Article Text

Abstract
Background Progression of joint destruction in rheumatoid arthritis (RA) is partly heritable; knowledge of genetic factors may increase our understanding of the mechanisms underlying joint destruction. The activity of the Wnt/β-catenin pathway influences osteoblast differentiation. Dickkopf-1 (Dkk-1) and sclerostin (Sost) are negative regulators and lipoprotein receptor-related protein-5 (LRP-5) and Kremen-1 are transmembrane receptors involved in this pathway.
Objective To study variants in the genes encoding these proteins in relation to progression of joint destruction.
Methods 1418 patients with RA of four cohorts with 4885 sets of hands and feet x-rays were studied. Explorative analyses were performed on 600 patients with RA from Leiden on single nucleotide polymorphisms (SNPs) tagging Dkk-1, Sost, Kremen-1 and LRP-5. SNPs significantly associating with joint damage progression were subsequently genotyped in cohorts from Groningen (NL), Sheffield (UK) and Lund (Sweden). Data were summarised in meta-analyses. Serum levels of functional Dkk-1 and sclerostin were measured and studied in relation to genotypes.
Results In the first cohort, six Dkk-1, three Sost, one Kremen-1 and 10 LRP-5 SNPs were significantly associated with radiological progression of joint destruction. Three Dkk-1 SNPs were associated significantly with progression of joint damage in the meta-analysis, also after correction for multiple testing (rs1896368, rs1896367 and rs1528873). Two Sost SNPs tended to significance (rs4792909 and rs6503475, p=0.07 after false discovery rate correction). Gene–gene interactions between SNPs on Dkk-1 and Sost were seen. Serum levels of Dkk-1 were significantly correlated with the genotypes in rs1896368 (p=0.02).
Conclusions Patients with RA carrying risk alleles of genetic variants in Dkk-1 have higher serum levels of functional Dkk-1 and more progressive joint destruction over time.
- Rheumatoid Arthritis
- Gene Polymorphism
- Bone Mineral Density
Statistics from Altmetric.com
Introduction
Persistent inflammation and destruction of joints are hallmarks of rheumatoid arthritis (RA). The natural course of the disease is highly variable between patients. Disease severity is often measured by progression of joint destruction. Known risk factors mainly relate to inflammation and autoantibodies and are estimated to explain 32% of the variance in joint destruction.1 Recently, the heritability of the rate of joint destruction was estimated to be 58%.2 Identifying individual genetic risk factors may increase our understanding of the mechanisms underlying differences in severity of joint destruction.
The Wnt/β-catenin pathway is central to bone development and homoeostasis. The original descriptions that causal mutations in low-density lipoprotein receptor-related protein-5 (LRP-5) or genetic defects in sclerostin are associated with syndromes characterised by high bone mass (high bone mass syndrome and sclerostosis, respectively) raised interest in this signalling pathway, and in recent years understanding of the Wnt/β-catenin pathway has progressed considerably. This pathway involves binding of Wnt proteins to a co-receptor complex, which includes LRP-5 or LRP-6 and a member of the frizzled family of proteins. This binding leads to a signalling cascade that results in intracellular release of β-catenin to the cytoplasm; ultimately, osteoblast differentiation is stimulated.3 ,4 Wnt signalling is regulated by several proteins, among which Dickkopf-1 (Dkk-1) and sclerostin (Sost). Sost binds to LRP-5 and prevents Wnt binding. Dkk-1 can bind to LRP-5 and the cell-surface co-receptor Kremen-1; the ternary complex is then internalised and degraded. Hence Dkk-1 and Sost both negatively regulate the Wnt signal.4
Aberrant regulation of the Wnt/β-catenin signalling pathway has been implicated in different types of cancer, cardiovascular and haematological diseases. Circulating levels of Dkk-1 have also been reported to be raised in RA5 ,6 and have been related to radiographic joint damage in RA.7–9 Diarra et al recently demonstrated that tumour necrosis factor α is a key inducer of Dkk-1 and that inhibiting Dkk-1 in a mouse model leads to reversal of bone destruction. Moreover, the expression of Sost seems to be positively correlated with Dkk-1 levels.10 Based on the known functions of Dkk-1, Sost, LRP-5 and Kremen-1 and the recent observation that a considerable part of the variance in joint destruction in RA is explained by genetic factors, we hypothesised that genetic variants in the genes encoding these four proteins that play a crucial role in the Wnt/β-catenin pathway are associated with progression of joint destruction in RA. We performed a multi-cohort candidate gene study to evaluate this.
Patients and methods
Study population
Four cohorts of adult European patients with RA were studied. RA was defined according to the 1987 American College of Rheumatology criteria except for the Lund cohort where the 1958 criteria were used. x-Rays of both hands and feet were available in all cohorts (table 1).
Characteristics of each cohort
Leiden early arthritis clinic (Leiden EAC)
Six hundred patients with early RA included in the Leiden early arthritis clinic (Leiden EAC) between 1993 and 2006 were studied.1 x-Rays were taken at baseline and yearly thereafter during 7 years of follow-up (in total 2846 sets of hands and feet x-rays). x-Rays were chronologically scored by one reader using the Sharp–van der Heijde score (SHS).11 The intraclass correlation coefficient (ICC) among the readers was 0.91. Treatment strategies were different in three treatment periods: 1993–5, initial treatment with non-steroidal anti-inflammmatory drugs, 1996–8, initial treatment with chloroquine or salazopyrine and 1999–2006, prompt treatment with methotrexate or salazopyrine.
Groningen
The second cohort involved 275 patients with RA from the northern part of the Netherlands who were diagnosed between 1945 and 2001. Over at most 14 years of follow-up the mean number of x-ray sets (hands and feet) per patient was 3.1. The x-rays were scored chronologically using SHS (ICC=0.96). Patients included in the 1990s were given (mild) disease-modifying antirheumatic drug (DMARD) treatment in contrast to patients included before 1990.
Sheffield
Three hundred and ninety-six patients with RA diagnosed in 1938–2003 with x-rays available were recruited from the rheumatology department of the Royal Hallamshire Hospital in Sheffield, UK in 1999–2006.12 Patients with RA were assessed once at a mean (±SD) disease duration of 15±11 years. Hands and feet x-rays were scored by one reader using a modification of Larsen's score.13 Ten per cent of films were scored twice to quantify the intraobserver variation by a weighted κ score which was 0.83.12
Lund
This cohort comprised 183 Swedish patients with early RA who were prospectively followed up for 5 years, of whom 147 had x-rays and DNA available.14 ,15 Patients were recruited during 1985–9. x-Rays of hands and feet were taken at baseline and annually for 5 years, resulting in a total of 781 sets of x-rays. x-Rays were scored chronologically according to Larsen by one of two readers (ICC between readers 0.94).16 In the first years DMARD treatment was uncommon and only half of the patients used any DMARD at 5-years’ follow-up, most commonly chloroquine, D-penicillamine, sodium aurothiomalate and auranofin.15
All patients gave informed consent. Approval was obtained from the local medical ethical committee of each cohort.
Single nucleotide polymorphism (SNP) selection and genotyping
Pairwise tagging SNPs (n=77) (±300 kb) were selected from the CEPH/CEU hapmap dataset (phase 2, release 21, NCBI build 35) using Haploview software (MAF>0.05, pairwise r2>0.8). For Dkk-1, Sost and Kremen, no amino acid-changing SNPs were known. For LRP-5, rs3736228 is amino acid changing and has been associated with bone mineral density in various studies.17 ,18 This SNP was therefore forced into the selection. SNPs that were significantly associated with joint destruction in phase 1 were genotyped in the other three cohorts. Genotyping was done with an Illumina GoldenGate assay in phase 1 and Sequenom iPLEX in phase 2, as described previously.19 The overall error rate was <1%; the success rates were >95%.
Measuring serum levels of Dkk-1 and plasma levels of sost
Levels of functional Dkk-1 in the baseline serum samples of 80 patients with RA from the Leiden EAC were measured using ELISA (BI-20412, Biomedica, Austria). Levels of sclerostin were determined in baseline plasma of a different set of 80 patients using ELISA (BI-20492, Biomedica, Austria). For both ELISAs, patients were selected based on the genotype in the most significantly associated SNP.
Statistical analysis
In all cohorts, the radiological scores were log transformed to obtain normal distributions (see online supplementary figures S1 and S2). Associations between genotypes and radiographic joint destruction were analysed in two phases. First, an explorative analysis was performed; tagged SNPs were tested additively and recessively. Since this phase was explorative, no correction for multiple testing was applied at this stage and SNPs with a p value <0.05 were studied in phase 2 using an additive or recessive model, based on the findings in phase 1.
Phase 1
A multivariate normal regression model for longitudinal data was used with the radiological score as a response variable. This method analyses all repeated measurements at once and takes advantage of the correlation between these measurements. To test for an association with the rate of joint destruction, an analysis with the SNP and its interaction with time in the model was conducted. The effect of time in the interaction term was linear. The resulting coefficient (β) indicates how many fold the progression of joint destruction increased per year per minor allele when an additive effect was tested, and how many fold the progression of joint destruction increased per year in the presence of two minor alleles when a recessive model was tested. Adjustment variables (age, gender, treatment strategies) were entered based on their univariate association with joint destruction.
Phase 2
For the analyses in Groningen and Lund a multivariate normal regression analysis was used, similar to that applied in the Leiden EAC. Analyses in the Groningen cohort were adjusted for age and a proxy for DMARD treatment (inclusion </>1990). The analyses at Lund were adjusted for age only, since gender and treatment were not associated with joint destruction in this dataset. In the Sheffield cohort, each patient had x-ray examinations at one time point only. To make scores comparable to those of the other cohorts, the estimated yearly progression rate was calculated by dividing the total SHS by the number of disease years at the time of the x-ray examination.20 The SNP association was tested in a linear regression analysis with the log-transformed estimated yearly progression rate as outcome. No adjustments were applied as none of the tested variables were associated with joint destruction. Also here, the resulting estimate reflects how many fold the rate of joint destruction increases per year in the presence of a risk variant. Analyses were done using SPSS V.17.0.
Meta-analysis
All three cohorts studied in phase 2 contained fewer patients and fewer x-ray examinations than the initiation cohort. Consequently, the power was insufficient to replicate findings in each cohort individually, as well as in the three replication cohorts together. Because of differences in study designs, the separate cohorts could not be combined directly in one analysis. Therefore, it was decided to test the SNPs in each cohort separately taking advantage of the specific dataset characteristics, and subsequently, to perform a fixed-effects meta-analysis with inverse variance weighting.21 Since the parameters in all datasets reflect the relative increase in the rate of joint destruction per year, pooling of the estimates of the individual cohorts in a meta-analysis was allowed.22 Stata V.10.1 was used.
Multiple testing
To reduce the chance of false-positive findings, multiple testing correction using the Benjamini and Hochberg false discovery rate (FDR) was done in phase 2. p Values <0.05 after correction for multiple testing were considered significant.23
Haplotype analysis
Haplotypes of Dkk-1 were studied. Blocks were defined by Gabriel's method.24 Haplotypes were assigned to each individual using PLINK 1.06, requiring a probability >0.8. Analyses of the haplotypes were performed with methods similar to those used for the analyses of the individual SNPs by testing the presence of a haplotype compared with the absence of the haplotype.
Gene–gene interactions
Multiplicative interactions between the significantly associating SNPs on Dkk-1 and Sost were tested by including the multiplication of the Dkk-1 SNP, Sost SNP and time into the additive model for the Leiden patients with RA. Also here, correction for multiple testing was performed.
Results
A complete overview of the results of all SNPs studied in phase 1 is presented in online supplementary table S1.
Dkk-1
Phase 1
Dkk-1 was tagged by eight SNPs. Six of these associated significantly with the rate of joint destruction in the Leiden EAC in an additive model: rs1896368, rs10762715, rs1528873, rs1441124, rs1194750 and rs1896367. Patients with one minor allele had respectively a 1.02 (95% CI 1.01 to 1.04), 1.03 (1.01 to 1.05), 1.02 (1.00 to 1.04), 1.03 (1.01 to 1.05), 1.04 (1.01 to 1.07) and 0.98 (0.97 to 0.99) fold progression rate per year per minor allele (figure 1). An estimate of 1.03 a year equals a 23% (1.037) higher rate of joint destruction over 7 years, whereas a β of 0.98 a year results in a 14% lower rate of joint destruction over 7 years. Haplotype analyses were performed but did not yield additional information to the SNP analyses.
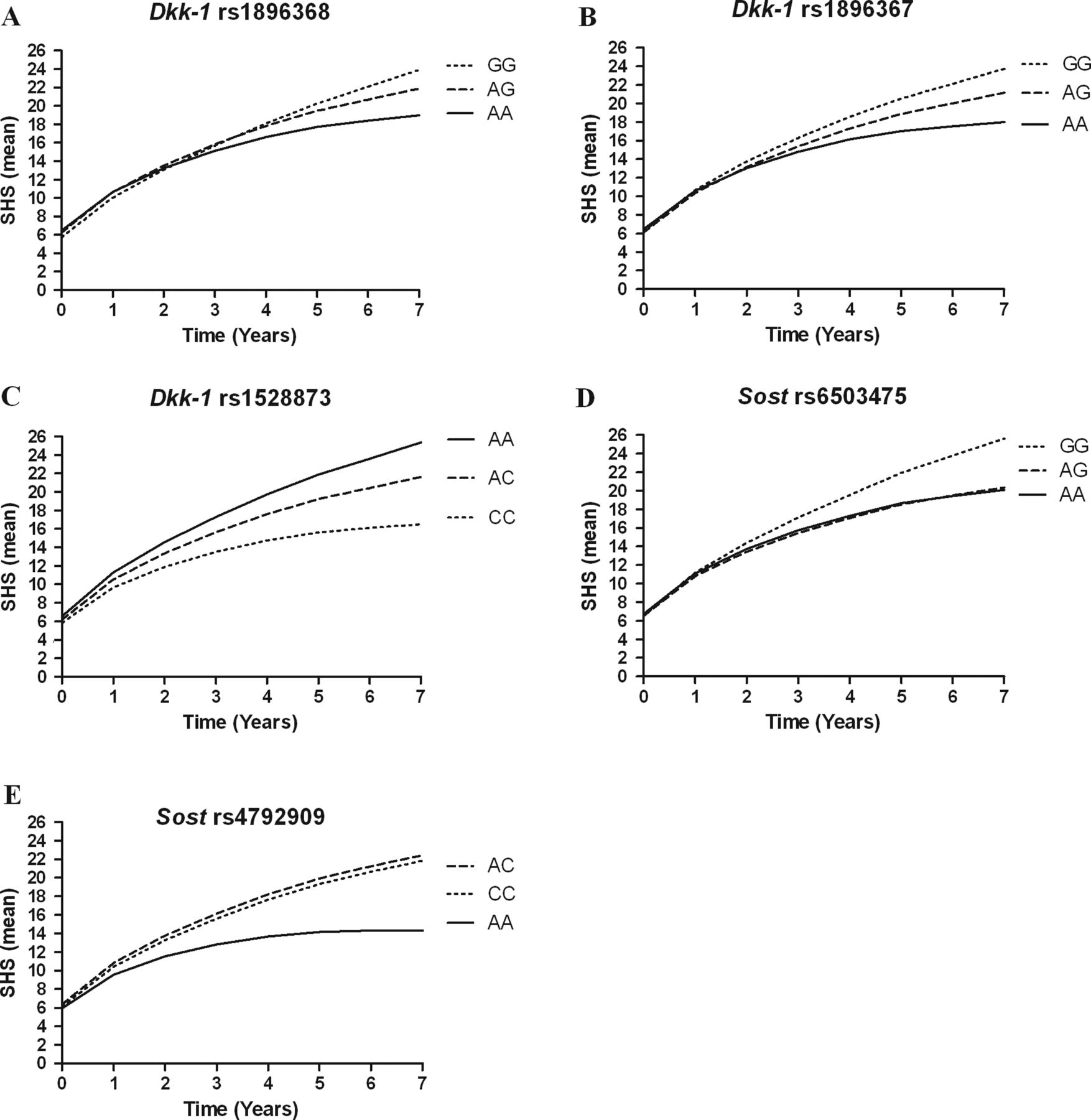
Radiographic joint damage progression in Leiden patients with rheumatoid arthritis. Depicted are the predicted (by the multivariate normal regression model) Sharp–van der Heijde scores (SHS, mean) during 7 years of follow-up of patients with different genotypes in phase 1 (Leiden early arthritis clinic) for the three single nucleotide polymorphisms (SNPs) in Dkk-1 that remained significant after correction for multiple testing and the two SNPs in Sost that were significant in the meta-analysis. All Dkk-1 SNPs were analysed in an additive model, all Sost SNPs in a recessive model.
Phase 2
The significantly associating SNPs were typed in the cohorts from Sheffield, Groningen and Lund. As the numbers of x-ray examinations of the individual cohorts were lower than those of phase 1, it was not expected that significant results would be achieved in each cohort. We therefore performed a meta-analysis summarising the results of the four cohorts in phase 2. All six SNPs associated significantly with radiographic joint damage progression (see table 2, figure 2). The strongest association was found for rs1896368 (p<0.001). After correction for multiple testing, three SNPs remained significant (rs1896368, rs1896367 and rs1528873, table 2).
Results of inverse variance meta-analyses on the four cohorts
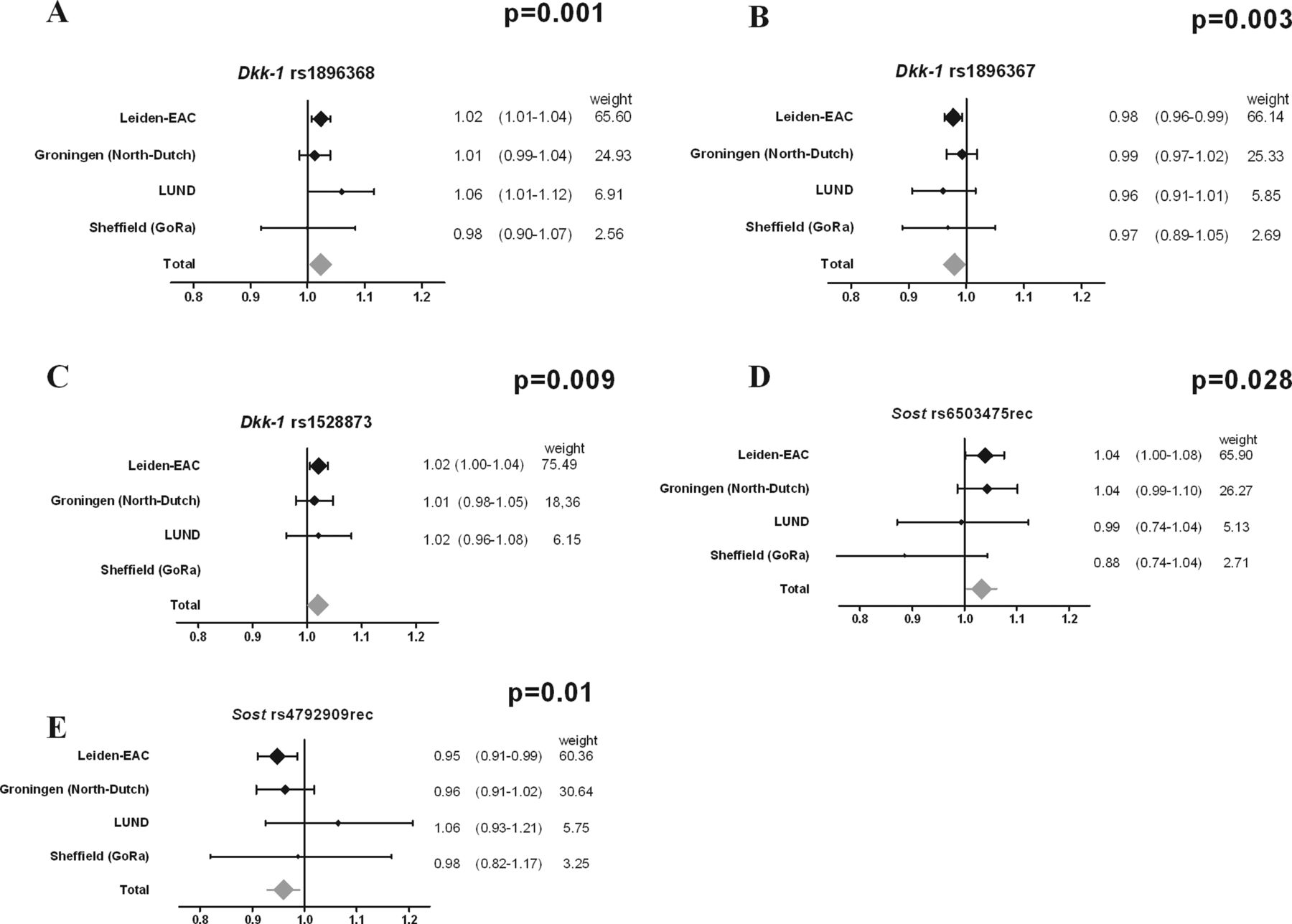
Results of the meta-analysis. Depicted are the results of the analyses of the individual cohorts and the results of the meta-analysis performed on the SNPs that were significant (after correction for multiple testing) in phase 2. The p values in the graphs are false discovery rate corrected for multiple testing. The meta-analyses are based on a fixed-effects model, which is applied to genetic studies to test whether there is a statistically significant effect; generalisability of the effect is of less importance. As a result of this choice, this method is less suitable for estimating the effect size overall. Therefore, the estimated effect of the meta-analysis is depicted in grey and without numbers. After false discovery rate correction, rs1896368, rs1896367 and rs1528873 in Dkk-1 were significant. The I2 for these SNPs were all 0.0%, indicating little heterogeneity between the cohorts. The effect sizes of these three SNPs in the meta-analyses were 1.02 (95% CI 1.01 to 1.04), 0.98 (95% CI 0.97 to 0.99) and 1.02 (95% CI 1.00 to 1.04), respectively. Rs1528873 could not be typed in the Sheffield patients. EAC, early arthritis clinic; SNP, single nucleotide polymorphism.
Sost
Phase 1
Sost was tagged by nine SNPs. Three of these associated significantly with the rate of joint destruction in phase 1 using a recessive model. Patients carrying two minor alleles of rs4792909 had a 0.95 (95% CI 0.91 to 0.99) fold higher progression rate a year than patients carrying one or no minor alleles. When patients carried two minor alleles of rs6503475, they had a 1.04 (1.00 to 1.08) fold higher progression rate per year. In addition, patients with one or two minor alleles of rs1260054 had a 0.95 (0.92 to 0.99) fold higher progression rate than patients carrying one or no minor allele.
Phase 2
These three SNPs were studied in the three other cohorts using a recessive model. In the meta-analysis two SNPs were significantly associated with joint damage progression: rs4792909 and rs6503475 (table 2). However, after FDR correction for multiple testing, only a trend towards significance remained (p=0.07 for both SNPs).
LRP-5
LRP-5 was tagged by 44 SNPs. Ten associated significantly with radiographic progression in phase 1, among which was rs3736228 (β=0.97 (0.95–0.99). Owing to technical difficulties, only one (rs3736228) could be typed in the three other cohorts. This SNP, which is amino acid changing, showed no significant association with radiographic joint damage progression in phase 2 assuming an additive model (table 2).
Kremen-1
Kremen-1 was tagged by 16 SNPs. Three associated significantly with joint destruction: rs1322774 (β=0.98 (0.96–0.99)). After typing in the three additional cohorts and analysing the combined results in a meta-analysis, rs1322774 showed a borderline significant effect (p=0.05), which was lost after correction for multiple testing (table 2).
Interactions between Dkk-1 and Sost
Subsequently it was analysed whether gene–gene interactions between components of the Wnt/β-catenin pathway exist. Several SNPs in Dkk-1 (n=6) and Sost (n=2) associated with progression of joint destruction (phase 2 before multiple testing correction). The 12 possible Dkk-1–Sost combinations were analysed for multiplicative interactions in Leiden patients with RA. Subsequently, the resulting p values were corrected for performing 12 tests, by both FDR and Bonferroni correction (see online supplementary table S2). After FDR correction, three combinations of SNPs were significant: rs6503475 (Sost) and rs10762715 (Dkk-1) (corrected p=0.05), between rs4792909 (Sost) and rs10762715 (Dkk-1) (corrected p=0.03), and between rs6503475 (Sost) and rs1528873 (Dkk-1) (corrected p=4.52×10−9). The SHS data in the presence of a combination of risk alleles of these SNPs are presented in figure 3.

Radiological destruction data of single nucleotide polymorphisms (SNPs) on Dkk-1 and Sost in which multiplicative interaction was seen. Depicted are the mean Sharp–van der Heijde scores (SHS) during 7 years of follow-up of patients with different genotypes of (A) Sost rs6503475 and Dkk-1 rs10762715, for patients with no risk alleles on one of these SNPs (n=69), one risk allele (n=158), two risk alleles (n=157), three risk alleles (n=53) and twice the risk allele for both SNPs (in total four risk alleles) (n=9); (B) Sost rs4792909 and Dkk-1 rs10762715, for patients with no risk alleles on one of these SNPs (n=108), one risk allele (n=263), two risk alleles (n=163), three risk alleles (n=49) and twice the risk allele for both SNPs (in total four risk alleles) (n=9); (C) Sost rs6503475 and Dkk-1 rs1528873, for patients with no risk alleles on one of these SNPs (n=40), one risk allele (n=114), two risk alleles (n=170), three risk alleles (n=104) and twice the risk allele for both SNPs (in total four risk alleles) (n=18).
Correlations between genotypes and levels of Dkk-1 and sclerostin
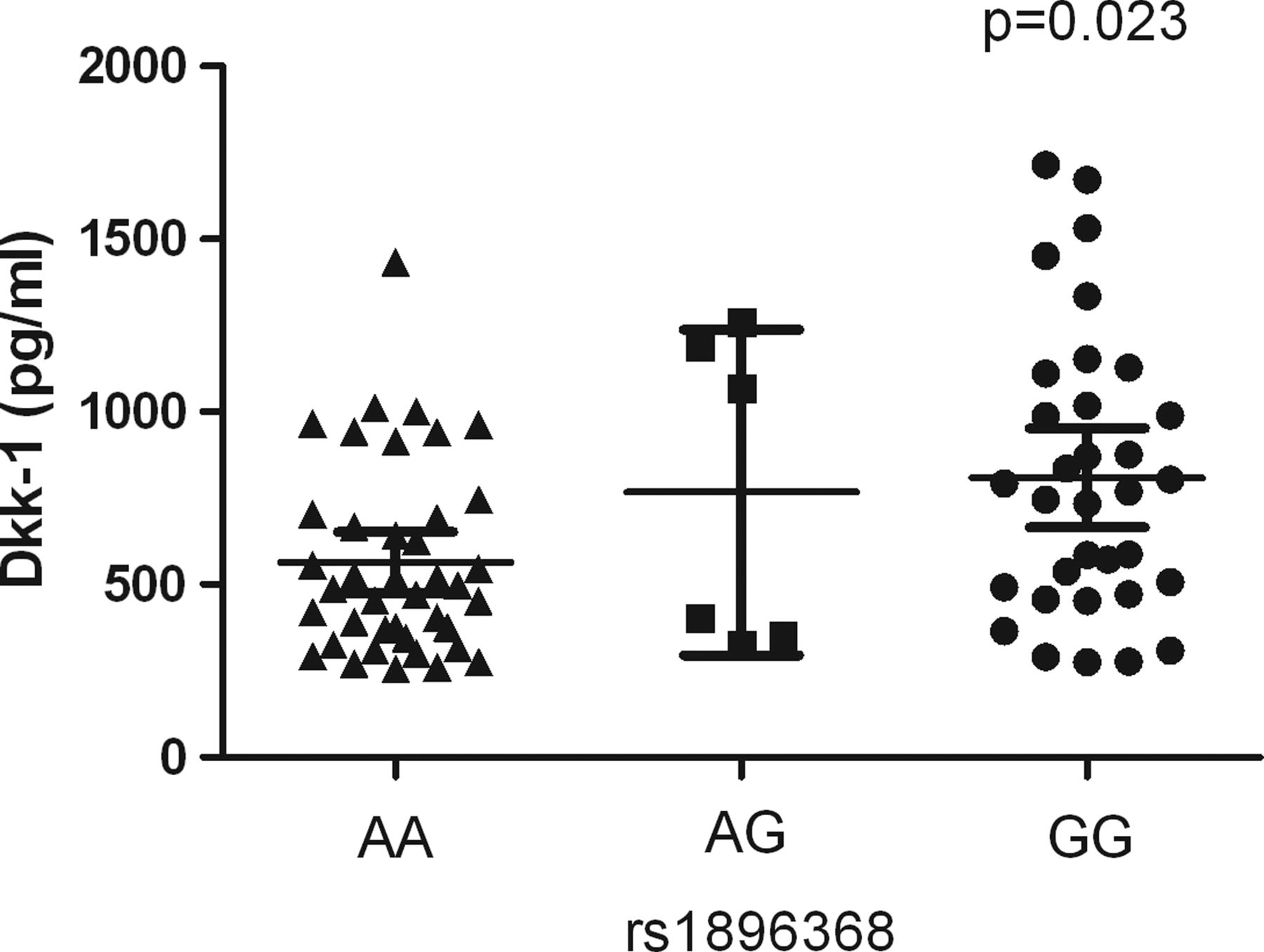
Next we studied whether the genetic variants in the best associating SNP on Dkk-1 (rs1896368) and on Sost (rs4792909 and rs6503475 were analysed both because of comparable effect sizes) associated with serum levels of functional Dkk-1 and plasma levels of sclerostin, respectively. Plasma levels of sclerostin were not significantly associated with rs4792909 and rs6503475 genotypes (data not shown). However, patients carrying the rs1896368-GG genotype in Dkk-1, which is associated with higher radiographic progression rates, had significantly higher serum levels of functional Dkk-1 than patients carrying the other genotypes (p=0.023, figure 4).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Levels of Dkk-1 in relation to various genotypes. Depicted are the levels of functional Dkk-1 in the serum samples of 80 patients and the corresponding genotypes in rs1896368 in Leiden patients with rheumatoid arthritis who were selected according to their rs1896368 genotypes.
Discussion
We evaluated four genes encoding proteins that are critically involved in the Wnt/β-catenin signalling pathway in relation to the severity of progression of joint destruction in RA in 4885 x-ray sets of hands and feet radiographs of 1418 patients with RA. The major finding is that patients with RA carrying the risk alleles of several genetic variants in Dkk-1 have higher serum levels of functional Dkk-1 and more progressive joint destruction over time. These findings are in line with recent observations that Dkk-1 serum levels are associated with joint destruction.7 ,8 Together these data illustrate the relevance of Dkk-1 to progression of joint destruction in RA. Based on these data it may also be hypothesised that blocking the binding of Dkk-1 to LRP-5 or LRP-6 may lead to new therapeutic strategies to protect patients against progression of joint damage.
Two genetic variants in Sost were suggestively associated with progression of joint destruction, but after correction for multiple testing only a tendency towards significance remained. Furthermore, the results for Sost were obtained using a recessive model and a small number of patients homozygous for the minor allele. Altogether, definite conclusions about Sost in relation to joint damage cannot be drawn. Nonetheless, rs1513670, a full proxy of rs6503475 in Sost, was recently associated with bone mineral density in a genome-wide study, raising interest in this particular variant in relation to bone loss.25 In order to search for functional consequences of carrying the risk genotypes of this SNP, we also studied plasma Sost levels; no association was detected. This does not preclude the possibility that carrying the risk genotypes is biologically relevant as Sost is mainly expressed by osteocytes, and local expression in bone may not be measurable in the systemic circulation.
Four Wnt signalling pathways have been described.26 The Wnt/β-catenin signalling pathway, in particular, is known for its pivotal role in bone homoeostasis. We therefore performed this candidate gene study, evaluating tagging SNPs of four relevant genes belonging to the Wnt signaling pathway. An advantage of analysing several factors in one pathway is the ability to look for interactions. Hypothetically, carrying two different risk factors may have an enhancing effect that is greater than would be expected from a combination of the two. Additionally, a cross-talk between Dkk-1 and Sost has recently been observed in vitro.10 We studied interactions between the Dkk-1 and Sost SNPs and observed significant multiplicative interactions. The most compelling effect on joint damage was present in patients carrying four risk alleles, which was only a small group. To evaluate whether the observed interaction was totally driven by these patients, we repeated the interaction analysis for rs1528873 and rs6503475 excluding the patients carrying four risk alleles. A significant effect still remained (p=0.021), indicating that the observed finding was not entirely based on this small group of patients. Since the numbers of patients in the replication cohorts were smaller than that in phase 1 and data could not be easily combined, interactions were not analysed in the cohorts of phase 2.
This study has limitations. The most relevant limitation is that the cohorts used in phase 2 contained fewer radiographs than the initial cohort. Hence the replication cohorts were insufficiently powered to achieve independent replications. However, since large prospective cohorts with high-quality phenotypic data are scarce, it was decided to combine the data in a meta-analysis.
In this study genetic variants in the Dkk-1 gene were associated with increased joint destruction rates and with higher serum levels. The serological data are in line with findings of other studies where low serum levels were found in diseases characterised by bony depositions, such as ankylosing spondylitis and diffuse idiopathic skeletal hyperostosis.27 ,28 To evaluate whether the functional effects of carrying the Dkk-1 risk genotype also influenced the Dkk-1 serum levels, both SNP and serology data were included in one model with the radiographic progression rate as an outcome variable. Here, only the SNP effect was significant. This may be due to the fact that the 80 patients who were studied at serum level were selected according to their genotypes.
In conclusion, this study evaluated phenotypic and genetic data of four longitudinal cohorts and showed that patients with RA carrying risk alleles of genetic variants in Dkk-1 have higher serum levels of functional Dkk-1 and more progressive joint destruction over time. These data illustrate the relevance of Dkk-1 to progression of joint destruction in RA.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online figure 1
- Data supplement 2 - Online figure 2
- Data supplement 3 - Online table 1
- Data supplement 4 - Online table 2
Footnotes
-
Correction notice This article has been corrected since it was published Online First. The author name Sasha A Zhernakova has been amended to read A Zhernakova.
-
Contributors AGW, EL, TS, MKL, TWJH, REMT, EB and AHMH-M collected the patients’ data. DPCdR, RK, AK, NAD, ST, JJH-D, TWJH, REMT and AHMH-M carried out the statistical analyses. NGY and DLPB were responsible for the measurements of Dkk-1 and Sost. DPCdR and AHMH-M wrote the first version of the manuscript. All authors read and approved the manuscript.
-
Funding This research has been funded by The European Community Seventh Framework Program FP7-256 Health-F2-2008-223404 (Masterswitch). The work of RK and AK is supported by grants of the Dutch Arthritis Foundation (Reumafonds). The work of AHMvdH is supported by the Dutch organisation of Health research and development (Zon-MW).
-
Competing interests None.
-
Ethics approval Medical ethical committee's of the participating centres.
-
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Correction