Article Text

Abstract
Aim MicroRNA-146a (miR-146a) is involved in certain immune-mediated diseases. Transcription factor Ets-1 strongly affects miR-146a promoter activity and directly regulates miR-146a expression. This study was performed to investigate the association of miR-146a and Ets-1 gene polymorphisms with Behçet's disease (BD) and Vogt–Koyanagi–Harada (VKH) disease in a Chinese Han population.
Methods A total of 809 patients with BD, 613 patients with VKH and 1132 normal controls were genotyped for miR-146a/rs2910164, rs57095329 and rs6864584, Ets-1/rs1128334 and rs10893872 using a PCR restriction fragment length polymorphism assay. miR-146a expression was examined in peripheral blood mononuclear cells (PBMCs) by real-time PCR. Cytokine production by PBMCs was measured by ELISA.
Results A significantly decreased frequency of the homozygous rs2910164 CC genotype and C allele was observed in patients with BD compared with controls (pca=1.24×10−5, OR 0.61; pca=1.33×10−4, OR 0.75, respectively). MiR-146a expression in GG cases was 2.45-fold and 1.99-fold higher, respectively, than that in CC cases and GC cases. There was no association of the other four single nucleotide polymorphisms (SNPs) with BD. There was also no association of these five SNPs with its main clinical features. No associations were found with the five SNPs tested or with its clinical manifestations in VKH disease. Interleukin (IL)-17, tumour necrosis factor (TNF)α and IL-1β production from rs2910164 CC cases was markedly lower than that in GG cases. No effect of genotype was observed on IL-6 and monocyte chemoattractant protein (MCP)-1 production and IL-8 expression was slightly higher in CC cases.
Conclusions Our study identified a strong association of rs2910164 of miR-146a with BD in a Chinese population and decreased expression of miR-146a and certain proinflammatory cytokines in individuals carrying the CC genotype.
- Behcet's disease
- Autoimmune Diseases
- Inflammation
Statistics from Altmetric.com
Introduction
Behçet's disease (BD) is a chronic, multisystem inflammatory disease. It is more prevalent in the region that extends from China and Japan in the Far East to the Mediterranean sea.1 ,2 Vogt–Koyanagi–Harada (VKH) disease is an autoimmune disorder characterised by bilateral granulomatous panuveitis frequently associated with systemic involvement and frequently affects Asians and Native Americans.3 ,4 Although the exact aetiology and pathogenesis of both diseases remain unclear, the role of genetic factors in their pathogenesis has long been recognised.5–7 Various studies have reported that human leukocyte antigen (HLA) genes, such as HLA-B51, and several non-HLA genes including IL23R-IL12RB2 and IL10 genes are associated with BD.8–10 HLA-DR4 and IL-17 polymorphisms have been found to be associated with VKH disease.11 ,12 These reports suggest that genetic factors may play a role in these two diseases.
Micro RNAs (miRNAs) have been recently recognised as an important regulator in immune homeostasis. Abnormalities in the miRNA-mediated regulation of immune-cell development and function have been linked to autoimmunity and inflammatory diseases.13 ,14 Tang et al15 demonstrated that miR-146a was a negative regulator of innate immunity in patients with systemic lupus erythematosus (SLE). Recent studies have demonstrated that single nucleotide polymorphisms (SNPs) located either in the pre-miRNAs or within miRNA binding sites are likely to affect the expression of the miRNA targets and, thus, may contribute to the susceptibility to diseases.16 ,17 V-Ets oncogene homologue 1 (Ets-1), the first member of the Ets oncogene family, has been identified in the avian erythroblastosis virus E26 and shown to contribute to tumour development and progression.18 Recently, Luo et al13 identified that Ets-1 binds to the miR-146a promoter region and strongly affected miR-146a promoter activity in vitro. In vivo experiments have shown that the knockdown of Ets-1 directly impaired the induction of miR-146a.13 It has recently also been shown that SNP rs1128334 and rs10893872 located in the 3′-untranslated region (3′-UTR) of Ets-1 are on putative miRNA binding sites and are both associated with SLE in Asian populations.19 All these results suggested that Ets-1 influences the expression and activity of miR-146a.
In view of the important role of miR-146a in the development of inflammation-related and autoimmune diseases and the potential modulatory effect of Ets-1 on miR-146a expression, we examined whether polymorphisms of both genes were associated with BD and, if so, whether the variant affected gene expression and relevant cytokine production. VKH disease was chosen as a disease control mainly based on a similar prevalence in Chinese population and the similar clinical features, such as recurrent episodes and difficulty in the treatment although they are different in disease nature (BD, non-granulomatous inflammation; VKH disease, granulomatous inflammation).
Materials and methods
Study population
The study group comprised 809 consecutive patients with BD who were recruited from the First Affiliated Hospital of Chongqing Medical University (Chongqing, China) and the Zhongshan Ophthalmic Center of Sun Yat-sen University (Guangzhou, China) between October 2006 and October 2012. A total of 1132 unselected, consecutive control subjects were matched ethnically and geographically with the patients. Patients with VKH were used as disease controls. BD and VKH disease were strictly diagnosed based on the criteria of the International Study Group for BD20 and First International Workshop for VKH,21 respectively. If the diagnosis was in any doubt, the patients were excluded from the study. A Hardy–Weinberg equilibrium (HWE) calculation was performed to ensure a balance in genotype and allele distribution between patients and normal controls. A binary logistic regression analysis was applied to avoid the effect of gender on the association of the variants with BD. Direct sequencing was performed in 20% of the total samples randomly chosen in order to validate the method employed in this study. The study received the approval of the Local Ethics Research Committee and all the investigated subjects provided informed consent before collection of blood. The tenets of the Declaration of Helsinki were upheld during all procedures of this study.
DNA extraction and genotyping
Genomic DNA samples from patients with BD, patients with VKH and healthy controls were extracted by using the QIAamp DNA Blood Mini Kit (Qiagen, Valencia, California, USA). The target DNA sequence was amplified by the PCR using proper primers (see online supplementary table S1). PCR products were digested with 2 U of HSP92II (Promega, Madison, Wisconsin, USA), MspI (Fermentas, Shenzhen, China), ApaI (Fermentas, Shenzhen, China), AflII (Fermentas, Shenzhen, China) and TSP509I (Fermentas, Shenzhen, China) restriction enzymes in a 10 μl reaction volume overnight. HSP92II digested the introduced mismatch restriction sites of SNPs rs2910164 and rs1128334. MspI and AflII digested the introduced mismatch restriction sites of SNPs rs57095329 and rs17057381, respectively. ApaI and TSP509I digested the natural restriction sites of SNP rs6864584 and SNP rs10893872, respectively. The formed digestion products were separated on 4% agarose gels and stained with GoldView (SBS Genetech Beijing, China).
Cell isolation and culture
Peripheral blood mononuclear cells (PBMCs) were isolated from heparinised blood samples by Ficoll-Hypaque density-gradient centrifugation. Isolated PBMCs cells (2×106 cells per well) were seeded in 24-well plates and cultured in RPMI medium 1640 supplemented with 10% fetal calf serum (FCS, Greiner, Wemmel, Belgium), 100 U/ml penicillin, 100 μg/ml streptomycin. In order to detect interleukin (IL)-17 production, PBMCs were cultured with anti-CD3 antibody (5 μg/ml, eBioscience, SanDiego, California, USA) and anti-CD28 antibody (1 μg/ml, eBioscience) for 3 days. For tumour necrosis factor (TNF)α, IL-1β, IL-8, IL-6 and monocyte chemoattractant protein (MCP)-1 detection, PBMCs were cultured with 100 ng/ml lipopolysaccharide (LPS, 100 ng/ml; Sigma, Missouri, USA) for 24 h.
Real-time PCR
Total RNA was extracted from PBMCs using TRIzol (Invitrogen, San Diego, California, USA), followed by reverse transcription using transcriptase kit (Applied Biosystems, ABI, Foster City, California, USA). The quantity of mature miR-146a was determined by TaqMan MicroRNA Assay kit (ABI) and normalised to snRNA U6. The assays were performed on a 7500 real-time instrument (ABI). Relative expression levels were calculated using the 2−ΔΔCt method.
ELISA
The concentration of IL-17, TNFα, IL-1β, IL-8, IL-6 and MCP-1 in the supernatants of PBMCs were measured using the human Duoset ELISA development kit (R&D Systems, Minneapolis, Minnesota, USA) according to the manufacturer's protocols.
Statistical analysis
For single SNP analysis, genotype and allele frequencies were compared between patients and controls by χ2 test using SPSS V.17.0 (SPSS, Inc., Chicago, Illinois, USA). The χ2 test was also applied to analyse the HWE. A binary logistic regression analysis was used to assess the influence of gender on the association of five SNPs with BD. The non-parametric Mann–Whitney test was used to compare miR-146a expression and cytokine levels among three genotype groups. Two-tailed p values <0.05 were considered to be statistically significant.
Results
Clinical features of patients with BD and patients with VKH
Detailed clinical findings of the enrolled BD and patients with VKH are presented in table 1. Age and gender distribution in patients with VKH, patients with BD and controls are also presented in table 1.
Clinical characteristic, age and gender distribution in patients with BD and patients with VKH
Allele and genotype frequencies of SNPs in patients and controls
A total of 5 SNPs were successfully genotyped in 809 patients with BD, 613 patients with VKH and 1132 normal controls. The sequencing details are shown in online supplementary figure S1. The distribution of genotype frequencies of the five SNPs did not deviate from HWE in the controls. Our results showed that there was a significant difference between patients with BD and normal controls concerning the frequency of rs2910164. The frequency of the CC genotype and C allele of rs2910164 in patients with BD was significantly lower than that in normal controls (p=3.60×10−7, pca=1.24×10−5, OR 0.61; p=1.44×10−5, pca=1.33×10−4, OR 0.75, respectively) (tables 2 and 3). The frequency of the rs2910164 CG genotype was significantly increased in patients with BD compared with controls (p=1.89×10−4, pca=5.95×10−3, OR 1.42). There was no statistically significant difference concerning the genotype and allele of the other four SNPs between patients with BD and controls after gender correction. As BD involves various organs, we further performed a study on the association of these five SNPs with clinical findings of this disease including hypopyon, genital ulcers, skin lesions and arthritis. Our study failed to find any association of these disease parameters with the tested SNPs. The genotype and allele frequencies of rs2910164, rs57095329, rs6864584, rs1128334 and rs10893872 showed no difference between patients with VKH and normal controls (tables 2 and 3). There was also no association of these five SNPs with extraocular manifestations of VKH disease including headache, tinnitus, vitiligo, alopecia and poliosis.
Frequencies of genotypes and alleles of micro RNA (miR)-146a and Ets-1 polymorphism in patients with BD and controls
Frequencies of genotypes and alleles of micro RNA (miR)-146a and Ets-1 polymorphism in patients with VKH and controls
The influence of rs2910164 on miR-146a expression
The aforementioned result identified a strong association of SNP rs2910164 with BD. We further evaluated the expression of miR-146a in PBMCs derived from 24 healthy individuals with a known SNP (rs2910164) status. Mature miR-146a level was measured by the TaqMan microRNA assay with available RNA samples. The mean expression of miR-146a in GG cases was 2.45-fold and 1.99-fold higher than that in CC (p=0.009) and GC cases (p=0.018), respectively (figure 1).

Micro RNA (miR)-146a expression with three different genotypes of single nucleotide polymorphism (SNP) rs2910164 in peripheral blood mononuclear cells (PBMCs). TaqMan real-time PCR analysis of mature miR-146a expression in PBMCs derived from healthy individuals of SNP rs2910164 GG, GC and CC genotypes (n=8 per group). The y axis represents relative mature miR-146a expression level by real-time PCR of each genotype of SNP rs2910164. Data are shown as mean±SD. *p<0.05; **p<0.01.
The influence of rs2910164 on cytokine production
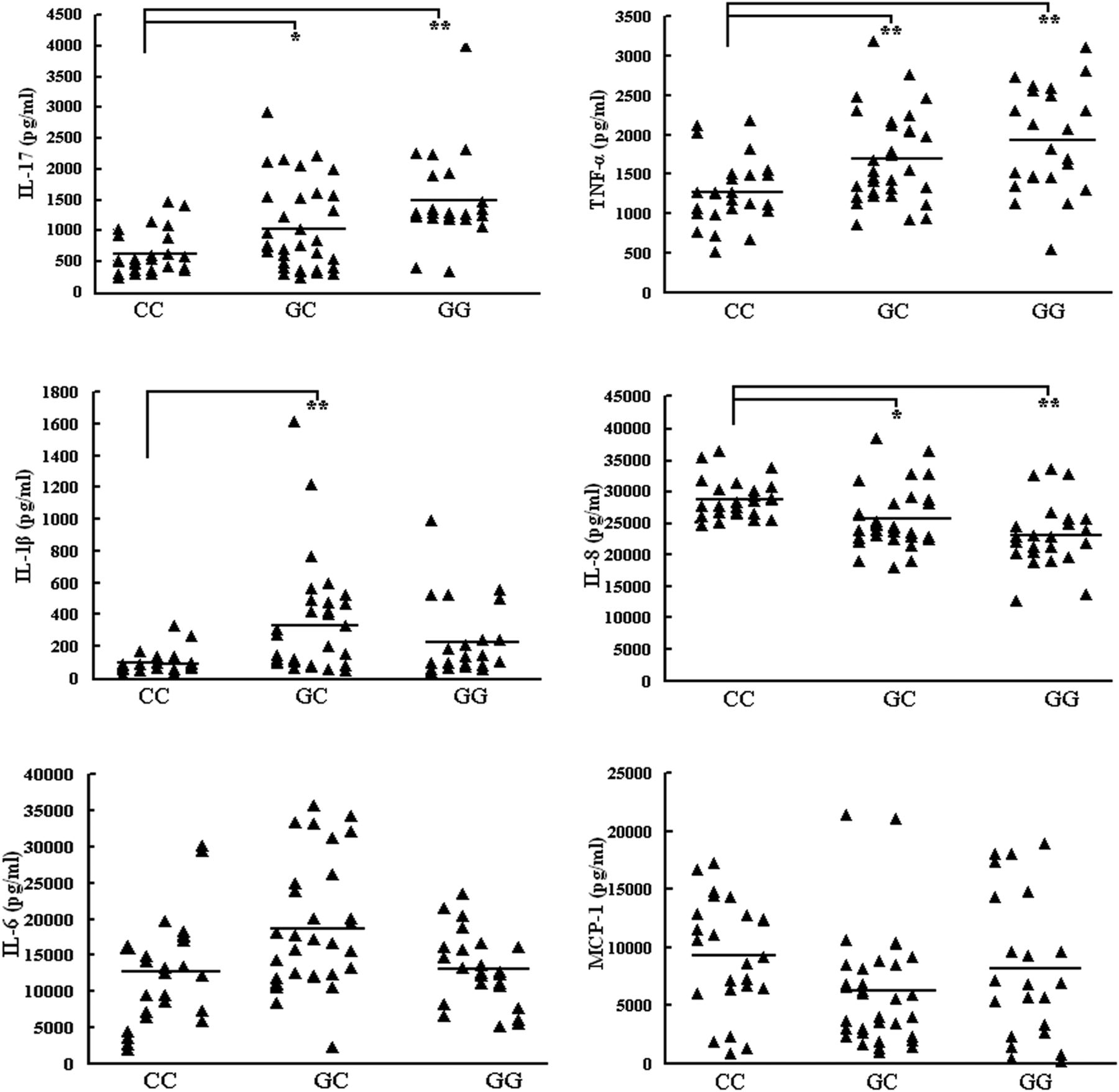
A further experiment was performed to examine different genotypes of SNP rs2910164 on cytokines important to the development of BD such as IL-17, TNFα, IL-1β, IL-8, IL-6 and MCP-1. The results showed that the IL-17 production by stimulated PBMCs from CC cases (603.57±352.73 pg/ml) were lower than that in GC (1012.55±716.65 pg/ml, p=0.028) and GG cases (1471.43±734.31 pg/ml, p<0.001), respectively. TNFα production by stimulated PBMCs from CC cases (1454.41±485.86 pg/ml) was lower than that in GC (1929.95±676.12 pg/ml, p=0.006) and GG cases (2200.98±758.27 pg/ml, p<0.001), respectively. A similar result was also present concerning the IL-1β expression between GC and CC cases. IL-1β production by stimulated PBMCs from CC cases (99.12±69.20 pg/ml) was lower than that in GC cases (328.85±309.53 pg/ml, p<0.001). There was no significant difference between GC and GG genotypes (p=0.057) and no significant association was observed concerning the IL-6 and MCP-1 production by stimulated PBMCs and the three different genotypes. However, the IL-8 expression in the supernatants of stimulated PBMCs from CC cases (28.73±3.11 ng/ml) was higher as compared to the GG (25.53±4.82 ng/ml, p=0.001) and GC cases (23.01±5.17 ng/ml, p<0.001), respectively (figure 2).
{kind=link}
{kind=link}
Cytokine production with three different genotypes of single nucleotide polymorphism (SNP) rs2910164 in peripheral blood mononuclear cells (PBMCs). Interleukin (IL)-17, tumour necrosis factor (TNF)α, IL-1β, IL-8, IL-6 and monocyte chemoattractant protein (MCP)-1 in supernatants of PBMCs derived from healthy individuals of SNP rs2910164 CC (n=25–26), GC (n=31–34) and GG (n=22–23) genotypes were measured by ELISA. The horizontal line indicates the mean expression level within each group. *p<0.05; **p<0.01.
Discussion
In the present study, we investigated the association of five SNPs of hsa-miR-146a and the transcription factor Ets-1 with BD and VKH in a Chinese Han population. The results showed that individuals carrying the CC genotype of miR-146a rs2910164 had a lower risk of developing BD. Individuals having the CC genotype of rs2910164 showed a lower miR-146a expression as compared to rs2910164 GG individuals and a lower expression of IL-17, TNFα and IL-1β. We failed to show any association of the tested five SNPs of miR-146a and Ets-1 gene with VKH disease.
The choice of SNPs of these two genes as candidates was principally based on earlier studies.19 ,22–24 A total of 12 variants of the miR-146a gene were identified by Luo et al.13 Of the 12, 9 were already reported in the dbSNP database Build 130. Five of these nine variants (rs2910164, rs73318382, rs57095329, rs6864584 and rs17057381) had a minor allele frequency of >1% and the other four variants were monozygous. SNPs rs57095329 and rs73318382 are in strong linkage disequilibrium (LD; r2=0.81) with each other and the former has been shown to be highly associated with SLE in a Chinese Han population,13 and was therefore included in our study. With regard to SNP rs17057381, only one GG genotype has been reported in a representative Asian population (Han Chinese in Beijing, CHB) (http://www.ncbi.nlm.nih.gov/projects/SNP/snp_ref.cgi?rs=17057381), although the frequency of C allele was reported to be 0.016 in another Chinese population.13 A total of 900 individuals (300 normal controls, 300 patients with BD and 300 patients with VKH) was therefore tested to examine whether there were variants in this SNP rs17057381. Our result only showed the GG genotype in this population (data not shown), which is consistent with the HapMap database and therefore this SNP was not included in this study. As Ets-1 may strongly affect miR-146a promoter activity and directly regulate miR-146a expression,16 ,17 we also examined whether its variants influence the susceptibility to BD. Five SNPs of Ets-1 gene were investigated with SLE, and only rs1128334 and rs10893872, which located in the 3′-UTR of the this gene, were identified to be linked with this disease. As SLE and BD may share a gene background in common,25–29 we therefore included these two SNPs in this study.
To the best of our knowledge, the possible association of miR-146a and Ets-1 gene polymorphisms with BD and VKH disease have not yet been reported. This is the first study to identify an association of SNP rs2910164 of miR-146a with autoinflammatory disease. Our study showed a significantly decreased frequency of the CC genotype and C allele of this SNP in patients with BD, suggesting that this miR may be a susceptibility factor to this disease. A similar association has been reported in papillary thyroid carcinoma in European Caucasians22 and oesophageal squamous cell carcinoma in Chinese Han.30 These results suggest that the rs2910164 CC genotype and C allele are common predisposing factors for these diseases despite the fact that the pathogenic mechanisms underlying cancer and BD are totally different.
The aforementioned results identified a strong association between rs2910164 in the pre-miR-146a gene with BD. A further study was designed to investigate whether this SNP influenced miR-146a expression and the production of cytokines importantly involved in BD development. The disease group is not homogenous due the influence of various proinflammatory processes and the fact that the patients are treated with immunosuppressive drugs. We expect that these factors would influence gene expression and cytokine production and we therefore only investigated the miR-146a expression and cytokine production in healthy individuals with different genotypes of SNP rs2910164 in the miR-146a gene. Our study showed that the rs2910164 GG genotype could upregulate mature miR-146a expression. This finding is consistent with findings reported in papillary thyroid carcinoma in European Caucasians.22 The results also revealed that the IL-17, TNFα and IL-1β levels from individuals carrying the rs2910164 CC genotype were significantly decreased as compared to the other two genotypes. The lower IL-17, TNFα and IL-1β response in the CC genotype would fit in with the protection against disease in the patients with this genotype. It is worthwhile to point out that there was a somewhat high variation in the release of the tested cytokines by PBMCs stimulated with LPS. This is probably due to the fact that other genes may also play a role in cytokine expression. A recent in vitro study showed that miR-146a was involved in LPS-induced crosstolerance in a human macrophage cell line and showed that miR-146a operated as a negative regulatory feedback mechanism to prevent an inflammatory reaction.31 These data are not in agreement with our study, where we observed protection against BD with a genotype associated with lower miR-146a expression. The reasons for this discrepancy are unclear but could be due to the use of different types of cells (macrophage cell lines versus freshly isolated PBMCs). Our results are, by and large, consistent with those reported in patients with rheumatoid arthritis, in which upregulated miR-146a expression was found to be associated with increased IL-17 expression in PBMCs.32 ,33 The findings presented here are also not in agreement with earlier studies in mice where miR-146a was shown to act as a negative regulator possibly via negative regulation of the Toll-like receptor (TLR) signalling pathway by targeting TNF receptor associated factor (TRAF)634 and via increasing forkhead box P3 (FoxP3) expression in regulatory cells.35 Although the reasons for these controversies are not clear, differences between humans and mice might be one of the explanations. It is worthwhile to point out that a dose–response relationship was only noted in IL-17 and TNFα expression, but not present in IL-1β expression. The reasons as to why there is no dose–response relationship in IL-1β expression need to be clarified in future studies. Unexpectedly, we found an increased production of IL-8 in the CC genotype, which seems to be paradoxical to the results for IL-17, TNFα and IL-1β expression. A similar result was also reported in a study on ulcerative colitis in which the expression of IL-8 was inconsistent with that of IL-17 and IL-1β.36 Collectively, these results suggest that this SNP functionally affects the miR-146a expression level and also influences proinflammatory cytokine production.
Contrary to the result in patients with BD, we did not observe an association of these five SNPs with susceptibility to VKH. The discrepancy in the association of miR-146a polymorphisms between BD and VKH disease disclosed in the present study may be partially due to the fact that these two uveitis entities are mediated by different immunological and inflammatory pathways. BD, for instance, is now seen as an autoinflammatory disease, whereas VKH is considered an autoimmune disease directed against melanocytes.1 ,37
There are a number of limitations to our study. Our study identified an association of miR-146a polymorphisms with BD in a predominantly male38 ,39 Han Chinese population. Although this result was from gender adjustment, it should be validated in a gender-matched population. As replication in an independent set of samples is an important requirement for all association studies, the association of SNP rs2910164 in the miR-146 gene with BD needs further confirmation. BD involves multiple organs and our patients, recruited from a department of ophthalmology, all had uveitis and may therefore represent a subpopulation of this disease. The susceptible SNP identified in our study may only be associated with uveitis in BD and further studies are needed to investigate the association of miR-146a with BD in patients from other medical departments, such as stomatology and dermatology. In addition, although we failed to find any association of SNP rs57095329, rs6864584, rs1128334, rs10893872 with BD and VKH disease, our study does not rule out the possibility that other SNPs of Ets-1 are associated with both diseases. Further studies are needed to clarify this issue.
In conclusion, our study identified a strong association of rs2910164 in the pre-miR-146a gene with BD. Furthermore, our study suggested that it may affect the genetic predisposition to this disease possibly through modulating expression of mature miR-146a and influencing the control of proinflammatory cytokine production.
Acknowledgments
The authors would like to thank all donors enrolled in the present study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online table
Footnotes
-
Handling editor Tore K Kvien
-
Contributors QZ designed and performed the experiment, performed data analysis and wrote the manuscript. SH was involved in data analysis and writing of the manuscript. PY designed the experiment and was involved in writing the manuscript. All of the authors have given permission to be named.
-
Funding This work was supported by Natural Science Foundation Major International (Regional) Joint Research Project (30910103912), National Basic Research Program of China (973 Program) (2011CB510200), Key Project of Natural Science Foundation (81130019), National Natural Science Foundation Project (30973242), Research Fund for the Doctoral Program of Higher Education of China (20115503110002), Chongqing Key Laboratory of Ophthalmology (CSTC, 2008CA5003), Key Project of Health Bureau of Chongqing (2012-1-003), Key Project of Health Bureau of Chongqing (2012-1-003) and Fund for PAR-EU Scholars Program.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval This study was conducted with the approval of the Ethical Committee of Chongqing Medical University.
-
Provenance and peer review Not commissioned; externally peer reviewed.