubation, non-adherent cells were washed away and the adherent fraction was cultured with RPMI (Life Technologies, Paisley UK) in addition with 20% human AB serum. After 3 days, cultures were fixed in 2% paraformaldehyde, permeabilised with 0.1% Triton X-100 and prepared for immunohistochemical studies.
BRAF sequencing
Genomic DNA was extracted from formalin-fixed paraffinated samples and frozen purified PBMC using the Qiamp DNA Mini Kit (Qiagen Gmbh, Hilden, Germany) according to the manufacturer's protocol. BRAF codon 600 mutations were investigated. Genomic DNA was amplified for BRAF using anti-EGFR monoclonal antibody (MoAb) response (BRAF status) kit (Diatech Pharmacogenetics, Jesi, Italy) following the manufacturer's instructions. Pyrosequencing analysis of PCR products was conducted according to standard procedures using PyroMark Gold Q96 (Qiagen) reagents with PyroMark Q96ID. Pyrogram outputs were analysed with PyroMark Q96ID Software (Qiagen) using the allele quantification mode. All analyses were performed at least twice on two independent PCRs.
Minor-allele amplification assay
To increase the sensitivity of the standard DNA pyrosequencing, we set up a locked nucleic acid (LNA)-PCR/pyrosequencing assay. We performed BRAFV600E LNA-PCR as previously described.31 The amplicon products were used as PCR template for pyrosequencing, following the same steps as above. The full details of this assay are illustrated in the online supplementary materials.
Droplet Digital PCR
BRAFV600E mutation was analysed by droplet digital PCR (ddPCR) in the histological and PBMC samples from the first eight patients with ECD, in five PMBC samples from healthy controls, and one BRAFV600E-positive melanoma tissue sample. ddPCR was performed on a QX100 ddPCR system (Bio-Rad, Hercules, CA) using prevalidated BRAFV600E PrimePCR ddPCR Mutation Assays (Bio-Rad), based on forward and reverse gene specific primers and fluorescent hydrolysis probes specific for either the mutant (V600E-FAM) or the wild type (WT V600-HEX) sequence at codon 600. Briefly, DNA was digested with Hind-III and a total of 200 ng were used to set up eight replicate ddPCR reactions for each sample; these were emulsified in a QX100 droplet generator, transferred to 96 well plates and subjected to thermal cycling on a T100 instrument, according to the manufacturer's instructions (Bio-Rad). After amplification, plates were read and individual sample droplets analysed on a QX100 droplet reader. The per cent fraction of mutant relative to wild type copies was determined using the QuantaSoft V.1.2.10 software, applying a correction based on the Poisson distribution to the counted number of droplets positive for either mutant or wild type BRAF DNA.
Immunohistochemical studies
Formalin-fixed biopsies and MDM cultures were stained with the VE1 monoclonal antibody that has been shown to be specific for BRAFV600E mutated protein.15 ,32
Staining for CD68 (Dako, Glostrup, Denmark), p16Ink4a (JC8 clone, Santa Cruz Biotechnology, Dallas, TX), Phospho-p44/42 MAPK Erk1/2 Thr202/Tyr204 (Cell Signalling Technology, Danvers, Massachusetts, USA) was performed incubating primary antibodies 1 h at room temperature, followed by detection with horseradish peroxidase-conjugate-polymer-conjugate-polymer (Thermo Scientific, Fremont, California, USA) and developed with DAB chromogen. Tissue sections were counterstained with haematoxylin.
Detection of phospho-ERK in peripheral monocytes
Assessment of phosphorylated extracellular signal-regulated kinase (phospho-ERK) expression in PBMC was performed by flow cytometry. Freshly drawn peripheral blood samples obtained from six patients with ECD and six controls were immediately placed on ice to stop intracellular signalling and fixed using the Lyse/Fix Buffer-BDPhosflow (Becton Dickinson, San Josè, California, USA). Cells were then permeabilised with 90% methanol for 30 min on ice, stained with Alexa Fluor 488-conjugated antiphospho-ERK-1/2 (pT202/pY204) and phycoerythrin-conjugated anti-CD14 monoclonal antibodies (Becton Dickinson), and analysed using FC500 flow cytometer (Beckman Coulter, Brea, California, USA).
Statistical analysis
Statistical analyses were performed using GraphPad Prism V.5.0 for Macintosh (GraphPad Software, San Diego, CA). We used as basic descriptors the mean, median, SD and SEM. Statistical comparisons between patients and controls and between our and literature data were carried out with the Mann-Whitney U test and with Fisher’s exact test. A two-tailed p value <0.05 was considered statistically significant.
Results
Patient population
We were able to analyse 18 biopsy tissue samples from 18 patients with ECD. Twelve out of 18 patients with ECD from the study cohort were men, and 6 were women. The median age at the diagnosis was 57 years (range, 32–79 years). As a negative control, we used samples from five patients with xanthogranulomatous diseases other than ECD. We were also able to obtain peripheral blood samples from 16 out of the18 patients with ECD.
BRAFV600E mutation in histological samples
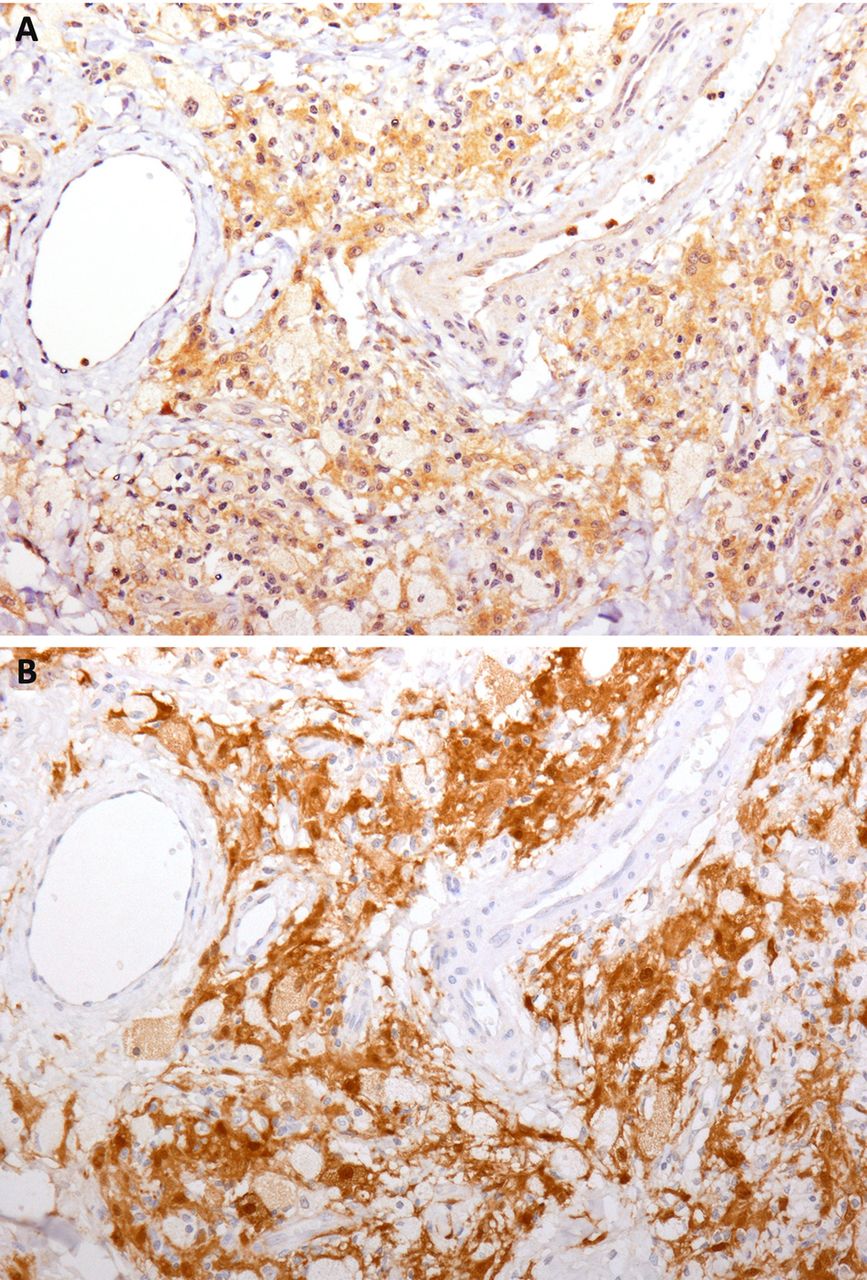
We first evaluated the presence of mutated BRAF in histological samples obtained from patients with ECD by means of direct pyrosequencing. We found BRAFV600E mutation in 12 of 18 studied biopsies (67%) and in none of 5 control tissues (table 1), in accordance with what was already described.15 ,17 Conversely, when we analysed ECD biopsies by immunohistochemistry using the specific anti-BRAFV600E monoclonal antibody VE1, we found that all the 18 studied biopsies stained positive for BRAFV600E. BRAFV600E expression was restricted to histiocytes, whereas other cells (such as lymphocytes, endothelial cells and fibroblasts) were negative. Also, a significant number of BRAF-negative histiocytes were present in ECD lesions (figure 1). Given the conflicting results obtained with the two techniques, we aimed to increase the sensitivity of BRAFV600E molecular detection in ECD lesions. Indeed, the great variability among samples in the proportion of mutated histiocytes (from 20% to 50%) over total infiltrating cells might have affected BRAFV600E detection by direct pyrosequencing. We therefore performed a BRAFV600E-specific LNA-PCR prior to performing pyrosequencing. This approach, which has been previously exploited to detect circulating BRAF-mutated melanoma cells,33 allows for the identification of 1 mutated DNA copy out of 20 000 wild type copies. By means of this combined assay, we were able to detect BRAFV600E mutation in all the available ECD specimens (18 out of 18) and in none of the 5 control tissues (tables 1 and 2).
Clinical characteristics and BRAF status of 18 patients with Erdheim-Chester disease (ECD)
BRAFV600E mutation in patients with Erdheim-Chester disease (ECD) and controls as evaluated by direct pyrosequencing alone and by a locked nucleic acid PCR (LNA-PCR)/pyrosequencing combined assay

BRAFV600E is expressed by a variable fraction of Erdheim-Chester disease (ECD) histiocytes. Immunohistochemical studies with the BRAFV600E-specific antibody VE1 on two representative cases show cytoplasmic staining of a variable fraction of histiocytes. Some histiocytes and all the other inflammatory cells are negative for the mutated protein. The two panels depict the variability of the proportion of the mutated cells over the entire cellular population (VE1 immunostaining with diaminobenzidine and haematoxylin counterstain, original magnification, ×200).
BRAFV600E mutation in PBMC samples
We then evaluated the presence of BRAFV600E in PBMC obtained from 16 patients with ECD and 16 age-matched and sex-matched controls. We were unable to detect BRAFV600E in any of the studied samples by direct pyrosequencing. However, when coupling pyrosequencing to LNA-PCR, we detected the BRAFV600E mutation in DNA from all PBMC samples from patients with ECD and in none of those obtained from healthy controls (tables 1 and 2).
Droplet digital PCR detection of BRAFV600E
To further confirm our findings, the BRAF gene mutation status at codon 600 was further analysed using ddPCR in PBMC and histological samples from the first eight patients with ECD, and in five non-mutated controls. BRAFV600E was detected in all the tested samples from patients with ECD, with a median fractional abundance of mutated DNA relative to wild type of 0.06% (range 0.04–0.32) in PBMC, and of 7.82% (range 3.04–14.10) in biopsies. BRAFV600E was not detected in any of the five PBMC samples obtained from healthy controls. ddPCR detected BRAFV600E in the control melanoma sample, with a fractional abundance of 49.3% (see online supplementary figure 1).
A small fraction of MDM from patients with ECD are BRAFV600E-positive
To further substantiate the presence of circulating cells bearing the BRAFV600E mutation, we propagated cultures of MDM from six patients with ECD and six healthy controls. Upon staining with the BRAFV600E-specific monoclonal antibody VE1, a small fraction (ranging from 1% to 3%) of positive adhering macrophages obtained from patients with ECD were identified (figure 2A), whereas no cells were positive in paired cultures obtained from healthy donors.
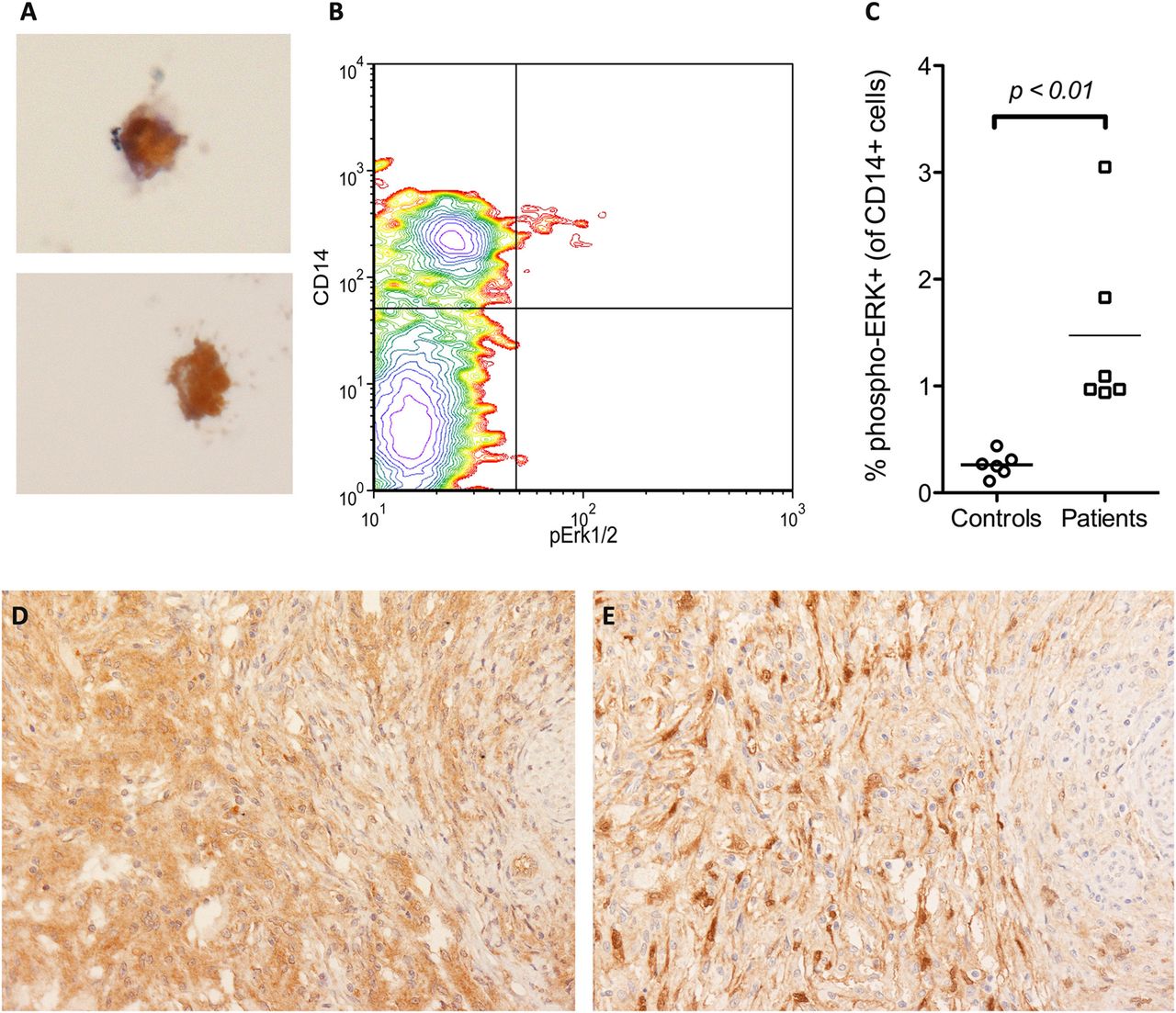
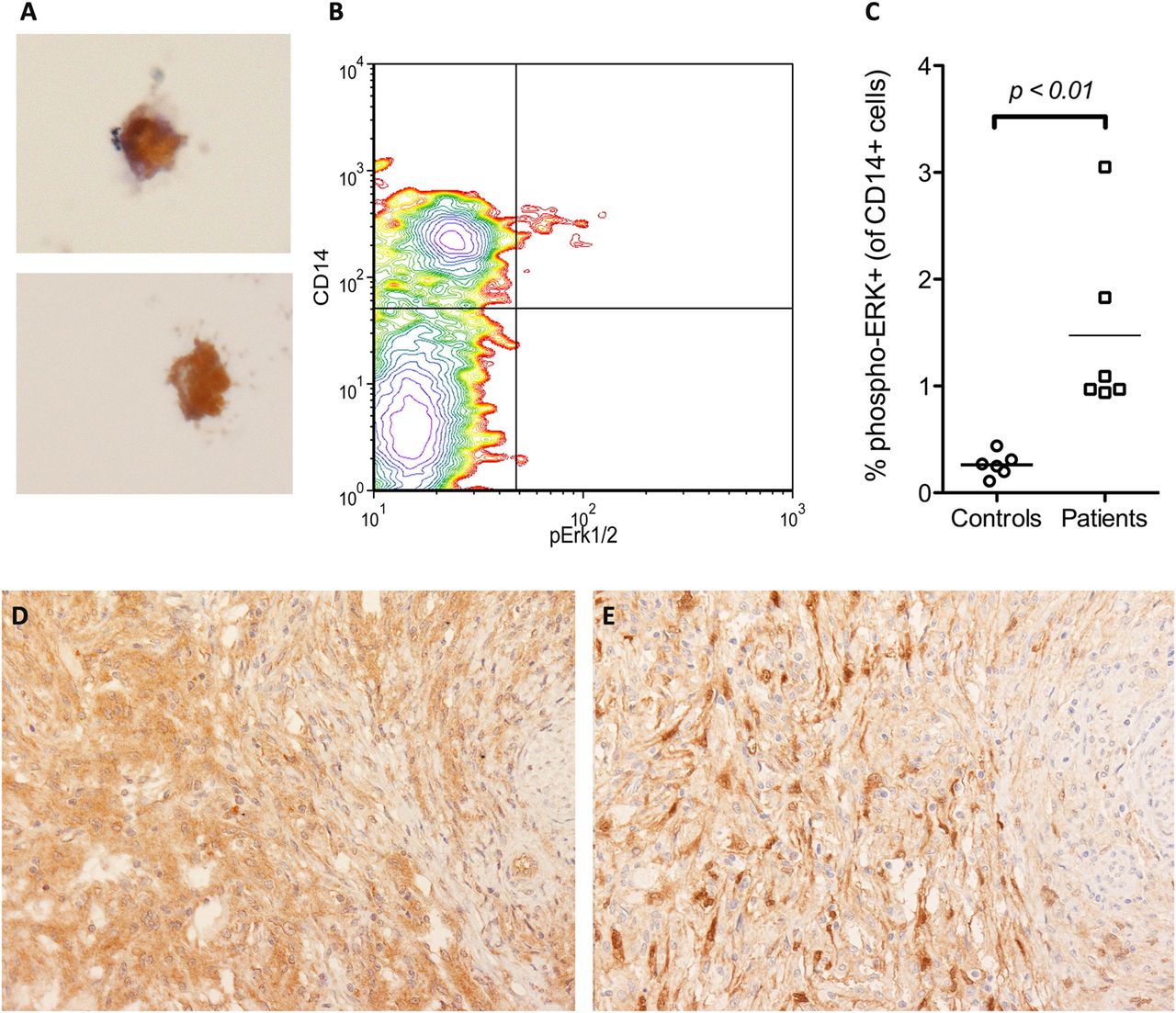
Peripheral blood monocytes and macrophages from patients with Erdheim-Chester disease (ECD) express BRAFV600E-mutated protein and have an activated mitogen-activated protein kinase pathway. A small fraction (1–3%) of peripheral blood monocyte-derived macrophages cultured ex vivo from patients with ECD stained positive for the BRAFV600E-specific VE1 antibody at immunohistochemistry (panel A, VE1 immunostaining with diaminobenzidine and haematoxylin counterstain, original magnification ×400). Flow cytometry analysis of a representative peripheral blood sample obtained from a patient with ECD, stained positive for the monocytic marker CD14 and for phosphorylated extracellular signal-regulated kinase (phospho-ERK), the major downstream target of BRAF activation; the phospho-ERK-positive population falls only inside the CD14-positive subset, and in this patient accounts for 2.7% of total CD14-positive cells (panel B). In the six evaluated patients with ECD, the percentages of phospho-ERK-positive monocytes ranged from 0.95% to 3.05% of total CD14-positive circulating cells and were significantly higher than those observed in six matched controls (Panel C). BRAFV600E-mutated histiocytes from ECD lesions (panel D, VE1 immunostaining, ×200) stained also for phospho-ERK in consecutive sections (panel E, phospho-ERK immunostaining, ×200), thus indicating that BRAFV600E-positive histiocytes have an activated mitogen-activated protein kinase pathway.
BRAF downstream pathway is activated in lesion histiocytes and in circulating monocytes from patients with ECD
Since BRAFV600E causes constitutive and direct activation of the downstream kinases MEK and ERK, we evaluated whether ECD histiocytes stained positive for the activated form of ERK. As expected, BRAFV600E-positive cells were also positive for phospho-ERK in all the 18 ECD biopsy samples (figure 2D,E). We then evaluated the ERK activation status in PBMC from patients with ECD by means of flow cytometry. We found a small fraction of phospho-ERK-positive circulating cells in all the studied patients (six out of six) and in none of the controls (zero out of six). This cellular population also stained positive for CD14, but not for lymphocyte markers, and represented 0.95–3.05% of total CD14-positive cells (figure 2B,C), consistent with the frequency of BRAF-mutated MDM previously demonstrated by immunohistochemistry.
Identification of senescence markers in ECD histiocytes
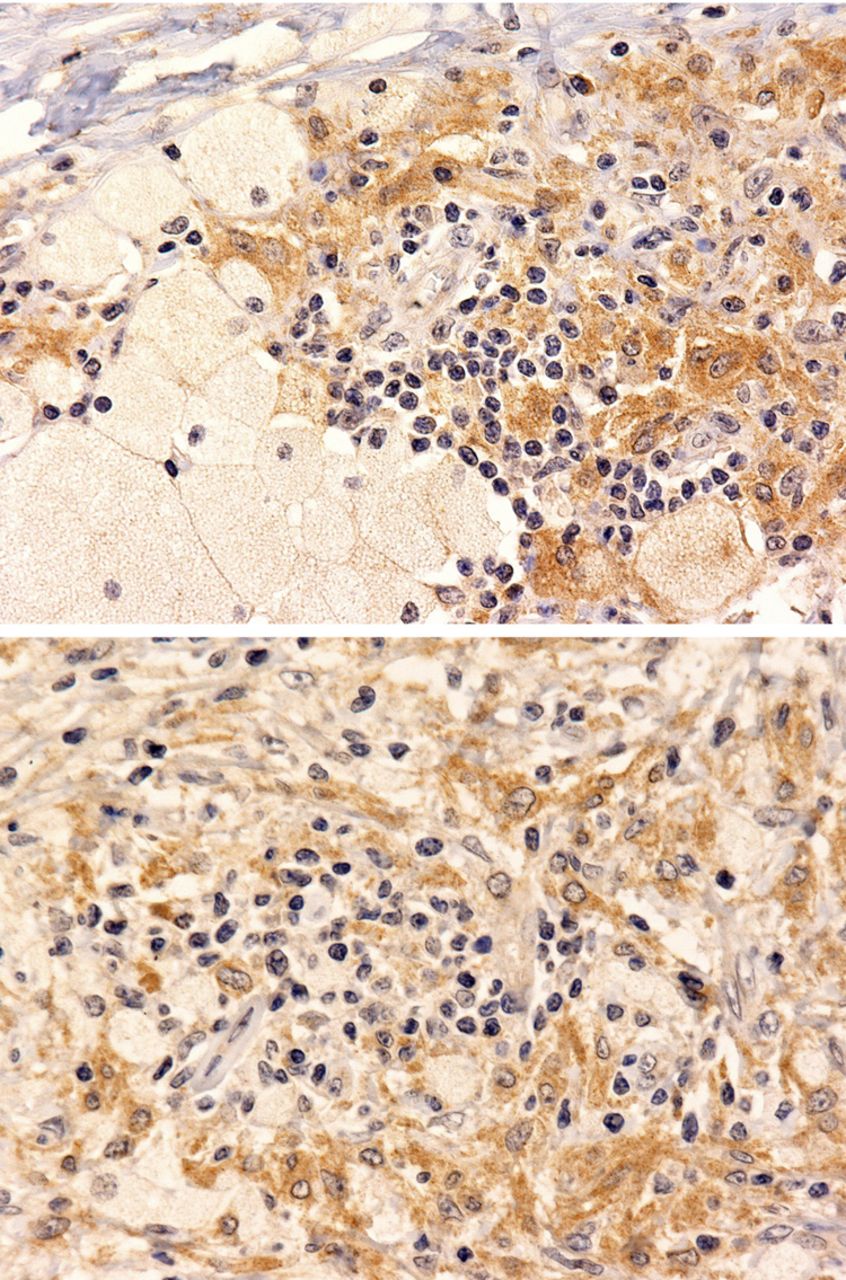
OIS is emerging as an important programme of cellular resistance against uncontrolled proliferation and neoplastic transformation, particularly in the context of BRAFV600E-mutated lesions. OIS is characterised by the expression of high levels of oncosuppressor proteins, such as p16Ink4a and p21, which act as negative regulators of the cell cycle.27 To evaluate the possible presence of an active OIS programme in ECD cells, we performed immunohistochemical analyses on consecutive slices from the same biopsy in every patient from our cohort and found that BRAFV600E-positive histiocytes were also positive for the senescence marker p16Ink4a (figure 3).

{kind=link}
{kind=link}
{kind=link}
BRAFV600E-mutated histiocytes in Erdheim-Chester disease (ECD) lesions express the senescence marker p16Ink4a. Immunohistochemical analyses on consecutive sections from a retro-orbital ECD lesion shows that BRAFV600E-positive histiocytes (Panel A, VE1 immunostaining with diaminobenzidine and haematoxylin counterstain, original magnification ×200) were also positive for the senescence marker p16Ink4a (Panel B, p16Ink4a immunostaining with diaminobenzidine and haematoxylin counterstain, ×200).
Discussion
Recent advances in understanding ECD pathogenesis suggest that at least two distinct but possibly interconnected events, that is, a full-blown chronic inflammation and the occurrence of an oncogenic mutation in histiocytes, are implicated. Our present data support a central role of BRAFV600E mutation in ECD and suggest OIS as the possible link between the two aforementioned pathogenic hypotheses.
Infiltrating histiocytes from a significant proportion of patients with ECD have been reported to bear a mutation in the proto-oncogene BRAF: in particular, Haroche and colleagues15 found BRAFV600E mutation by direct pyrosequencing in 19 out of 37 (51%) patients with ECD and in 11 out of 29 (38%) patients with LCH.15–17 Using the same technique, we could detect BRAFV600E in histological samples from a non-significantly different fraction (12 out of 18, 67%) of patients with ECD. Notably, when we exploited an ultrasensitive technique in which pyrosequencing was preceded by LNA-PCR, we were able to detect the mutation in all the biopsy samples. It is tempting to speculate that a low fraction of BRAFV600E-mutated histiocytes over wild type cells (non-mutated histiocytes and other infiltrating inflammatory cells) may lead to significantly underestimate the BRAF status of the lesion. By means of the same assay, we were also able to detect BRAFV600E in PBMCs from all the patients of our cohort (16 out of 16) and in none of the controls. To further confirm these findings we exploited an alternative ultrasensitive approach: indeed, BRAFV600E was detected in all the studied samples (biopsies and PBMCs) from patients with ECD and absent in PBMCs from healthy control also by means of ddPCR.
The data on PBMC were also supported by the finding that a small fraction of circulating monocytes obtained from patients with ECD but not from controls stained positive by flow cytometry for phospho-ERK, the major downstream target of BRAF activation. Also, single scattered cells from MDM cultures from patients with ECD but not from controls were positive for the BRAFV600E-mutated protein at immunohistochemistry. We can speculate that mutated circulating monocytes, recruited to lesion sites through the described cytokine/chemokine network, might contribute to the multifocal diffusion of ECD.
The aberrant BRAFV600E isoform is characterised by a conformational change that leads to a chronically activated mitogen-activated protein kinase pathway. This signal transduction cascade plays a fundamental role in several cellular activities, including proliferation, differentiation and eventually neoplastic transformation. Although BRAFV600E mutation has been mostly studied for its ability to induce proliferation in mutated cells, it has also been associated with OIS, a recently identified major protective mechanism against oncogenic transformation.27 In OIS, the activation of a specific oncogene in the absence of additional cooperating mutations does not end up in cell proliferation. OIS activated by BRAFV600E is associated with the expression of senescence markers such as p16Ink4a, a major tumour suppressor protein.27 By means of immunohistochemistry, we showed that OIS programmes are activated in histiocytes bearing the BRAFV600E mutation inside ECD lesions. Intriguingly, OIS has been in turn associated with the induction of SASP, a proinflammatory response characterised by the same pattern of proinflammatory chemokines and cytokines that is known to be expressed by ECD histiocytes.9–11 ,14 ,34 ,35 SASP has also been shown to reinforce cellular senescence in an autocrine fashion, and to be responsible for the induction of senescence and of a senescent cellular and secretory phenotype in bystanding cells, and for the recruitment of inflammatory cells to lesion sites via soluble factors.30 ,36 ,37 It is likely that BRAFV600E-mutated histiocytes could attract inflammatory cells to ECD lesion sites and transfer the inflammatory/senescent phenotype to by-standing non-mutated cells. These observations support the hypothesis that BRAFV600E is responsible for the inflammatory alterations observed in ECD through the activation of OIS and SASP programmes. This hypothesis explains why only a limited fraction of inflammatory cells in ECD lesion bares the BRAFV600E mutation, and reconciles the dichotomy between alternative pathogenic hypotheses and explains why several previous therapeutic approaches interfering with single downstream alterations yielded unsatisfactory results. Also, since ECD lesions are composed of BRAFV600E-mutated histiocytes variably interspersed with non-mutated chemoattracted histiocytes, these findings may explain why past clonality studies on ECD yielded conflicting results.
Taken altogether, our data suggest that BRAFV600E mutation can be demonstrated by ultrasensitive molecular biology techniques in virtually all patients with ECD. This finding holds significant clinical relevance given the availability of several drugs selectively inhibiting BRAF or its downstream kinases that could represent potential therapeutic tools for severe, refractory forms of ECD.38–41 The recent report of a single case of a patient with BRAF wild type, neuroblastoma RAS viral oncogene homolog (NRAS)-mutated ECD further substantiates the central role of the mitogen-activated protein kinase pathway in the disease, and underscores its relevance as a therapeutic target.42 Moreover, since BRAFV600E mutation in histiocytes seems to contribute to ECD pathogenesis through OIS and SASP, it is tempting to speculate that drugs that are able to lethally target senescent cells could also be evaluated as possible future treatments for ECD.43 Finally, our findings suggest that the detection of the BRAFV600E mutation in PBMC can be exploited as a potential new diagnostic tool in ECD, complementing imaging techniques and histology. The availability of a straightforward, rapid, sensitive, specific and relatively inexpensive diagnostic tool may prove beneficial for physicians and patients, and may help overcoming the need for invasive diagnostic procedures.
Acknowledgments
The authors thank Lorenza Pecciarini, Elisabetta Ferrero and Daniela Belloni for their critical support and advices, Roberto Cairella, Stefano Grassi and Francesca Invernizzi for their technical assistance. The authors also thank Kathy Brewer and the ECD Global Alliance (De Ridder, LA) for their continuous activities for patients with Erdheim-Chester disease and their unceasing support to a better understanding of this neglected disease.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
Handling editor Tore K Kvien
Contributors MGC, MF, CD and LD designed research; RB, GC, GG, ED-C, CC, BG, AB, VL performed research and acquired data; LD performed statistical analysis; MGC, RB, GC, VL, AvD, MGS, MF, CD, LD analysed and interpreted data; RB, GC, MF and LD wrote the first draft of the manuscript; and all authors critically revised the manuscript and approved its final version.
Funding Supported in part by a grant from the Italian Ministry of Health to LD (GR-2009–1594586).
Competing interests AvD holds a patent on the diagnostic use of BRAF V600E mutant-specific antibody VE1.
Provenance and peer review Not commissioned; externally peer reviewed.