Article Text

Abstract
Background: Interleukin 17 (IL17) is produced by activated T cells and has been implicated in the development of bone lesions and cartilage degradation in rheumatoid arthritis (RA).
Objective: To determine whether IL17, alone or together with tumour necrosis factor α (TNFα), induces cartilage destruction in vitro.
Methods: Fetal mouse metatarsals stripped of endogenous osteoclast precursors were used to study the effect of IL17 on cartilage degradation independently of osteoclastic resorption. Cartilage destruction was analysed histologically by Alcian blue staining.
Results: IL17 alone, up to 100 ng/ml, had no effect on the cartilage of fetal mouse metatarsals. IL17 (≥0.1 ng/ml), however, induced severe cartilage degradation when given together with TNFα (≥1 ng/ml). The cytokine combination decreased Alcian blue staining, a marker of proteoglycans, throughout the metatarsals and induced loss of the proliferating and early hypertrophic chondrocyte zones. TNFα alone also decreased Alcian blue staining, but not as dramatically as the cytokine combination. In addition, it did not induce loss of chondrocyte zones. Treatment with inhibitors of matrix metalloproteinase (MMP) activity and nitric oxide synthesis showed that MMP activity played a part in cartilage degradation, whereas nitric oxide production did not.
Conclusions: IL17, together with TNFα, induced cartilage degradation in fetal mouse metatarsals in vitro. IL17 may, therefore, participate in the development of cartilage destruction associated with RA by enhancing the effects of TNFα and may provide a potential therapeutic target.
- interleukin 17
- tumour necrosis factor α
- cartilage
- matrix metalloproteinase
- DMSO, dimethylsulphoxide
- FCS, fetal calf serum
- IL, interleukin
- l-NMMA, NG-monomethyl-l-arginine
- MMP, matrix metalloproteinase
- NO, nitric oxide
- RA, rheumatoid arthritis
- Th, T helper
- TNFα, tumour necrosis factor α
- TRAP, tartrate resistant alkaline phosphatase
Statistics from Altmetric.com
- DMSO, dimethylsulphoxide
- FCS, fetal calf serum
- IL, interleukin
- l-NMMA, NG-monomethyl-l-arginine
- MMP, matrix metalloproteinase
- NO, nitric oxide
- RA, rheumatoid arthritis
- Th, T helper
- TNFα, tumour necrosis factor α
- TRAP, tartrate resistant alkaline phosphatase
Interleukin 17 (IL17) is a cytokine which is produced exclusively by activated memory CD4+ T cells.1 T cells are currently thought to have a central role in the initiation and perpetuation of rheumatoid arthritis (RA).2–5 It has been suggested that CD4+ memory T helper (Th) cells, especially, are pivotal.6,7 IL17 bioactivity has been found in synovium supernatants,8 and high levels of IL17 have been reported in the synovium of patients with RA.9–12
Recent studies have shown that IL17 inhibited proteoglycan synthesis in intact murine articular cartilage in vivo13 and that IL17 producing Th1 cells were among the most potent inhibitors of collagen synthesis by synoviocytes in vitro.14 Furthermore, treatment of RA synovium with anti-IL17 blocking antibody reduced the release of type 1 collagen C-telopeptide fragments in the supernatant,15 and intra-articular injection of IL17 induced joint inflammation and cartilage proteoglycan depletion.16–18 We19,20 and others11,16,21 have reported that IL17 increases osteoclastic bone resorption either alone or in combination with tumour necrosis factor α (TNFα), a major pathogenic factor in RA. In this study we investigated whether IL17, alone or in combination with TNFα, may in addition induce cartilage destruction. Potential mediators are nitric oxide (NO) and matrix metalloproteinases (MMPs), because they are found in excess in the synovium of patients with RA and are mediators of cartilage degradation.22–28
To study the effects of IL17, alone or in combination with TNFα, on cartilage, we used an experimental model of fetal mouse bone explants that contain both bone and cartilage. These bone explants produce NO in response to cytokine stimulation, contain matrix MMP activity, and are responsive to inhibitors of NO synthesis and MMP activity.29,30
METHODS
Materials
Alpha minimal essential medium was purchased from Gibco BRL (Breda, The Netherlands), penicillin and streptomycin from Flow Laboratories (Amstelstad, Zwanenburg, The Netherlands), collagenase from Worthington Biochemicals Corp (Freehold, NJ, USA), and fetal calf serum (FCS) from Integro (Zaandam, The Netherlands). Recombinant human interleukin 17 was obtained from R&D Systems (Abingdon, UK), NG-monomethyl-l-arginine (l-NMMA) from Calbiochem (San Diego, CA, USA), and dimethylsulphoxide (DMSO) from Sigma-Aldrich Chemie BV (Zwijndrecht, The Netherlands). Recombinant mouse TNFα was a generous gift of Dr A van de Voorde (Innogenetics, Antwerp, Belgium), and marimastat of Dr A Baxter (Celltech Inc, Cambridge, UK).
Organ cultures
Pregnant Swiss albino mice were killed by cervical dislocation at day 17 or 19 of gestation. Fetal metatarsals were aseptically excised in phosphate buffered 0.9% NaCl containing 10% heat inactivated FCS. 17 Day old fetal mouse metatarsals do not yet contain mature osteoclasts, but only osteoclast precursors, which are still confined to the periosteum. These cells can be removed together with the periosteum by collagenase treatment as described previously.31 Osteoclast-free 17 day old and osteoclast-containing 19 day old fetal metatarsals were cultured in 250 μl alpha minimal essential medium supplemented with 10% heat inactivated FCS in 24 well plates (Costar, Badhoevedorp, The Netherlands) in a humidified atmosphere of 5% CO2 and 95% O2 at 37°C. This animal protocol was approved by the Leiden University Committee for Animal Experiments (UDEC).
Histological staining
Fetal mouse metatarsals were fixed in 10% neutral buffered formalin for four hours at 4°C. For tartrate resistant alkaline phosphatase (TRAP) staining metatarsals were subsequently decalcified in 5% formic acid and 5% formalin overnight at 4°C. Metatarsals were embedded in paraffin and sections 5 μm thick were cut. Subsequently, the sections were analysed for TRAP, Alcian blue, or von Kossa staining. Osteoclasts were stained with TRAP as a marker. For this, naphthol ASBI phosphate was used as substrate, p-rosaniline as coupler, and 30 mM l+-tartaric acid as inhibitor.32 Sections were counterstained with Mayer's haematoxylin. Proteoglycans in the cartilage matrix were stained with Alcian blue33 and sections were counterstained with “kernechtrot”. Mineralised matrix was stained according to the method of von Kossa using AgNO3 as reagent.34 Sections were counterstained with methyl green.
Measurement of bone area and length of metatarsals
Macroscopical analysis of bone area and length of the metatarsals was carried out by computerised analysis. Bone explants were projected using a microscope. Drawings of the outline of the explants were scanned with a Hewlett Packard ScanJet 5p scanner and Corel PhotoPaint 6.0 (Corel Corp, Ottawa, Ontario, Canada). Images were analysed for bone area and length using Image Pro Plus 3.0 for windows 95/NT (Media cybernetics, Maryland, USA). Bone area was quantified as arbitrary units within the outline of the explants and bone length as arbitrary units along the medial axis of the explants.
Nitrite production
NO production was measured as nitrite secretion into the medium by using Griess reagent consisting of 0.5% sulphanilamide, 0.05% naphthylethylene-diamine-dihydrochloride, and 2.5% H3PO4.30 Briefly, 70 μl culture supernatant was mixed with 50 μl Griess reagent and incubated in a 96 well plate for 10 minutes at room temperature under continuous shaking. Nitrite concentration, proportional to the optical density at 550 nm (OD550), was determined using a microtitre plate reader (Thermomax, Molecular Devices, Menlo Park, CA, USA) with a 650 nm reference filter and with reference to a standard curve of serial dilutions of sodium nitrite.
Statistics
Values are expressed as mean (SEM). Statistical differences between values were examined by one way analysis of variance, followed by Fischer's protected least significant difference test and considered to be significantly different at p<0.01.
RESULTS
Effect of IL17 and TNFα on cartilage morphology of 19 day old fetal mouse metatarsals
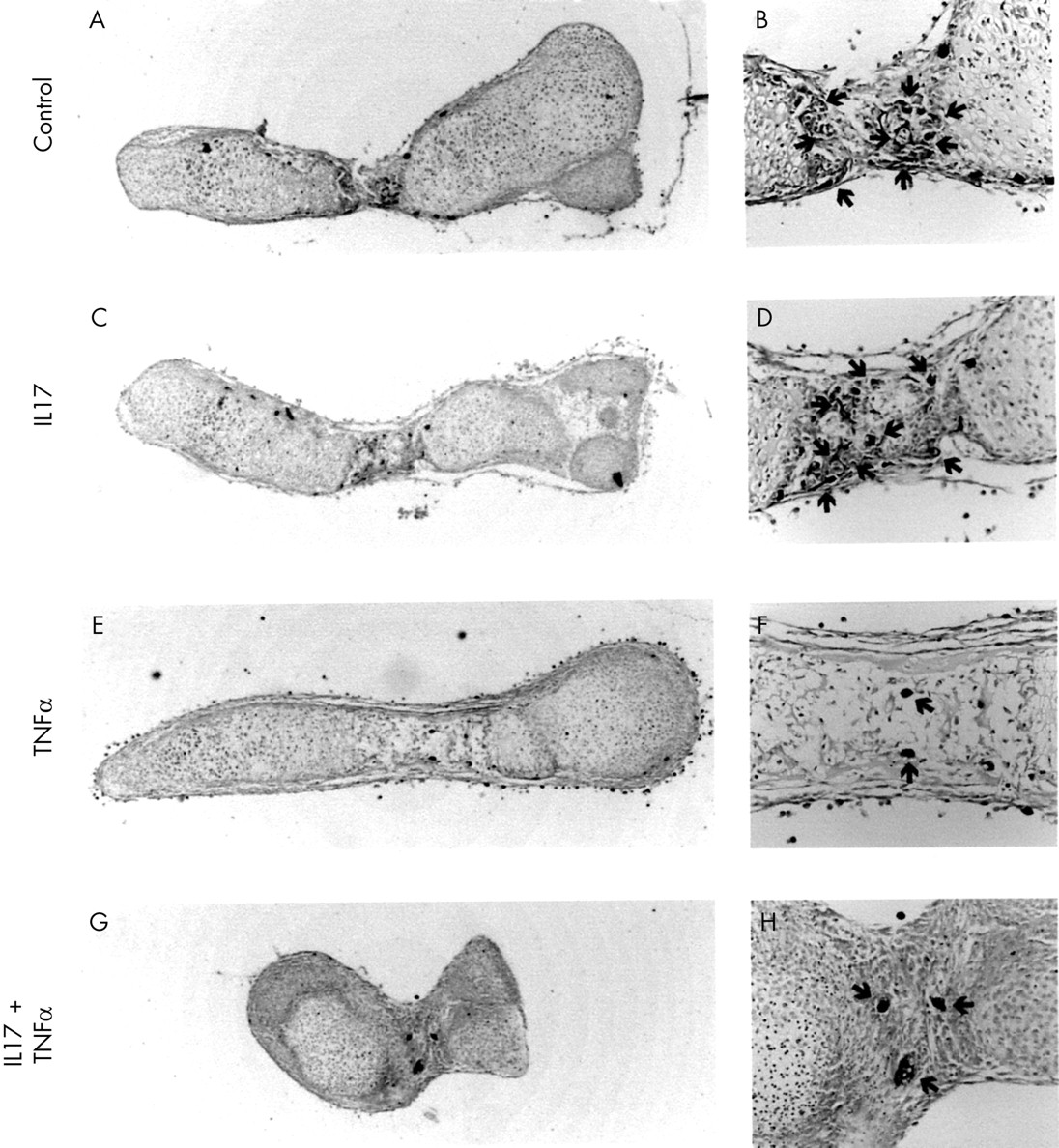
Neither IL17 nor TNFα, at concentrations up to 100 ng/ml, had any effect on cartilage morphology of osteoclast-containing 19 day old fetal mouse metatarsals after seven days of culture (fig 1). The combination of these two cytokines, however, induced shrinkage and folding of the metatarsals, which was evident already from day 4 of culture. Neither IL17 nor TNFα had any affect on bone area and length of these metatarsals (data not shown), whereas the cytokine combination decreased both parameters dose dependently (table 1). A similar synergistic effect between IL17 and TNFα was found with 10 ng TNFα/ml and IL17 concentrations of 1 ng/ml and higher. Mononuclear and multinuclear osteoclasts, visualised using TRAP staining as a marker, were present in control and IL17 treated cultures, but their number was strongly reduced in cultures treated with TNFα (fig 1). Simultaneous addition of IL17 and TNFα prevented a reduction in TRAP+ osteoclasts and restored resorption of mineralised hypertrophic cartilage. These effects are in agreement with our previous findings on osteoclastic resorption.19
The cytokine combination IL17 + TNFα, but not the cytokines alone, decreased bone area and length of fetal mouse metatarsals

Histological staining of 19 day old fetal mouse metatarsals cultured in the absence or presence of IL17, TNFα, or a combination of the two cytokines. 19 Day old fetal mouse metatarsals were cultured for seven days in the absence (A, B) or presence of IL17 (100 ng/ml) (C, D), TNFα (100 ng/ml) (E, F), or a combination of the two cytokines (G, H). The presence of mononuclear and multinuclear osteoclasts was visualised using TRAP staining. Magnification: A, C, E, G ×40; B, D, F, H ×200. Cultures were performed in quintuplicate (n=5).
Effect of IL17 and TNFα on cartilage morphology of osteoclast-free 17 day old fetal mouse metatarsals
In 19 day old explants the observed decrease in bone area and length induced by the cytokine combination may be due to resorption caused by the osteoclasts. To study this further we used 17 day old fetal mouse metatarsals which do not yet contain mature osteoclasts, but only osteoclast precursors which are still confined to the periosteum. The osteoclast precursors were then enzymatically and mechanically removed. Any changes seen in these metatarsals are, therefore, independent of osteoclastic resorption. In these explants also, IL17 or TNFα given alone at concentrations up to 100 ng/ml did not change the area and length of the explants compared with controls (data not shown). However, when the two cytokines were given together they induced a dramatic decrease in bone area and length (table 1). In contrast with 19 day old fetal mouse metatarsals, however, growth of the metatarsals occurred normally during the first week of culture and shrinkage and folding were evident only in the second week. The findings in the osteoclast-free 17 day old fetal mouse metatarsals thus provide a new experimental model to study effects on cartilage independent of osteoclastic resorption. All further experiments were, therefore, performed using osteoclast-free 17 day old fetal mouse metatarsals.
Effect of IL17 and TNFα on proteoglycans in the cartilage matrix
In osteoclast-free 17 day old fetal mouse metatarsals, all chondrocyte zones contained proteoglycans, as shown by Alcian blue staining (fig 2A). During culture, development of the metatarsals continued, resulting in larger bone explants still containing all chondrocyte zones after 11 days of culture. Treatment with IL17 had no effect on Alcian blue staining and cell morphology, but TNFα decreased Alcian blue staining in the early hypertrophic and proliferating chondrocyte zones (fig 3). The combination of the two cytokines had a dramatic effect, decreasing Alcian blue staining throughout the metatarsals. In addition, it induced complete disappearance of the early hypertrophic and proliferating chondrocyte zones; which was associated with folding and shrinkage of the metatarsals.

Zones of chondrocyte differentiation in fetal mouse metatarsals. 17 Day old fetal mouse metatarsals were fixed directly after isolation and stained for Alcian blue (A) or von Kossa (B). Four different zones of chondrocyte differentiation are shown: lhc, mineralised late hypertrophic chondrocytes (black von Kossa staining); ehc, unmineralised early hypertrophic chondrocytes; pc, proliferating chondrocytes; rc, resting chondrocytes. Magnification ×40.
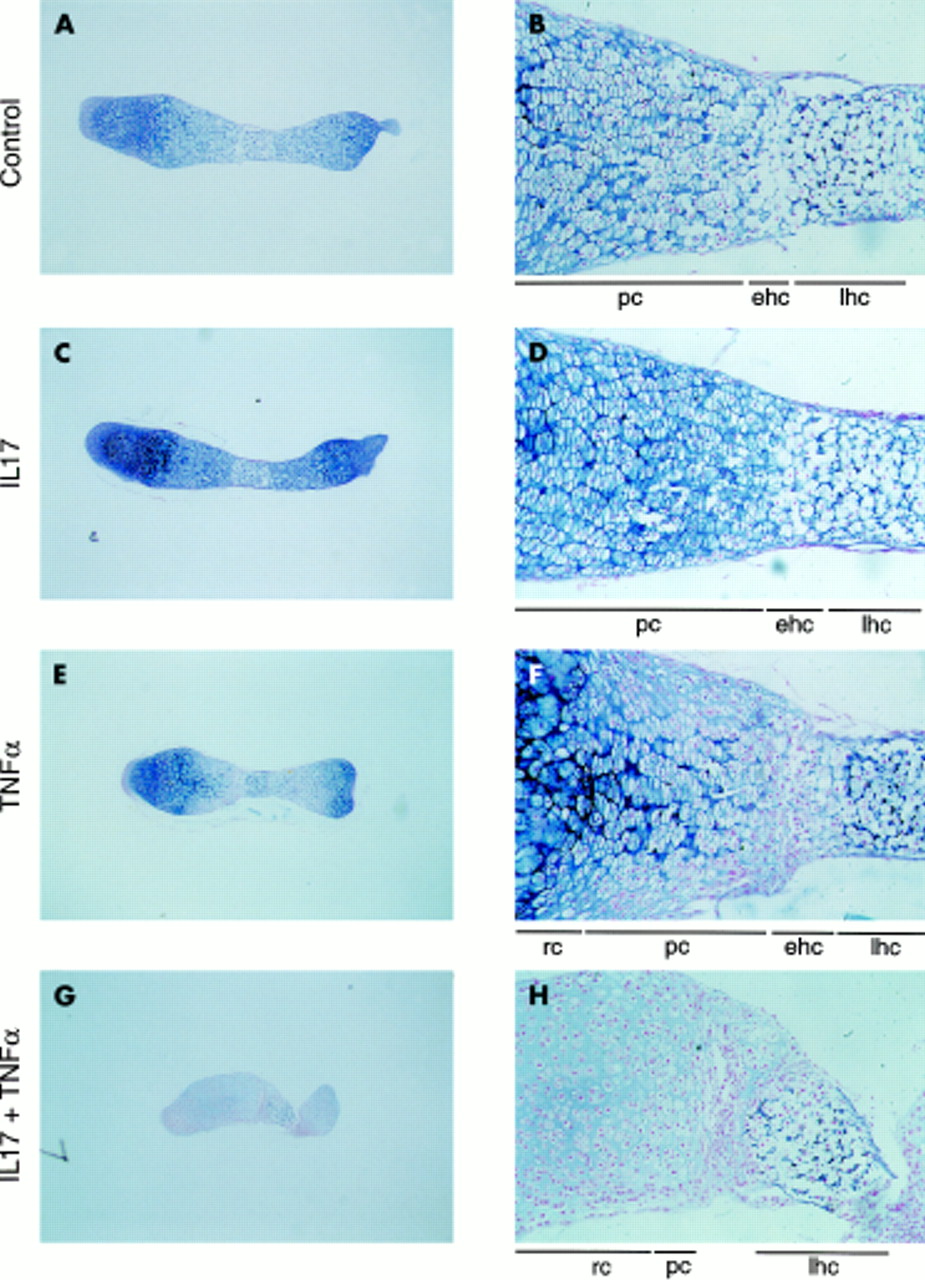
Effect of IL17, TNFα, and a combination of the two cytokines on Alcian blue staining. Osteoclast-free 17 day old fetal mouse metatarsals were cultured for 11 days in the absence (A, B) or presence of IL17 (50 ng/ml) (C, D), TNFα (50 ng/ml) (E, F), or a combination of the two cytokines (G, H) and stained with Alcian blue. lhc, late hypertrophic chondrocytes; ehc, early hypertrophic chondrocytes; pc, proliferating chondrocytes; rc, resting chondrocytes. Magnification: A, C, E, G ×40; B, D, F, H ×200. Cultures were performed in quadruplicate (n=4).
Effect of IL17 and TNFα on mineralisation of the cartilage matrix
Von Kossa staining showed that late hypertrophic chondrocytes in the centre of 17 day old fetal mouse metatarsals had formed a mineralised matrix (fig 2B). At both sites of this mineralised matrix, there is a small zone of early unmineralised hypertrophic chondrocytes. During culture, mineralisation of cartilage was not affected by IL17 (fig 4). TNFα disturbed mineralisation as shown by irregular von Kossa staining of the metatarsals. The combination of the two cytokines inhibited further mineralisation of the cartilage matrix during the second week of culture.

Effect of IL17, TNFα, and a combination of the two cytokines on von Kossa staining. Osteoclast-free 17 day old fetal mouse metatarsals were cultured for 11 days in the absence (A) or presence of IL17 (50 ng/ml) (B), TNFα (50 ng/ml) (C), or a combination of the two cytokines (D) and stained with von Kossa. Magnification ×100. Cultures were performed in quadruplicate (n=4).
Role of NO and MMPs in IL17 + TNFα-induced cartilage degradation
The role of NO production in the histological changes induced by the cytokine combination was studied using l-NMMA, an inhibitor of NO synthesis. While l-NMMA (1 mM) completely blocked the IL17 + TNFα stimulation of NO production (from 3.25 nmol/metatarsal to zero), it did not prevent any of the histological changes induced by the cytokine combination (fig 5). l-NMMA alone had no effect on the morphology of the metatarsals. Metatarsals in untreated cultures were similar to those shown in figs 3A and 4A.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effect of an NO synthesis inhibitor and an MMP inhibitor on cartilage destruction induced by IL17 + TNFα. Osteoclast-free 17 day old fetal mouse metatarsals were treated with IL17 (100 ng/ml) + TNFα (100 ng/ml) for 11 days in the presence of the nitric oxide synthesis inhibitor l-NMMA (1 mM) (A, B) or the MMP inhibitor marimastat (10−6 M) (C, D) and stained with Alcian blue (A, C) or von Kossa (B, D). Marimastat was dissolved in DMSO and the final concentration of DMSO in culture medium was 0.05%. This DMSO concentration had no effect on the histological picture of control and cytokine treated cultures (data not shown). DMSO, dimethylsulphoxide. Magnification ×40. Cultures were performed in quadruplicate (n=4).
Involvement of MMP activity in the histological changes induced by the cytokine combination was studied using marimastat, a broad spectrum inhibitor of MMP activity. Marimastat (10−6 M) prevented IL17 + TNFα-induced folding of the metatarsals and loss of the early unmineralised hypertrophic and proliferating chondrocyte zones. It did not, however, prevent the decrease in Alcian blue staining nor the inhibition of mineralisation. Marimastat alone had no effect on the morphology of the metatarsals.
DISCUSSION
In this study we showed that the cytokine IL17 in combination with TNFα induces cartilage destruction in fetal mouse metatarsals in culture. These bone explants have been successfully used in the study of bone for many years to examine the effect of multiple agents on osteoclast formation and activity.19,20,29–31 This was feasible because of the different stages of osteoclast development present in these metatarsals depending on the age of the fetus. At day 17 of gestation, no mature osteoclasts but only precursors which are still confined to the periosteum are present. After removal of this periosteum, 17 day old fetal mouse metatarsals consist only of cartilage with all stages of chondrocyte differentiation present, as also shown here. In these explants, therefore, effects on cartilage can be studied directly and independently of osteoclastic resorption. This provides an in vitro model to study the effects of agents on cartilage. It should be mentioned that articular cartilage is different from cartilage in these explants in that it is an acellular tissue with hardly any regenerative ability.35 We show here that IL17 in combination with TNFα induced cartilage destruction shown by decreased Alcian blue staining, a marker of proteoglycans, throughout the bone explants and shown also by loss of proliferating and early hypertrophic chondrocyte zones during the second week of culture. IL17 alone had no effect, whereas TNFα decreased Alcian blue staining in the proliferating and early hypertrophic chondrocyte zones. The latter effect, however, was not as dramatic as that induced by the combination with IL17 and it did not induce loss of proliferating and early hypertrophic chondrocytes. We are aware of the limited quantitative value of histological data, but the very small size (±2 mm) of the fetal mouse metatarsals made it impossible to measure degradation products of proteoglycans and collagens in the media using various methods.
Both IL17 and TNFα have been found in the synovium of patients with RA,10–12 and antibody treatment against TNFα decreased disease activity.36 In vitro, IL17 has been found to be produced by synovium samples8 and Th1/Th0 clones, but not Th2 clones, generated from synovial membranes and fluids37 of patients with RA. Moreover, IL17 has been shown to inhibit proteoglycan and collagen synthesis13,14,16,18 and to induce collagen and proteoglycan degradation.15–18 These data, together with the present finding that IL17 and TNFα synergistically induced cartilage degradation in cultures of fetal mouse metatarsals, suggest that the combination of both cytokines may be of importance in the progressive destruction of cartilage associated with RA. A cartilage degenerating effect of IL17 alone may also be possible, because IL17 stimulated proteoglycan release in mouse cartilage explants of patellae16 and porcine articular cartilage explants.18 Differences in type of cartilage and species may be responsible for the need for synergism with TNFα or not.
It has been reported that NO is involved in cartilage degradation either directly or indirectly through the stimulation of other factors, for example MMPs, and NO has been associated with the development of RA.22–28 Furthermore, IL17 induced NO production in human chondrocytes38,39 and fetal mouse metatarsals.20 Under the experimental conditions used here, however, NO production induced by IL17 + TNFα was not involved in cartilage destruction. This was shown using the NO synthesis inhibitor l-NMMA which could not prevent the changes induced by the cytokine combination. On the other hand, our data strongly suggest that MMPs play a part in IL17 + TNFα-induced cartilage destruction. MMP activity may mediate the effect of the cytokine combination on cartilage, because MMPs degrade cartilage23 and MMPs are induced by IL17 in human chondrocytes,40 monocytes/macrophages,10,41 and synoviocytes.15 Furthermore, treatment with anti-IL17 blocking antibody reduced MMP-1 production and collagenase activity by RA synovium.15 MMP-13 mRNA expression has been reported in the hypertrophic chondrocytes of 17 day old fetal mouse metatarsals, and the explants have been found to respond to MMP inhibitors.29 We show that marimastat, a broad spectrum MMP inhibitor that has been used in clinical trials in the field of oncology,42,43 prevented IL17 + TNFα-induced folding of metatarsals and loss of early hypertrophic and proliferating chondrocyte zones, but it did not decrease Alcian blue staining. Although inhibition of other metalloproteinases by marimastat cannot be excluded, the data strongly suggest that MMP activity plays a part in the destruction of the early hypertrophic and proliferating chondrocyte zones, but not in the degradation of proteoglycans. Similarly, it has been suggested that IL17-induced proteoglycan release is due to aggrenase activity, but not MMP activity, in bovine and porcine articular cartilage.18 Furthermore, degradation of collagen by IL1 was mediated by MMP activity in bovine nasal cartilage explants, whereas degradation of proteoglycans was not.44–47
In conclusion, we showed that IL17 in combination with TNFα, in addition to its stimulating effect on osteoclastic resorption, also induces cartilage destruction in fetal mouse bone explants. This synergism of the two cytokines may be responsible for the progressive cartilage destruction and bone lesions in RA. IL17 may, therefore, participate in the pathogenesis of RA by enhancing the effects of TNFα, and presents a potential therapeutic target.
Acknowledgments
The authors thank Professor Dr FC Breedveld, Department of Rheumatology, Leiden University Medical Centre, Leiden, The Netherlands, for critically reading the manuscript.