Article Text

Abstract
Objectives To increase understanding of the aetiology and pathogenesis of rheumatoid arthritis (RA), genetic and environmental risk factors for RA subsets, defined by the presence or absence of different anticitrullinated protein/peptide antibodies (ACPAs) targeting citrullinated peptides from α-enolase, vimentin, fibrinogen and collagen type II, were investigated.
Methods 1985 patients with RA and 2252 matched controls from the EIRA case-control cohort were used in the study. Serum samples were assayed by ELISA for the presence of anticyclic citrullinated peptides (anti-CCP) antibodies and four different ACPA fine specificities. Cross-reactivity between ACPAs was examined by peptide absorption experiments. Genotyping was performed for HLA-DRB1 shared epitope (SE) alleles and the PTPN22 gene, while information regarding smoking was obtained by questionnaire. The association of genetic and environmental risk factors with different subsets of RA was calculated by logistic regression analysis.
Results Limited cross-reactivity was observed between different ACPA fine specificities. In total, 17 RA subsets could be identified based on their different ACPA fine specificity profiles. Large differences in association with genetic and environmental determinants were observed between subsets. The strongest association of HLA-DRB1 SE, PTPN22 and smoking was identified for the RA subset which was defined by the presence of antibodies to citrullinated α-enolase and vimentin.
Conclusion This study provides the most comprehensive picture to date of how HLA-DRB1 SE, PTPN22 and smoking are associated with the presence of specific ACPA reactivities rather than anti-CCP levels. The new data will form a basis for molecular studies aimed at understanding disease development in serologically distinct subsets of RA.
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) has for many years been considered a prototype for immune-mediated diseases owing to the strong genetic association with certain MHC class II alleles (ie, HLA-DRB1 shared epitope (SE)).1,–,3 More recently the MHC class II association in RA was shown to be confined to a subset of patients characterised by the presence of antibodies to synthetic cyclic citrullinated peptides (CCP).4 ,5 The concomitant recognition that the best known environmental risk factor for RA (ie, smoking) constitutes a risk only for the development of CCP-positive disease, together with the finding that smoking induces the exposure of citrullinated proteins in the lungs,4 ,6 ,7 suggests an aetiology for RA where HLA-DRB1 SE-restricted immune reactions to citrullinated epitopes trigger and drive disease pathogenesis.8 Subsequently, it has been shown that the CCP assays such as the most frequently used CCP2 ELISA is able to capture anticitrullinated protein/peptide antibodies (ACPAs) with a broad spectrum of fine specificities including naturally occurring citrullinated autoantigens.9 ,10
In order to increase our understanding of the aetiology and pathogenesis of RA it has therefore become important to define which ACPA fine specificities are associated with the different HLA-DRB1 SE alleles. Such studies have been performed for some ACPA fine specificities11,–,15 and differences within the CCP-positive population have been described. However, no comprehensive picture has emerged to date, partly because only single target antigens have been studied and partly due to small study populations and lack of information on appropriate controls. In the present study we have used the large and well-characterised Epidemiological Investigation of Rheumatoid Arthritis (EIRA) case-control cohort16 to investigate HLA-DRB1 SE in combination with PTPN22 polymorphism—the second most important genetic risk factor for CCP-positive RA17—and cigarette smoking in relation to the presence of antibodies targeting four well-defined citrullinated autoantigens (citrullinated α-enolase, -vimentin, -fibrinogen and -collagen type II).
Methods
Subjects
EIRA is an ongoing Swedish population-based case-control study which is described in detail elsewhere.16 In the present study we analysed 1985 incident cases of RA included in the EIRA study between May 1996 and May 2006 and 2252 controls randomly selected from the national population register to match RA cases in terms of age, gender and residential area. Cases were diagnosed with RA according to the 1987 American College of Rheumatology criteria.18 Cases and controls donated blood at the time of inclusion and completed a self-administered questionnaire relating to life style and environmental exposures.
ELISA
Anti-CCP antibody status was determined in serum from 1985 cases and 1210 controls using the CCP2 assay (Immunoscan CCPlus, Euro-Diagnostica, Malmö, Sweden) according to the manufacturer's instructions. The cut-off for positivity was 25 AU/ml. Antibody responses to citrullinated peptides from α-enolase (CEP-1), vimentin (Cit-vim), fibrinogen (Cit-fib) and collagen type II (Cit-C1) or the arginine-containing equivalents were assayed in serum samples from 1985 EIRA cases and 150 randomly selected EIRA controls using an in-house ELISAs, as previously described.11 ,19 The cut-off for positivity was 10 AU/ml for all ACPA fine specificities. A detailed description of the ELISA method is given in the online supplement.
Cross-reactivity assay
Ten serum samples with high anti-CCP levels (>800 AU/ml) that also displayed reactivity to CEP-1 (n=10), Cit-vim (n=8), Cit-fib (n=9) and Cit-C1 (n=8) were selected from EIRA for cross-reactivity experiments. Each serum (diluted 1:100) was incubated with each of the four peptides (CEP-1, Cit-vim, Cit-fib or Cit-C1) at 10 µg/ml or in the presence of buffer alone. Preabsorption was performed in liquid phase during constant agitation for 2 h at room temperature. Following centrifugation (15 min, 1000 g), the supernatants were transferred to the four peptide ELISA plates and assayed as described above. Inhibition was determined based on the differences in AU/ml values between the absorbing peptide and buffer alone.
Peptides
Freeze-dried synthetic peptides (>95% purity) corresponding to sequences from human α-enolase (amino acid 5–21), vimentin (amino acid 60–75) and fibrinogen (amino acid 36–52) in its ‘native’ form or with arginine to citrulline substitutions (Innovagen, Lund, Sweden), were dissolved at 10 mg/ml (Milli-Q water, 10% DMSO) and stored at −20°C before used. The homotrimeric triple helical collagen type II peptides C1 and Cit-C1 were synthesised as previously described.20 ,21 A detailed description of the peptide sequences is given in table 1 in the online supplement.
ORs for disease risk in different subsets of rheumatoid arthritis in carriers of HLA-DRB1 shared epitope (SE) compared with non-carriers
Genotyping
HLA-DRB1 subtyping was performed on DNA samples from 1961 RA cases and 1278 controls by sequence-specific primer PCR, as previously described.11 DRB1*01 (except DRB1*0103), DRB1*04 and DRB1*10 were classified as shared epitope (SE) alleles of HLA-DRB1. The protein tyrosine phosphatase gene (PTPN22 rs2476601) was genotyped on 1943 RA cases and 1284 controls, as previously described.17
Cigarette smoking
Information regarding cigarette smoking prior to the appearance of symptoms of arthritis in 1727 RA cases and the corresponding time points among 1913 controls were obtained by questionnaire. Individuals were categorised as ever smokers (current/past) or never smokers.16
Statistical analyses
To determine the association of HLA-DRB1 SE (alone or in combination with PTPN22 and/or smoking) with different subsets of RA, ORs with 95% CI were calculated using unconditional logistic regression analyses, with cases and controls that were negative for the risk factors as the referent category. All analyses were performed with adjustment for design variables (age, residential area and gender).
Interaction, defined by departure from additivity of effects,22 was evaluated between HLA-DRB1 SE and smoking and between HLA-DRB1 SE and PTPN22. The attributable proportion due to interaction (AP) was calculated together with 95% CI, as previously described.11 ,23 The AP between two interacting factors reflects the joint effect beyond the sum of the independent effects. All analyses were performed using SAS Version 9.1.
Differences in ACPA levels between different RA subsets or between absorbing peptides and buffer in the cross-reactivity experiment were examined using the Mann–Whitney U test for independent groups.
Results
Identification of different RA subsets based on ACPA fine specificity profiles
Sixty-three percent of RA cases in EIRA were anti-CCP antibody positive (35% anti-CEP-1 positive, 37% anti-Cit-vim positive, 28% anti-Cit-fib positive and 37% anti-Cit-C1 positive. Low-level antibody responses to the arginine-containing control peptides from α-enolase, vimentin and fibrinogen were detected in less than 2.4% of patients, while reactivity to the arginine-containing C1 peptide20 ,21 ,24 of collagen type II was detected in 9% (data not shown). The presence of different ACPA fine specificities overlapped to a large extent and, by analysing the different patterns, 17 serologically distinct subsets could be identified (figure 1A). Most of the patients who were positive for the specific ACPAs were confined to the CCP-positive population, although 18% of CCP-negative patients were positive for at least one ACPA fine specificity (figure 1B). Conversely, 8.7% of all patients were CCP-positive but negative for the four ACPA fine specificities.

Subsets of rheumatoid arthritis (RA) defined based on the presence (+) or absence (−) of four different anticitrullinated protein/peptide antibody (ACPA) fine specificities (α-enolase (CEP-1), vimentin (Cit-vim), fibrinogen (Cit-fib) and collagen type II (Cit-C1)) as detected by ELISA in (A) 1985 cases with RA and (B) 730 anticyclic citrullinated peptides (anti-CCP) negative RA cases. In (A) the subsets may comprise CCP positive and CCP negative RA cases, with percentages of anti-CCP positive cases shown in brackets.
In the CCP-negative population, ACPA-positive patients typically showed reactivity to only one citrullinated antigen where anti-Cit-C1 antibodies were dominant (9.45%). In the CCP-positive subset, on the other hand, the presence of two (16.6%), three (13.4%) or four (11%) ACPA fine specificities was common. ACPA fine specificity levels were generally lower in the CCP-negative population than in the CCP-positive population (data not shown). Moreover, there was a strong correlation between the number of ACPAs present and anti-CCP antibody levels, with a significant (p<0.0001) increase in anti-CCP antibody levels for every additional ACPA fine specificity (see figure 1 in online supplement).
ACPAs show limited cross-reactivity
In order to analyse the extent of cross-reactivity between antibodies reactive with the different citrullinated peptides, serum samples from 10 CCP-positive RA patients with antibodies to CEP-1 (n=10), Cit-vim (n=8), Cit-fib (n=9) and Cit-C1 (n=8) were preabsorbed against each of the four peptides before being retested for ACPA reactivities. All four ACPAs were efficiently absorbed by preincubation with its homologous peptide, while only limited cross-reactivity was observed between specificities. Anti-CEP-1 IgG showed a low degree of cross-reactivity with Cit-fib in some serum samples (p=0.043) but no cross-reactivity with Cit-vim or Cit-C1 (figure 2A). Anti-Cit-vim IgG showed cross-reactivity with Cit-fib in two serum samples (figure 2B). Anti-Cit-fib IgG showed some degree of cross-reactivity with CEP-1 in some serum samples (p=0.05, figure 2C) and anti-Cit-C1 antibodies showed cross-reactivity mainly with Cit-fib in approximately half of the serum samples (p<0.001, figure 2D).
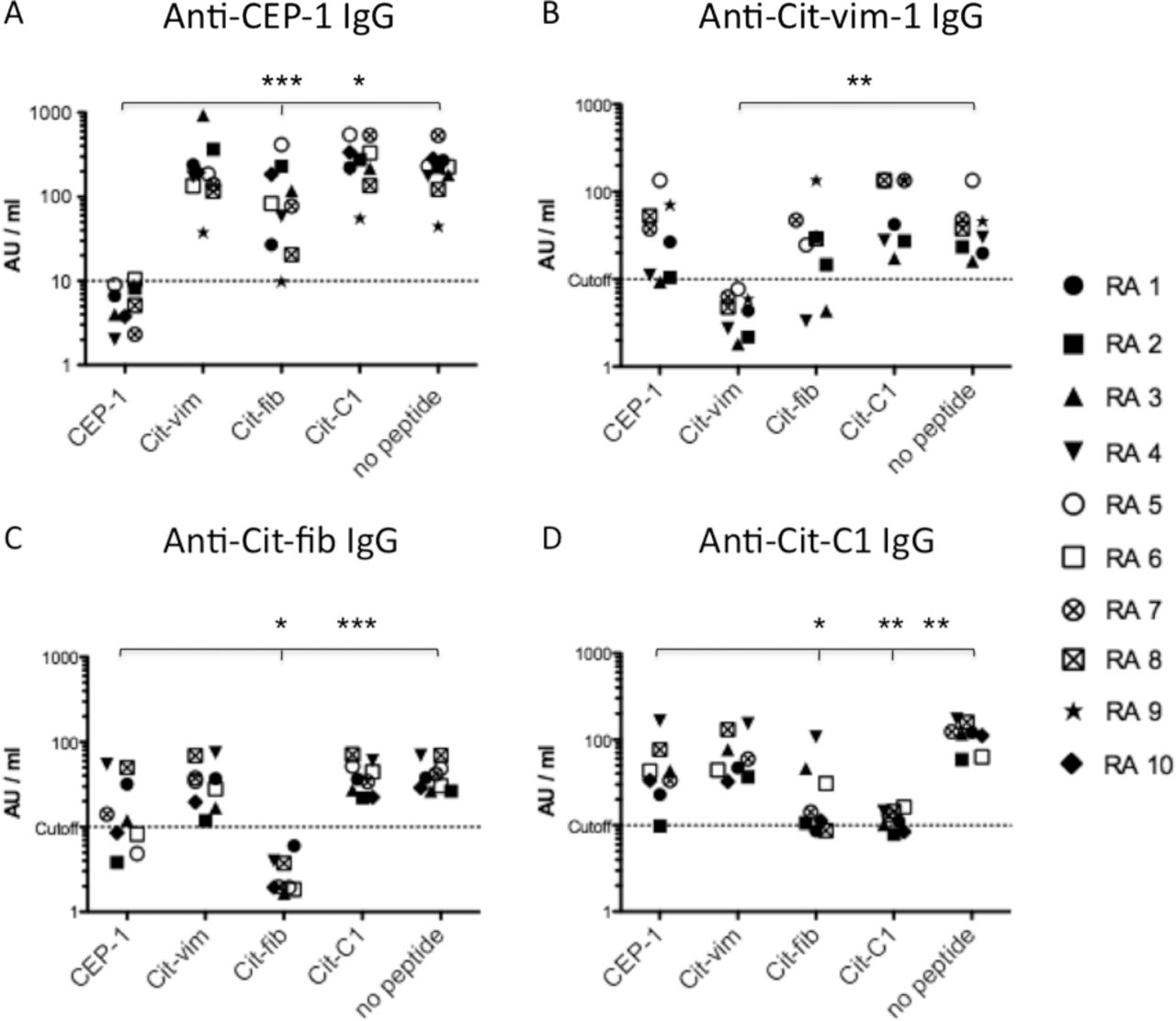
Cross-reactivity between different anticitrullinated protein/peptide antibody (ACPA) fine specificities. Antibody responses to citrullinated peptides from α-enolase (CEP-1), vimentin (Cit-vim), fibrinogen (Cit-fib) and collagen type II (Cit-C1) were measured in serum samples from 10 patients with cyclic citrullinated peptide (CCP) positive rheumatoid arthritis (RA1–RA10) after preabsorption with the different peptides: (A) anti-CEP-1 IgG levels; (B) anti-Cit-vim IgG levels; (C) anti-Cit-fib IgG levels; (D) anti-Cit-C1 IgG levels. Serum samples that were negative for one or several ACPA fine specificities were removed from those specific analyses: RA6 and RA10 (B); RA9 (C); RA5 and RA9 (D). Log scales of AU/ml values are shown on the y-axis. Dotted lines indicate cut-off for positivity (10 AU/ml). The x-axis indicates preabsorbing peptides or no peptide (buffer alone). ***p<0.0001, **p<0.001, *p<0.05 vs no peptide.
HLA-DRB1 SE in combination with PTPN22 and smoking is associated mainly with CEP-1 and Cit-vim positive RA
To investigate associations of genetic and environmental risk factors with different subsets of RA, we first defined RA subsets by the presence or absence of antibodies to single citrullinated peptides—that is, without taking into account which additional ACPA fine specificities may also be present in the subset. As shown in figure 3A, being positive for HLA-DRB1 SE, PTPN22 and smoking was associated mainly with CEP-1 positive RA (OR 39.1) compared with CEP-1 negative/CCP positive RA (OR 7.9). A similar pattern was observed for antibodies to Cit-vim where the association of HLA-DRB1 SE in combination with PTPN22 and smoking was stronger with Cit-vim positive disease OR 25.6 compared with Cit-vim negative/CCP positive disease OR 8.9, figure 3B). Subsets defined by antibodies to Cit-fib or Cit-C1, on the other hand, did not demonstrate such specific associations beyond the effect of CCP (OR 15.6 vs 12.9 for Cit-fib and 10.0 vs 13.8 for Cit-C1; figure 3C,D).

{kind=link}
{kind=link}
{kind=link}
Combined effect of HLA-DRB1 SE, PTPN22 and smoking on disease risk in different subsets of rheumatoid arthritis (RA). ORs with 95% CI were calculated using unconditional logistic regression analysis by comparing individuals positive for the risk factors with individuals who were negative. RA subsets were defined based on the presence or absence of one anticitrullinated protein/peptide antibody (ACPA) fine specificity without taking into account which other ACPA fine specificities may be present simultaneously. Antibody responses to citrullinated peptides from (A) α-enolase (CEP-1), (B) vimentin (Cit-vim), (C) fibrinogen (Cit-fib) and (D) collagen type II (Cit-C1). White bars show subsets negative for the ACPA fine specificity and negative for cyclic citrullinated peptide (CCP) (−/−); grey bars show subsets negative for the ACPA fine specificity but positive for CCP (−/+); black bars show subsets positive for the ACPA fine specificity but where the CCP status was not considered (+/+ or −). 95% CIs for −/+ and +/+ or − subsets are: (A) 4.58 to 13.53 vs 19.07 to 79.97; (B) 5.03 to 15.68 vs 13.95 to 47.02; (C) 7.55 to 0 vs 8.51 to 4; (D) 7.55 to 9 vs 6.07 to 9. Asterisks indicate non-overlapping 95% CI.
Impact of HLA-DRB1 SE on disease risk in different RA subsets
The results described above may be influenced by the coexistence of multiple ACPA fine specificities. We next investigated the four RA subsets which were defined by the sole presence of one ACPA fine specificity—that is, the subsets that were ‘single positive’ for anti-CEP-1, anti-Cit-vim, anti-Cit-fib or anti-Cit-C1 antibodies. In this analysis we focused on HLA-DRB1 SE, the risk factor with the strongest impact. We found a relatively strong association of HLA-DRB1 SE with CEP-1 positive RA (OR 5.1) as well as with Cit-vim positive disease (OR 6.0) but not with Cit-fib positive disease (OR 1.4), and the association with Cit-C1 positive RA was weak (OR 1.6) (table 1). A significant association of HLA-DRB1 SE (OR 2.5, 95% CI 1.8 to 3.6) could also be observed with the subset that was positive for CCP but negative for all four ACPA fine specificities (data not shown).
We then studied the effect of being positive for different combinations of two ACPA fine specificities as the impact of HLA-DRB1 SE may differ between subsets defined by specific combinations of non-cross-reactive ACPAs. The strongest influence of HLA-DRB1 SE was identified for CEP-1/Cit-vim double-positive RA (OR 49.6) which could be compared with an OR of 2.5 for the association with Cit-fib/Cit-C1 double-positive RA. Interestingly, anti-CCP antibody levels were higher in the Cit-fib/Cit-C1 double-positive subset than in the CEP-1/Cit-vim double-positive subset, suggesting that HLA-DRB1 SE mainly influence the fine specificity of the ACPA response rather than anti-CCP levels (see figure 2 in online supplement).
Impact of smoking in combination with HLA-DRB1 SE on disease risk in RA subsets
Since the data presented above do not support an association of HLH-DRB1 SE with the presence of anti-Cit-fib antibodies, we restricted the next analysis to RA subgroups defined by the presence or absence of antibodies to CEP-1, Cit-vim and Cit-C1. We had sufficient power to investigate the combined effect of HLA-DRB1 SE and smoking. As shown in table 2, smoking in the absence of HLA-DRB1 SE did not significantly associate with any RA subset but strengthened the HLA-DRB1 SE association when present. This effect was most pronounced for double-positive and triple-positive subsets compared with single-positive subsets, but only when anti-CEP-1 antibodies were present. Adding Cit-vim reactivity to the CEP-1 single-positive subset increased the OR for the combined HLA-DRB1 SE and smoking association from 12.5 to 18.4. A similar effect was seen when adding Cit-C1 reactivity to the CEP-1 single-positive subset, with an increase in the OR from 12.5 to 17.2. However, in the absence of anti-CEP-1 antibodies, addition of Cit-C1 reactivity to the Cit-vim single-positive subgroup did not increase the OR further (7.3 vs 5.6). The highest OR (23.3) for the combined HLA-DRB1 SE and smoking association was observed for the triple-positive subset (CEP-1+/Cit-vim+/Cit-C1+).
ORs for disease risk in different subsets of rheumatoid arthritis in subjects exposed to different combinations of HLA-DRB1 shared epitope (SE) and smoking compared with non-exposed subjects
Impact of PTPN22 in combination with HLA-DRB1 SE on disease risk in RA subsets
We then investigated the combined effect of HLA-DRB1 SE and PTPN22 in subsets defined by anti-CEP-1, anti-Cit-vim and anti-Cit-C1 antibody status. As seen with smoking, PTPN22 also mainly had an effect in the presence of HLA-DRB1 SE, and this association was also most pronounced for double-positive and triple-positive subsets encompassing anti-CEP-1 antibodies. Addition of Cit-vim reactivity to the CEP-1 single-positive subset increased the OR for the combined HLA-DRB1 SE and PTPN22 association from 8.7 to 29.2 while addition of Cit-C1 to the CEP-1 single positive subset changed the OR from 8.7 to 17.1. However, similar ORs were observed for the Cit-vim single-positive and the Cit-vim/Cit-C1 double-positive subsets (8.4 vs 8.7). The strongest association (OR 34.8) of the HLA-DRB1 SE and PTPN22 combination was seen with the triple-positive subset (CEP-1+/Cit-vim+/Cit-C1+) (see table 2 in online supplement).
Combined effect of HLA-DRB1 SE, PTPN22 and smoking on disease risk in RA subsets defined by anti-CEP-1 and anti-Cit-vim antibody status
In the final analysis we investigated the combined effect of all three risk factors (HLA-DRB1 SE, PTPN22 and smoking) and calculated gene–environment and gene–gene interactions. We focused on what appears to be the two most important ACPA fine specificities in this context and defined RA subgroups based on CEP-1 and Cit-vim reactivity alone (table 3). The presence of HLA-DRB1 SE alleles, PTPN22 polymorphism and cigarette smoking conferred a rather modest risk for the development of CEP-1 negative/Cit-vim negative disease (OR 2.4), while the risk was high for CEP-1 positive/Cit-vim negative RA (OR 24.4) and for CEP-1 negative/Cit-vim positive RA (OR 11.5). However, the highest risk was identified for the CEP-1/Cit-vim double-positive subset (OR 50.1).
ORs for disease risk in different subsets of rheumatoid arthritis in subjects exposed to different combinations of HLA-DRB1 shared epitope (SE), smoking and PTPN22 compared with non-exposed subjects
Analyses of interaction, calculated as the AP,22 ,23 demonstrated significant interactions between HLA-DRB1 SE and smoking in the CEP-1 positive/Cit-vim negative as well as in the CEP-1/Cit-vim double-positive subsets with AP values of 0.33 and 0.64, respectively. Interactions were also observed between HLA-DRB1 SE and PTPN22 in the CEP-1 positive/Cit-vim negative (AP=0.44), the CEP-1 negative/Cit-vim positive (AP=0.56) and in the CEP-1/Cit-vim double positive subsets (AP=0.53) (see table 3 in online supplement).
Discussion
The present study provides novel data concerning the relationships between specific autoimmune reactions and genetic and environmental determinants for the risk of developing RA, with the overall conclusion that HLA-DRB1 SE genes, the PTPN22 risk allele and cigarette smoking are associated with the presence of specific ACPA-reactivities rather than total levels of anti-CCP antibodies.
We have focused our analyses on antibody responses to well-defined citrullinated epitopes on α-enolase,25 ,26 vimentin,13 ,27 fibrinogen28 ,29 and collagen type II,21 representing physiological antigens present in the rheumatoid joint. Most of the patients who were positive for these ACPA fine specificities were also positive in the CCP2 ELISA, the current standard assay for ACPA detection. Although important to note, 18% of the CCP negative patients were positive for at least one ACPA fine specificity. Conversely, nearly 14% of the CCP positive patients were negative for the four ACPA fine specificities, and this subset also showed some association with HLA-DRB1 SE, clearly indicating that other ACPA targets exist, which has also been reported previously25 ,30 ,31 and should be considered in future studies.
Our peptide absorption experiments showed that the ACPAs were mainly non-cross-reactive. Although some degree of cross-reactivity was observed in some serum samples, the majority did not demonstrate cross-reactivity, in line with previous reports.9 ,14 We therefore consider these ACPAs as four distinct fine specificities, and we subsequently based our RA subsets on the presence or absence of anti-CEP-1, anti-Cit-vim, anti-Cit-fib and anti-Cit-C1 antibodies rather than CCP status. The fact that the ACPAs occur together in some patients, but on their own in others, imply that their production may be governed by partly different mechanisms. It is therefore very interesting to note the specific relationship between HLA-DRB1 SE and certain ACPA fine specificities (particularly CEP-1 and Cit-vim) but not others (mainly Cit-fib).
The estimated risks in some of the analyses should be viewed in the context of small sample sizes with large CI. However, our data collectively point to clear differences between ACPA fine specificities, thus confirming and extending previous reports.11,–,13 Importantly, the present study answers the previously unanswered question concerning the influence of HLA-DRB1 SE on the specificity versus the magnitude of the ACPA response.32 Also notable is the fact that the effects of smoking and PTPN22, in the context of HLA-DRB1 SE, appear to be most pronounced in subsets defined by multiple ACPAs where reactivity to CEP-1 is present. These data may suggest that smoking is involved in citrullinating and exposing some—but not other—autoantigens to the immune system, and that PTPN22 exerts its effects only when certain MHC class II-dependent immune reactions are involved. These data will form the basis for future experimental studies on molecular pathways of relevance for disease development in distinct subsets of RA.
In summary, our data highlight the complexity of the gene–environment influence on ACPA-positive RA. A particularly interesting finding is that HLA-DRB1 SE together with PTPN22 and smoking associate so strongly with the presence of anti-CEP-1 antibodies, on the one hand, and anti-Cit-vim antibodies on the other, but mainly with the simultaneous presence of both ACPAs. Since the SE comprises several different alleles and since a dose-effect has been demonstrated for the HLA-DRB1 SE association,4 ,5 ,11 one may speculate that peptides from citrullinated α-enolase may be preferentially presented by one specific SE allele and subsequently trigger the production of anti-CEP-1 antibodies, while peptides from citrullinated vimentin are preferentially presented by another SE allele giving rise to anti-Cit-vim antibodies. Hence, in such a scenario, the presence of two different SE alleles would drive the simultaneous production of two different ACPAs which may tentatively synergise in contributing to the development of RA. Detailed studies of the SE alleles, other MHC class II alleles, as well as other genetic polymorphisms and environmental exposures in the context of different ACPA fine specificities will increase our understanding of the aetiology in different subsets of ACPA-positive RA. The results from such studies may also help to develop future personalised therapies for patients with RA and preventive strategies for individuals at risk of developing the disease.
Acknowledgments
The authors thank the investigators and study participants from the EIRA study.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Web Only Data - This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Web Only Data - This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
- Web Only Data - This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
Karin Lundberg and Camilla Bengtsson are joint first authors. Lars Alfredsson and Lars Klareskog are joint last authors.
-
Contributors KL had main responsibility for the study including design, experimental set-up, analyses, figures, supervising and writing the paper. CB had main responsibility for the statistical analyses and tables (association and interaction analyses). NK and ER performed ELISA assays, cross-reactivity experiments and produced figures with statistics. XJ produced the tables and, together with HK, contributed to the statistical analyses (association and interaction analyses). IP-D performed ELISA assays. LI set up the in-house ELISA assays. CK produced the Cit-C1 and C1 peptides. LP performed the HLA-DRB1 and PTPN22 genotyping. RH had main responsibility for the production of Cit-C1 and C1 peptides. LA supervised the statistical analyses (association and interaction analyses) and, together with LK, had main responsibility for the EIRA case-control study. LK and KL designed the study. All authors contributed to the final paper.
-
Funding The Swedish Research Council, Vinnova, the Swedish Council for Working Life and Social Research, King Gustaf V’s 80-year foundation, the Swedish Rheumatic Foundation, the EU-funded projects Gums&Joints (FP7-Health-2010-261460) and MasterSwitch (FP6-Health-2007-2.4.5-12), the IMI program BTCure (115142-2).
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.