Article Text

Abstract
Objectives Obesity is a risk factor for the development of osteoarthritis (OA) in hands and knees. Adipose tissue can secrete different adipokines with powerful immunomodulatory effects. The infrapatellar fat pad (IFP) is an intra-articular organ in the vicinity of the synovium and cartilage. It is hypothesised that IFP-derived soluble factors could contribute to pathological processes in the knee joint. A study was therefore undertaken to compare the release of inflammatory mediators in the IFP and subcutaneous adipose tissue (ScAT) and to characterise the adipocytes and immune cell infiltrate in these tissues.
Methods Paired IFP and ScAT samples were obtained from 27 patients with primary OA. The stromal vascular cell fraction (SVF) was isolated and characterised by fluorescence activated cell sorting. Cytokine and adipokine release in fat- and adipocyte-conditioned media was measured by luminex.
Results IFP secreted higher levels of inflammatory mediators such as interleukin 6 (IL-6), adipsin, adiponectin and visfatin than ScAT. This could be due to differences in the phenotype of adipocytes and/or in the composition and phenotype of the SVF cells. IFP adipocyte-conditioned media showed a trend towards more IL-6 and adipsin than ScAT. Moreover, the SVF fraction of IFP contained more cells/g tissue, a lower percentage of T cells and a higher percentage of mast cells than ScAT. In addition, T cells had a predominantly pro-inflammatory phenotype while macrophages had a mixed pro- and anti-inflammatory phenotype in the IFP.
Conclusion There are profound differences in secreted inflammatory factors and immune cell composition between the IFP and ScAT. These data indicate that IFP-derived soluble mediators could contribute to pathophysiological processes in the OA knee joint.
Statistics from Altmetric.com
Introduction
Osteoarthritis (OA) is a rheumatic disorder involving mostly weight-bearing joints such as the spine, hip and knee. The onset of disease is a multifactorial process and presents clinically with a heterogeneous disease phenotype. Typically, the entire synovial joint is affected, as evidenced by joint space narrowing, osteophyte formation and subchondral sclerosis.1 2 Risk factors for OA include age, female gender, joint malalignment and obesity.2 Although it is usually accepted that increased mechanical load contributes to joint destruction in obese subjects, recent advances in the physiology of adipose tissue add further insights to the understanding of the relationship between obesity and OA.2 Obesity is not only associated with the development of knee OA, but also with the development of OA in non-weight bearing joints such as the hands.3 4 This could imply a role for adipose tissue-derived factors—in addition to biomechanical factors—as an independent risk factor for the development of OA.
It has become increasingly evident over the last years that adipose tissue, as well as being responsible for the storage of lipids, can also function as an endocrine organ that secretes several factors with potent regulatory effects which influence whole body metabolism.5 Besides adipocytes, adipose tissue contains connective tissue matrix, nerve tissue, endothelial vascular cells and immune cells.6 Studies in mice and men have shown that the immune cell composition differs according to fat depot and varies with body mass index (BMI).7,–,13 Moreover, both adipocytes and infiltrating immune cells actively secrete numerous cytokines and adipokines.8 9 11 14 15 These mediators have profound effects on glucose homeostasis and the regulation of food intake, and also on inflammatory responses.15
The infrapatellar fat pad (IFP), or Hoffa's fat pad, an adipose tissue depot that is located intracapsularly and extrasynovially in the knee joint, has so far received little attention. The function of the IFP in the joint is unknown and there is only limited information to indicate the secretion of some inflammatory mediators by the IFP.16,–,19 Furthermore, adipokines have been shown to be present in the synovial fluid of patients with OA.20 Because it is conceivable that soluble mediators secreted by the IFP could play a role in local inflammatory processes in the joint, we performed an extensive characterisation of the IFP by studying adipokines and cytokines secreted in fat-conditioned media and compared these with subcutaneous adipose tissue (ScAT). These analyses revealed profound differences between the IFP and ScAT in secreted inflammatory factors. To gain insight into the possible cellular origin of these differences, we compared the phenotype of adipocytes and immune cell infiltrate in these adipose tissues and found substantial differences in the composition and phenotype of the constituent cells of the IFP and ScAT.
Methods
Human subjects
Patients with primary OA undergoing joint replacement surgery were recruited into the study and rest material was acquired. IFP and ScAT samples were obtained during knee replacement surgery. ScAT samples were taken from the thigh, next to the incision. Owing to the limited size of the sample, not all experiments could be performed with each tissue sample.
Cell isolation
The isolation of stromal vascular cells was performed as previously described.21 Briefly, tissue fragments were incubated with 1.8 mg/ml collagenase type 1A (Sigma-Aldrich, Zwijndrecht, The Netherlands) in RPMI 1640 (Invitrogen, Breda, The Netherlands) for 1 h at 37°C under continuous rolling. The digested tissue was filtered through a nylon mesh with a pore size of 250 μm and the remaining tissue was mashed and washed through a 70 μm nylon mesh. The cell suspension was washed three times in warm DMEM 4.5 g/l glucose/F12/0.5% BSA/15 mM Hepes/glutamax/pen/strep (Invitrogen) (medium). Erythrocytes were lysed by adding lysis buffer (0.154 mmol/l NH4Cl, 5.7 mmol/l K2HPO4 and 0.1 mmol/EDTA, pH 7.3) for 5 min at room temperature. The cells were washed once and surface stainings were performed. The remaining cells were plated overnight in a six-well plate in a density of 5×106 cells/well maximum in medium supplemented with 50 IU/ml interleukin 2 (IL-2; Peprotech, Hamburg, Germany). After 12 h the cells were stimulated for 5 h with 20 ng/ml phorbol myristate acetate (PMA)/200 ng/ml ionomycin for polyclonal activation of T cells or 10 µg/ml lipopolysaccharide (LPS) for activation of macrophages. Brefeldin A 10 µg/ml was added directly to the LPS-stimulated cells or, after 1 h, to the PMA/ionomycin-stimulated cells. Cells were harvested using a cell scraper and surface and intracellular stainings were performed.
Generation of fat- and adipocyte-conditioned medium
Fat-conditioned medium was obtained by culturing small pieces of IFP and ScAT in vitro at a concentration of 100 mg/ml in medium. After 2 h the medium was refreshed to remove possible contamination with cell rests and blood-derived soluble factors.22 Fresh medium was added for another 24 h before collecting the supernatants. For adipocyte-conditioned medium, adipose tissue was digested with collagenase type 1A (Sigma-Aldrich) and subsequently filtered through a 250 µm nylon mesh. The adipocytes were washed three times by allowing them to float to the surface, followed by removal of medium and addition of fresh medium. The adipocytes were then cultured in six-well plates for 24 h in vitro at a concentration of 100 μl/ml. The supernatants were collected and stored at −80°C until use.
Flow cytometric analysis
Approximately 100 000 unstimulated stromal vascular cells were stained with surface antibody (Ab) solutions containing mixes of the following Abs: PE-conjugated CD3, CD19, FcεRI; FITC-conjugated CD45RA, CD3, CD8; APC-conjugated CD8, CD117; PE-Cy-7-conjugated CD25, CD14 and Pacific Blue-conjugated CD4 (all Abs were from BD Biosciences (Breda, The Netherlands) except the Ab to FcεRI which was from eBiosciencey (Vienna, Austria)). When specified, approximately 400 000 stimulated stromal vascular cells were first stained with the surface Ab solutions. Next, intracellular cytokines were detected upon permeabilisation and fixation using a Cytofix/Cytoperm Fixation/Permeabilization Solution Kit (BD Biosciences) according to the manufacturer's instructions. The following Abs were used for intracellular staining: PE-conjugated Abs to interferon γ (IFNγ), IL-10, tumour necrosis factor α (TNFα), IL-6 (all BD Biosciences except the Ab to IL-6 which was from eBioscience), Alexa fluor 647-conjugated Ab to IL-17 (eBioscience) and the appropriate isotype controls. All incubations were performed on ice for 20 min. The cells were fixed with paraformaldehyde and analysed with a LSR II flow cytometer using Diva 6 software (BD Biosciences).
Detection of adipokines and cytokines
Adipokines and cytokines were measured in supernatants of fat- and adipocyte-conditioned media using the Bio-Plex Pro Human Diabetes kit (Bio-Rad, Veenendaal, The Netherlands), the Bio-Plex array reader and Bio-Plex software in accordance with the manufacturer's instructions. Adipocyte viability was assessed by measuring lactate dehydrogenase activity in IFP- and ScAT-derived adipocyte-conditioned media. No differences were observed between these tissues (data not shown).
Statistical analyses
The Wilcoxon paired rank test was used to compare differences between groups. Correlations were evaluated using the Spearman ranked correlation test. Adjustment for multiple testing was performed when assessing the significance of the studied associations. The adjusted p values ≤0.007, ≤0.013 or ≤0.05 were considered significant, as indicated in each case.
Results
Adipokine and cytokine secretion
Adipose tissue can actively secrete different cytokines and adipokines. It was recently reported that the IFP of patients with OA is a major source of the pro-inflammatory cytokine IL-6.17 Here we confirm and expand these findings by showing that IFP adipose tissue from patients with OA is more active than ScAT and is able to secrete higher amounts of various cytokines and adipokines. We therefore determined the concentration of adipokines and cytokines in fat-conditioned medium of paired samples of IFP and ScAT from patients with primary OA. The characteristics of the patients are shown in table 1. The concentrations of IL-6, TNFα, adipsin, adiponectin, visfatin, leptin and resistin were determined using the luminex technology. Fat-conditioned media of the IFP contained significantly higher concentrations of the pro-inflammatory cytokine IL-6 and of the adipokines adipsin, adiponectin and visfatin than ScAT (figure 1A). Moreover, we observed a trend towards a higher concentration of TNFα in IFP (figure 1A).

Adipokine and cytokine secretion of the infrapatellar fat pad (IFP) and subcutaneous adipose tissue (ScAT). Interleukin 6 (IL-6), tumour necrosis factor α (TNFα), adipsin, adiponectin, resistin, visfatin and leptin concentrations (pg/ml or ng/ml) in (A) fat-conditioned medium and (B) adipocyte-conditioned medium derived from IFP and ScAT. Each line represents samples from one patient. Indicated are means±SEM. p Values were determined using the Wilcoxon signed rank test and a p value ≤0.007 (corrected for multiple testing) was considered significant.
Characteristics of patients with osteoarthritis (OA) undergoing knee/hip replacement
Because both adipocytes and cells in the stromal vascular cell fraction (SVF) can secrete cytokines and adipokines, we wanted to gain insight into the cellular origin of these inflammatory factors. We first compared the soluble factors released by adipocytes isolated from these tissues. IFP-derived adipocyte-conditioned media showed a trend towards higher levels of IL-6 and adipsin, whereas leptin levels were significantly higher in ScAT-derived adipocyte-conditioned media (figure 1B), indicating qualitative differences between the adipocytes in these adipose tissues.
Immune composition of the IFP and ScAT
Immune cells can also secrete a large variety of inflammatory mediators. We therefore compared the immune cell composition of the SVF fraction of the IFP and paired ScAT samples. Owing to the limited sample size obtained from some patients, not all experiments could be performed with each tissue sample. Variable numbers of samples are therefore depicted in different experiments. Flow cytometric analysis of the SVF revealed a heterogeneous forward/side scatter, as was previously published for SVF originating from other adipose tissue depots.23 An example of the gating strategy is shown in figure 2A. A large ‘live’ gate was used to ensure that most cell types were included in further analyses. This was followed by exclusion of doublets and staining with the appropriate Abs. The SVF of the IFP contained significantly more cells/g tissue than ScAT (figure 2B). Different immune cell types were then determined by flow cytometry in both adipose tissues (figure 2A,C). Macrophages (CD14) were the most abundant immune cells present in both adipose tissues followed by T cells (CD3), mast cells (FcεRI+/CD117c+) and B cells (CD19) (figure 2C). Among the cell types identified in these two adipose tissues, the percentage of T cells was significantly higher in ScAT (p=0.013) and the percentage of mast cells was significantly higher in IFP (p=0.012, figure 2C). No differences were observed in the percentages of macrophages and B cells.

Immune composition of the infrapatellar fat pad (IFP) and subcutaneous adipose tissue (ScAT). Cells were isolated from the stromal vascular fraction (SVF) of IFP and ScAT of patients with osteoarthritis undergoing total joint replacement surgery and immune cells were characterised by flow cytometry. (A) Forward scatter (FSC)/side scatter (SSC) of SVF cells. A live gate (first plot) was combined with ‘single cell’ gates (next two plots). A, area of the cells; H, height; W, width. Lineage-specific stainings were performed. An example is shown in the last two plots. (B) Number of cells present in the SVF of IFP and ScAT. (C) Percentage of indicated cell types in each adipose tissue. Each line represents samples from one patient. Indicated are means±SEM. p Values were determined using the Wilcoxon signed rank test and p≤0.05 (B) or p≤0.013 (C) (corrected for multiple testing) were considered significant.
Phenotypic characterisation of T cells in the IFP and ScAT
Because the percentage of T cells differed significantly between the IFP and ScAT, we investigated the phenotype of the CD3 T cells in more detail. No differences in the percentage or phenotype of CD4 or CD8 T cell subsets were detected between paired IFP and ScAT samples (figure 3A). Analysis of CD45RA and CD25 expression revealed that the CD4 and CD8 T cell populations in both adipose tissues consisted mainly of memory cells (CD45RA−). Notably, a significant percentage of CD4 T cells were positive for the activation marker CD25 (figure 3B) in both adipose tissues.
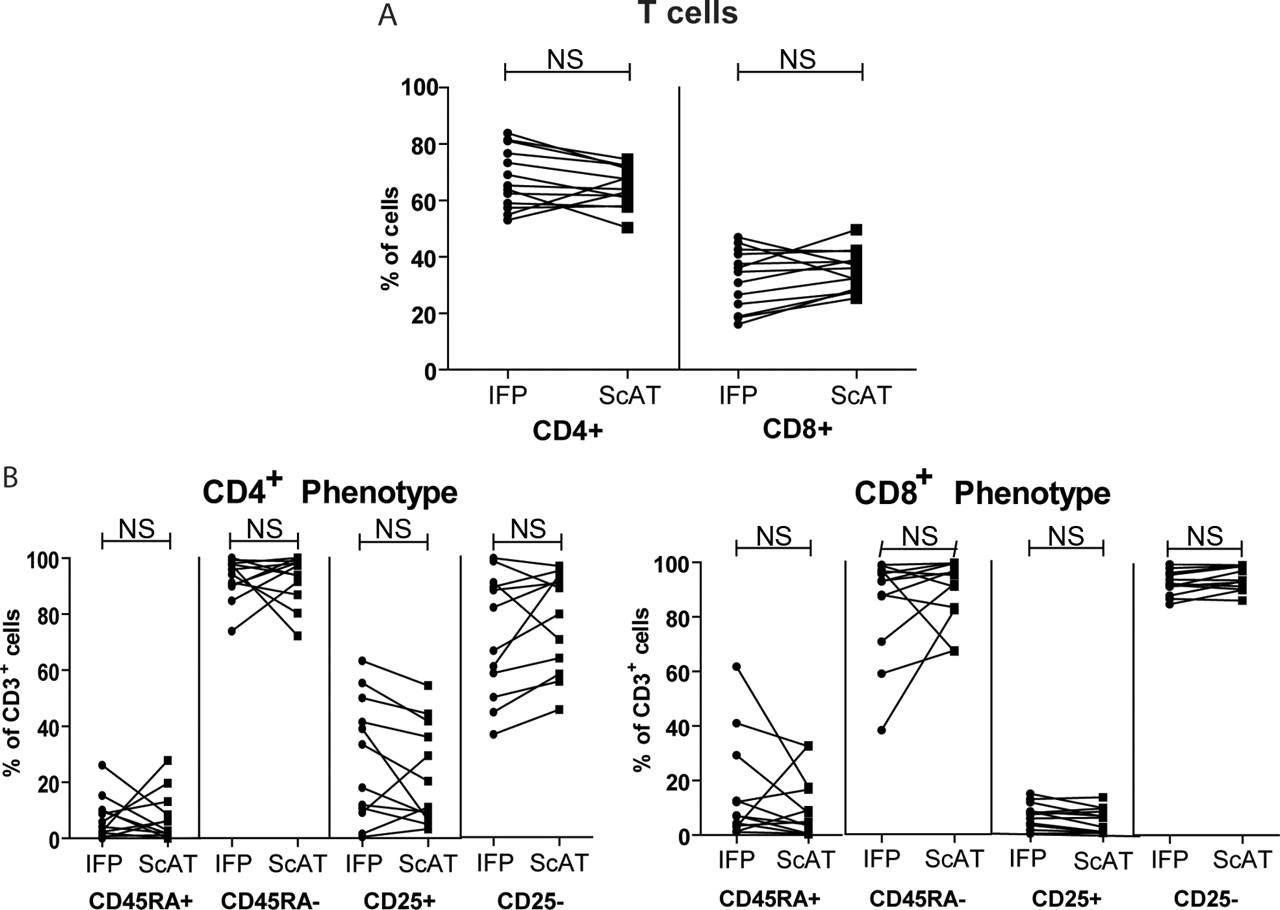
Phenotypic characterisation of T cells from the infrapatellar fat pad (IFP) and subcutaneous adipose tissue (ScAT). (A) Percentages of CD4 and CD8 T cells in paired IFP and ScAT samples. (B) CD45RA and CD25 expression in CD4 (first panel) and CD8 (second panel) T cells. Each line represents samples from one patient. A p value ≤0.013 (corrected for multiple testing) was considered significant.
T cell and macrophage cytokine production in the IFP
Macrophages and T cells are the most abundant immune cell types present in the IFP and ScAT. To assess their possible contribution to the release of inflammatory mediators by adipose tissue, we investigated the polarisation state of these cells. Because the limited sample size precluded these analyses in SVF cells from ScAT, we investigated the polarisation state of CD4 and CD8 T cells and of the CD14 macrophages in the IFP by studying their cytokine-producing capacity on in vitro stimulation of the SVF fraction. Intracellular staining showed that both T cell subsets were able to produce the inflammatory cytokines IFNγ and TNFα, IL-6 and very little, if any, IL-10 and IL-17 (figure 4A), while macrophages produced IL-6 and TNFα as well as IL-10 (figure 4B).

T cell and macrophage cytokine production in the infrapatellar fat pad (IFP). (A) Percentage of cytokine-producing CD4 and CD8 T cells from IFP upon in vitro stimulation with phorbol myristate acetate (PMA)/ionomycin. (B) Percentage of cytokine production by CD14 macrophages upon in vitro stimulation with lipopolysaccharide (LPS). Indicated are mean±SEM percentage cytokine-positive cells. Each dot represents one patient. IFNγ, interferon γ; IL, interleukin; TNFα, tumour necrosis factor α.
TNFα levels in IFP correlate with BMI
It has previously been shown that both the abundance and phenotype of the infiltrating immune cells in adipose tissue varies with BMI.8,–,12 24 We therefore investigated whether the immune cells studied or the cytokine/adipokine production in the IFP and ScAT correlated with BMI. No significant correlations with BMI were found for the percentages of any of the immune cells studied (data not shown). Interestingly, however, a positive correlation of TNFα levels with BMI was found in IFP-derived fat-conditioned media (figure 5A). This correlation is probably not due to adipocyte-secreted TNFα as it was not present in IFP-derived adipocyte-conditioned media (figure 5B). Likewise, this correlation was absent in ScAT-derived fat- and adipocyte-conditioned media. No significant correlations with BMI were found for any of the other cytokines and adipokines in fat-conditioned media or adipocyte-conditioned media in the IFP or ScAT (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Spearman rank test for correlation between tumour necrosis factor α (TNFα) secretion (pg/ml) and body mass index (BMI) in (A) fat-conditioned medium or (B) adipocyte-conditioned medium derived from IFP. Each dot represents one patient. A p value ≤0.007 (corrected for multiple testing) was considered significant.
Discussion
Adipose tissue secretes a large number of soluble mediators that regulate whole body metabolism. It is therefore conceivable that the IFP, through the secretion of different factors, could play a role in local inflammatory processes in the joint. In this initial study we compared cytokine/adipokine production, immune cell composition and phenotype of paired IFP and ScAT samples from patients with OA. Our study revealed marked differences in cytokine and adipokine levels in fat-conditioned media, indicating that the IFP released more inflammatory factors than ScAT. Characterisation of the cells in these tissues revealed that both the composition and phenotype are different between the IFP and ScAT and that both adipocytes and infiltrating stromal vascular cells most probably contribute to the observed differences in release of inflammatory mediators. Finally, we show evidence for a correlation between BMI and the release of TNFα by the IFP, indicating obesity-related inflammatory changes in this organ. In summary, our data indicate that the IFP could play a role in pathophysiological processes in the knee joint through the release of inflammatory mediators.
Adipose tissue can function as an endocrine organ and can actively secrete different cytokines and adipokines.5 15 This is also the case for the IFP, as shown by us and previously by others.17 19 Interestingly, IFP adipose tissue is more active in secreting several cytokines and adipokines than ScAT (figure 1A,B). Some of the soluble factors more actively secreted by the IFP than ScAT—such as IL-6, adiponectin and visfatin—have previously been shown to be able to induce the release of matrix metalloproteinases in vitro.25,–,30 Others like adipsin (complement factor D) are also thought to have pro-inflammatory properties, but their effect on chondrocytes/cartilage is not yet known.
To gain insight into the possible cellular origin of the secreted inflammatory mediators in the IFP and ScAT, we performed an extensive characterisation of the different cell types present in these tissues. Our results reveal substantial differences between cytokines/adipokines secreted by adipocytes from the IFP than ScAT, indicating a more inflammatory phenotype of IFP-derived adipocytes which could explain the differences between these tissues. However, some of the cytokines such as IL-6 and TNFα are produced by several cell types including macrophages, T cells and adipocytes (figures 1B, 4A and B), whereas others probably have a more restricted pattern. Our results do not exclude the possibility that cells in the SVF fraction could also have a different cytokine secretion pattern in the two tissues. Although owing to limited sample size we were not able to assess these differences directly, our data indicate that T cells in the IFP are polarised towards a pro-inflammatory phenotype while macrophages display a mixed pro- and anti-inflammatory phenotype and could thus contribute to the release of inflammatory mediators by the IFP. Moreover, of the tested soluble mediators, TNFα levels in IFP-derived fat-conditioned media showed a positive correlation with BMI whereas the TNFα levels in adipocyte-conditioned media did not (figure 5A,B), providing indirect evidence that cells from the SVF contribute substantially to the total cytokine profile of the IFP. More interestingly, these data suggest a change in the phenotype of SVF cells with BMI, which is indicative of obesity-induced inflammatory changes in the knee-associated adipose tissue. Importantly, this correlation is not seen in ScAT, indicating that the SVF fraction is different in subcutaneous and IFP adipose tissues. The relative contribution of the different cell types within the SVF to the production of adipokines/cytokines by the IFP and ScAT remains to be further investigated.
Previous studies have shown that the immune cell composition of the SVF fraction differs with adipose tissue depot.31 Upon digestion of both adipose tissues, the SVF fraction of the IFP contained a higher total amount of cells/g than ScAT. Flow cytometry analyses of the SVF fractions revealed the presence of macrophages, T cells, B cells and mast cells, with a higher percentage of mast cells in the IFP. In the light of a recent publication which proposes that mast cells in adipose tissue might play a role in angiogenesis,32 we speculate that a higher percentage of mast cells in the IFP could result in higher vascularisation of the IFP compared with ScAT, which could explain the differences in the number of stromal vascular cells in the IFP and ScAT.
Phenotypic characterisation of the CD3 T cells showed that CD4 T cells were the most abundant in both adipose tissues. Interestingly, significantly higher percentages of CD3 T cells were present in ScAT, whereas no differences were observed in the distribution of CD4 and CD8 subpopulations. In line with previous studies in ScAT,9 we found that T cells in the IFP and ScAT had a predominantly memory phenotype (CD45RA−). Although minor contamination with T cells from peripheral blood could not be excluded, the predominance of memory T cells suggests that most of the observed T cells are tissue-infiltrated. The CD4 T cell subset in both adipose tissues contained a considerable percentage of CD25 T cells, indicating the presence of activated T cells in adipose tissue (figure 3B). This finding is interesting in view of a recent publication which suggested that CD4 T cells in mice could undergo antigen-specific expansion in the adipose tissue.25 Further studies are required to address this hypothesis in humans.
A limitation of this study is the lack of information on possible obesity-associated comorbidities such as type 2 diabetes or cardiovascular disease in our patients. These could affect our results. However, by comparing only paired IFP and ScAT patient samples we believe that the systemic effects of these complications have been largely corrected for and the differences that we show are inherent to the tissues studied.
In conclusion, our results show a more inflammatory phenotype of the IFP compared with ScAT in patients with OA. Both adipocytes and infiltrating stromal vascular cells are likely to contribute to this phenotype, as suggested by profound differences in the composition and phenotype of these cells in these tissues. Our data indicate that the IFP could contribute to the pathophysiological changes in the OA joint via the local production of cytokines and adipokines.
Acknowledgments
The authors thank E van Beelen for technical assistance with the cytokine measurements.
References
Footnotes
-
Funding This work was financially supported by TI-Pharma, EU FP6 program Autocure, FP7 program Masterswitch and a grant from the Centre for Medical Systems Biology (CMSB) within the framework of the Netherlands Genomics Initiative (NGI).
-
Ethics approval This study was conducted with the approval of the Leiden University Medical Center.
-
Provenance and peer review Not commissioned; externally peer reviewed.