Article Text

Abstract
Objectives: To better understand the role of collagenase-3 (MMP-13) in joint inflammation by investigating the consequences of transient overexpression of human collagenase-3 (matrix metalloproteinase-13 (MMP-13)), introduced by adenoviral gene delivery, in the mouse knee joint.
Methods: A single dose (5×107 pfu) of recombinant adenovirus coding either for β-galactosidase (RAdLacZ) or human MMP-13 (RAdMMP-13) was injected intra-articularly into the knee joint of adult mice. The joints were analysed at frequent intervals up to 4 weeks by histology, immunohistochemistry, and RNA analysis.
Results: When RAdLacZ reporter virus was used, adenoviruses efficiently infected synovial cells, chondrocytes of articular cartilage, and hypertrophic chondrocytes of the growth plate. The infection was transient as no reporter gene activity was detected 3 weeks after the injection. After RAdMMP-13 injection into the knee joints, expression of human MMP-13 in joint tissues resulted in an arthritis characterised by recruitment of inflammatory cells and increased production of cytokines and chemokines, synovial hyperplasia, and pannus formation. After the loss of MMP-13 transgene expression at 3 weeks, these inflammatory changes began to diminish.
Conclusions: MMP-13 has a role in the onset of inflammatory reaction in synovium. However, damage to articular cartilage was only rarely detected after the short term overexpression of MMP-13.
- arthritis
- matrix metalloproteinases
- adenovirus
- inflammation
- mice
- FGF, fibroblast growth factor
- IL, interleukin
- MMP, matrix metalloproteinase
- PBS, phosphate buffered saline
- RAdLacZ, recombinant adenovirus coding for β-galactosidase
- RadMMP-13, recombinant adenovirus coding for MMP-13
- RT-PCR, reverse transcriptase-polymerase chain reaction
- TC, threshold cycle
- TNFα, tumour necrosis factor α
Statistics from Altmetric.com
- FGF, fibroblast growth factor
- IL, interleukin
- MMP, matrix metalloproteinase
- PBS, phosphate buffered saline
- RAdLacZ, recombinant adenovirus coding for β-galactosidase
- RadMMP-13, recombinant adenovirus coding for MMP-13
- RT-PCR, reverse transcriptase-polymerase chain reaction
- TC, threshold cycle
- TNFα, tumour necrosis factor α
Arthritides comprise a heterogeneous group of inflammatory degenerative disorders, characterised by different levels of chronic inflammation, invasive synovitis, destruction of cartilage and bone, and narrowing of the joint cavity. Interplay between resident connective tissue cells of the joints, such as synoviocytes and chondrocytes, and the infiltrating inflammatory cells has an important role in the pathogenesis of synovitis in the affected joint.1 This inflammatory process is largely mediated by inflammatory cell derived chemokines and cytokines, which in turn activate synoviocyte proliferation and secretion of other cytokines, chemokines, and proteolytic enzymes. Among the latter, matrix metalloproteinases (MMPs) are believed to play a central part in the inordinate matrix degradation, thus mediating destruction of joint architecture.1–3
MMPs are a family of structurally related, zinc dependent endopeptidases with a combined activity to degrade essentially all components of the extracellular matrix.4 They not only have an important function during development and in many physiological events but are also blamed for much of the excessive matrix degradation in numerous pathological conditions, ranging from arthritis to cancer invasion and metastasis.4,5 In addition, MMPs participate in cell-cell and cell-matrix signalling by modulating the activity of growth factors, cytokines, and chemokines such as tumour necrosis factor α (TNFα), transforming growth factor β, fibroblast growth factors (FGFs), and monocyte chemoattractant protein-3 (MCP-3).6–9
In articulating joints MMPs are mainly produced by synovial cells and chondrocytes, which maintain the homoeostasis of cartilage matrix by producing new matrix components as well as degrading enzymes in a dynamic equilibrium.10 Disturbances in this balance may lead to development of pathological conditions; as has been shown in mice deficient in MT1-MMP (MMP-14), which develop arthritis and fibrosis of joint tissues.11 In normal joints, matrix turnover seems to be quite low, and the mRNA and protein levels as well as the activities of most MMPs are undetectable. In contrast, increased production of several MMPs (MMP-1, -2, -3, -7, -9, -12, -13, -14, -16, and -19) by synovial cells, chondrocytes, and inflammatory cells has been demonstrated in arthritic diseases.12–14 The exact role of most of these proteolytic enzymes in the pathogenesis of disease is, however, still unclear.
Among the members of the collagenase subfamily of MMPs—the principal mammalian MMPs capable of cleaving native fibrillar collagens—collagenase-3 (MMP-13) has evoked attention as a potentially important mediator of tissue destruction in many pathological conditions owing to its wide substrate specificity and normally highly stringent control of expression. Of the three collagenases, MMP-13 is the most efficient to degrade type II collagen—the main constituent of articular cartilage.15 MMP-13 can also cleave other cartilage collagens, such as type IX collagen and aggrecan, the most abundant proteoglycan in articular cartilage.16,17 Expression of MMP-13 has been reported in rheumatoid synovium18,19 and in osteoarthritic cartilage.15,20 Recently, overexpression of an active mutant of human MMP-13 in articular cartilage chondrocytes was shown to result in cartilage degradation in a transgenic mouse model.21
To further elucidate the role of MMP-13 in the pathogenesis of cartilage destruction we used adenovirus mediated gene transfer to transiently overexpress human MMP-13 in the mouse knee joint. Our results indicate that expression of human MMP-13 induces a transient arthritic phenotype and synovial inflammation in the mouse knee joint.
METHODS
Adenovirus vectors and experimental protocol
Recombinant adenoviruses harbouring the bacterial β-galactosidase gene (RAdLacZ)22 or the full length cDNA for human MMP-13 (RAdMMP-13)23 under the control of cytomegalovirus promoter were used. Empty control virus (RAd66) (kindly provided by Dr GW Wilkinson, University of Cardiff, Wales) was used as an additional control.
For adenovirus injections, 10–12 week old female C57 black mice were anaesthetised with an intraperitoneal injection of 2.5% Avertin 0.015 ml/g. After disinfection with 70% ethanol, either one or both knee joints were injected laterally with 5×107 pfu of adenoviral construct RAdLacZ, RAdMMP-13, or RAd66 in phosphate buffered saline (PBS). For the analysis of β-galactosidase expression, the animals were killed 2 days, and 1, 2, 3, and 4 weeks after the injection. Systematic histological and molecular biological analyses of joints treated with RAdMMP-13 were performed at 3 days, 1 week, and 3 weeks after the injection. Uninjected, PBS injected, RAd66 injected, and RAdLacZ injected knees were included as controls in these series. The study protocol was approved by the institutional committee for animal welfare.
Determination of β-galactosidase activity
The hind limbs were dissected free of skin and superficial muscle layers, fixed, and stained in X-Gal solution.24 After washing, the knees were photographed for visual estimation of the X-Gal derived chromogen. Uninjected knees were used as negative controls. Some RAdLacZ injected knees were not incubated with X-Gal, but processed directly for immunohistochemical determination of β-galactosidase.
Histological grading
Dissected limbs were fixed in 4% paraformaldehyde, demineralised in 10% EDTA for 5–20 days, dehydrated, embedded in paraffin, sectioned sagittally, and stained with haematoxylin and eosin. For the assessment of intra-articular changes and the degree of synovitis, a modified qualitative grading system was used.25 Synovial hyperplasia, fibrosis, inflammation, and necrosis were used as criteria in the scoring (table 1).
Qualitative grading system used to assess the degree of synovitis in the mouse knee joint, modified from Podworny et al25
RNA extraction and analysis
For RNA analyses, knee samples were dissected at the femoral and tibial growth plates and prepared free of surrounding muscles. The samples thus contained femoral and tibial articular cartilages and epiphyses, menisci, tendons, synovial tissue, and the surrounding ligaments. Before RNA extraction using the guanidinium isothiocyanate method and sedimentation through a CsCl gradient,26 the samples were pulverised under liquid nitrogen.
The expression of human MMP-13 mRNA in RAdMMP-13 injected knee joints was determined by reverse transcriptase-polymerase chain reaction (RT-PCR). Aliquots of total RNA (100 ng) were reverse transcribed into cDNA and a 300 bp fragment of human MMP-13 cDNA corresponding to nucleotides 534–833 was amplified using the RobusT II RT-PCR kit, according to the manufacturer’s instructions (Finnzymes, Finland), and oligonucleotides specific to the human sequence. Specific oligonucleotides were also used to amplify transcripts of mouse glyceraldehyde-3-phosphate dehydrogenase, a reference control gene. The PCR products were electrophoresed on a 3% agarose gel and visualised by ethidium bromide staining.
For northern hybridisations, aliquots (10 μg) of total RNA were denatured with glyoxal and dimethylsulphoxide, electrophoresed on 0.75% agarose gels, transferred by blotting into Pall Biodyne membranes, and hybridised with 32P labelled cDNAs for murine MMP-13 and ribosomal 28S RNA under conditions recommended by the supplier of the membrane (Pall Europe, Portsmouth, UK). After high stringency washes, the bound radioactivity was detected and quantified on a molecular phosphoimager (Bio-Rad, Hercules, CA).
Taqman RT-PCR
cDNA was synthesised from 1 µg of total RNA in a 50 µl reaction mix using MultiScribe reverse transcriptase and random hexamers (Applied Biosystems, Foster City, CA), according to the manufacturer’s instructions. After primer incubation and reverse transcriptions, the amplifications were performed using reagents from Applied Biosystems in a Mastercycler gradient thermal cycler (Eppendorf, Hamburg, Germany). PCR primers and probes were purchased from Applied Biosystems as predeveloped reagents (18S ribosomal RNA, interleukin (IL) 1β, TNFα, MCP-1, MIP-1α, RANTES) or were designed (MCP-2) by PrimerExpress version 2.0 software.
Real time quantitative PCR was performed with the AbiPrism 7700 sequence detector system, according to the instructions of the supplier (Applied Biosystems). For cDNA amplification, a 10 minute incubation at 95°C was done to activate AmpliTaqGold DNA polymerase; this was followed by 40 cycles with 15 seconds at 95°C and 1 minute at 60°C for each cycle. PCR amplification of the endogenous 18S rRNA was performed for each sample to control sample loading and to allow normalisation between samples, according to the manufacturer’s instructions (Applied Biosystems). The probes for target genes were 5′ end labelled with a reporter dye 6-carboxyfluorescein (FAM), whereas the probe for the endogenous control rRNA was 5′ end labelled with fluorescent dye VIC. Accumulation of PCR products was detected directly by monitoring the increase in fluorescence of the reporter dye. The results were expressed as relative units (fold differences)—that is, the ratio of PCR amplification efficiency of sample cDNA to that of the calibrator (FAM TC calibrator = 40, VIC TC calibrator = 11.0→ΔTC calibrator = 29.0, where TC means a threshold cycle).
Relative units were calculated by the comparative TC method. Firstly, the TC for the target amplicon and the TC for the endogenous control (18S rRNA) were determined for each sample. Differences in the TC for the target and the TC for the endogenous control, called ΔTC, were calculated to normalise for the differences in the amounts of total nucleic acid added to each reaction mixture. The ΔTC of the calibrator (29.0) was subtracted from the ΔTC of each experimental sample and termed the ΔΔTC. The amount of target normalised to an endogenous control and relative to the calibrator was then calculated by the equation 2−ΔΔTC.
Immunohistochemistry
Paraffin embedded samples were cut into 5 μm sections, which were deparaffinised, rehydrated, and digested for 1 hour with hyaluronidase (2 mg/ml) in PBS (pH 5). For the detection of LacZ expression, polyclonal antibodies against bacterial β-galactosidase raised in rabbits (Biodesign, Saco USA) were applied in dilutions of 1:50 and 1:100 in PBS containing 1% bovine serum albumin for an overnight incubation at 4°C. After rinses with PBS, a biotin conjugated secondary antibody was applied for 20 minutes at room temperature. After washes, the slides were incubated with alkaline phosphatase conjugated streptavidin for 30 minutes. The sections were counterstained with haematoxylin.
The distribution of MMP-13 was studied with polyclonal sheep antibodies prepared against human MMP-13, which also detected murine MMP-13 (Biotrend, Koln), in a dilution of 1:100 as above. After an overnight incubation at 4°C the sections were processed further as described for LacZ. The production of human MMP-13 was studied with a 1:100 dilution of a monoclonal antibody detecting specifically the human MMP-13 protein (Calbiochem, San Diego, CA). After rinses with PBS, a biotin conjugated secondary antibody was applied for 10 minutes at room temperature. After washes, the slides were incubated with streptavidin conjugated horseradish peroxidase for 10 minutes (Histo-Mouse kit, Zymed, South San Francisco, CA). Colour was developed with diaminobenzidine, and the sections were counterstained with haematoxylin.
RESULTS
Capacity of adenovirus to infect cells of the knee joint
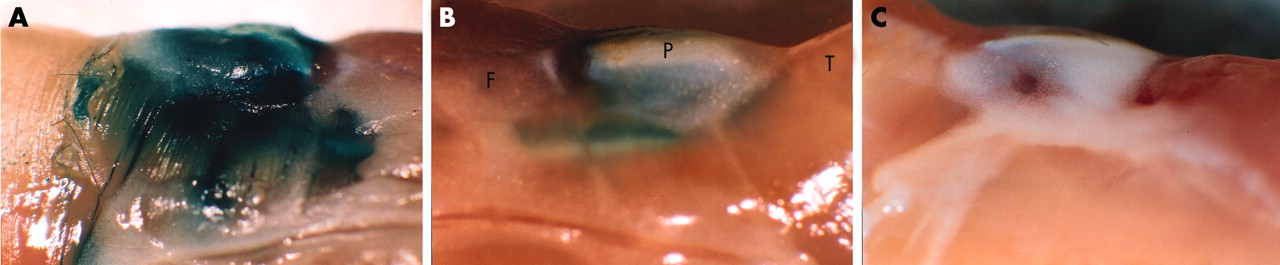
The in vivo transduction efficiency of the adenoviruses was first tested by intra-articular injection of RAdLacZ into the knee joints of 12 week old mice. Strong β-galactosidase activity was detected by enzymatic staining already at two days after the injection and persisted through the first week (fig 1A). Thereafter the staining intensity started to decrease (fig 1B) and became undetectable by 4 weeks (not shown). No staining could be seen in the contralateral knee joint (fig 1C) or in the other control joints.
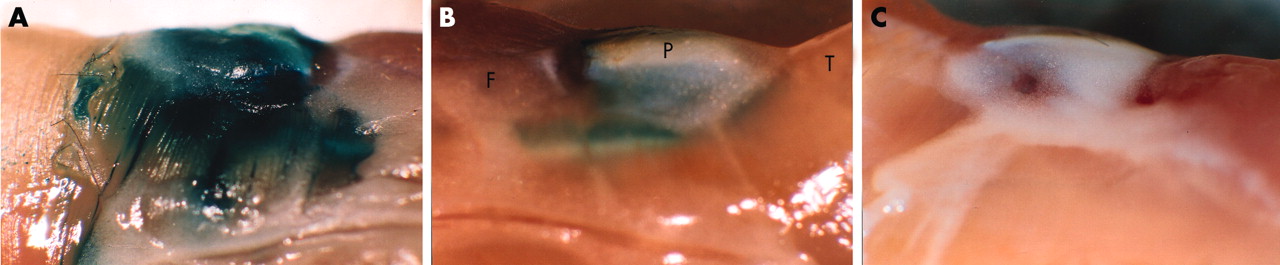
The in vivo transduction efficiency of RAdLacZ. Lateral view of mouse knee joints (A) 1 and (B) 2 weeks after intra-articular injection of 5×107 pfu of RAdLacZ. The joint was dissected as a block, fixed, and stained with X-Gal. The contralateral, uninjected knee exhibited no β-galactosidase activity as shown 1 week after the injection (C). P, patella; T, tibial muscles; F, femoral muscles.
Localisation of cells expressing the β-galactosidase activity was performed by immunohistochemical analysis of sagittal sections of knee joints 1 week after the injection. These analyses showed that recombinant adenoviruses can efficiently infect several different cell types in the joint. Synovial cells were infected most efficiently; about 70% of the synoviocytes showed immunoreactivity for β-galactosidase (fig 2A). Synovial lining cells were less frequently positive for β-galactosidase than synoviocytes in deeper layers of the synovium. Also chondrocytes in the articular cartilage, especially those in the intermediate zone, stained positive for β-galactosidase (fig 2B), as did hypertrophic chondrocytes in the growth plate (fig 2C). Uninjected control animals exhibited no staining for β-galactosidase (figs 2D, E, and F).

Recombinant adenoviruses infect different tissues and cells in the knee joint. Immunohistochemical localisation of β-galactosidase in mouse knee joints using a monoclonal antibody was performed 1 week after injection with either RAdLacZ (A, B, and C) or a control virus RAd66 (D, E, and F). Representative sections of synovium (A and D), articular cartilage (B and E), and growth plate (C and F) are shown. These immunohistochemical analyses were also used to measure the transduction efficiency of RAdLacZ to various cells and tissues of the knee joint. Scoring shown to the left is based on the percentage of the cells positive for β-galactosidase: −, no positive cells; +/−, 1–24%; +, 25–49%; ++, 50–74%; +++, 75–100%.
Adenoviral expression of MMP-13 induces synovial inflammation
A single intra-articular injection of RAdMMP-13 resulted in a condition resembling inflammatory synovitis characterised by influx of inflammatory cells, synovial hyperplasia, and pannus formation (fig 3A). Control joints treated with an equal amount of RAdLacZ or the empty vector RAd66 showed considerably less evidence of inflammatory or other changes than uninjected litter mates in systematic histological examination and scoring based on criteria summarised in table 1 (fig 3B).

Overexpression of MMP-13 results in inflammatory synovitis. Histopathology of RAdMMP-13 treated mouse knee joints 1 week after injection. (A) Photomicrograph of a sagittal section of the inflamed knee joint; (B) qualitative analysis of synovitis based on synovial hyperplasia, fibrosis, inflammation, and necrosis in the RAdMMP-13, RAdLacZ, and RAd66 injected knees (n = 5–6 in each group). Values are the mean (SD); (C) synovial hyperplasia; (D) inflammatory reaction centre consisting of mononuclear cells; (E) invasive pannus formation; (F) superficial erosion of articular cartilage and meniscus were found in the RAdMMP-13 injected joints. The sections were counterstained with haematoxylin and the scale bar represents 800 µm (panel A), 200 µm (panels C and E), and 100 µm (for panels D and F). ac, articular cartilage; b, bone; m, meniscus.
The strong inflammatory response in the joints of animals injected with RAdMMP-13 was most evident 7 days after the injection. The histopathological alterations included hyperplasia throughout the synovial tissue (fig 3C), an increased number of mainly mononuclear infiltrating cells (fig 3D), and an increased number of cells with synovial fibroblast-like morphology. In some areas the synovium adhered to articular cartilage forming a pannus-like structure, which invaded the underlying cartilage surface (fig 3E). Meniscal degradation and superficial fibrillation of articular cartilage were also seen in a limited number of RAdMMP-13 treated knee joints as shown in fig 3F. No systemic effects were detected after RAdMMP-13 injection.
The inflammatory reaction started to diminish after the first week. At 21 days, synovial hypertrophy was markedly decreased, the presence of inflammatory cells less abundant, and no progressive cartilage degradation could be detected. The synovial lining, however, remained thickened.
Endogenous MMP-13 expression is up regulated after RAdMMP-13 treatment
Analysis of adenovirally delivered human MMP-13 mRNA expression by RT-PCR showed that the mRNA could be detected 3 days after adenovirus injection, but not any more 7 or 21 days after injection (fig 4A). Analysis of endogenous murine MMP-13 mRNA levels by northern hybridisation showed significantly increased levels 1 week after injection, compared both with uninjected controls (p = 0.001) and with the RAdLacZ injected controls (p = 0.017) (fig 4B). This increase in murine MMP-13 mRNA production was transient; by 3 weeks the mRNA levels had returned to control levels (fig 4B).
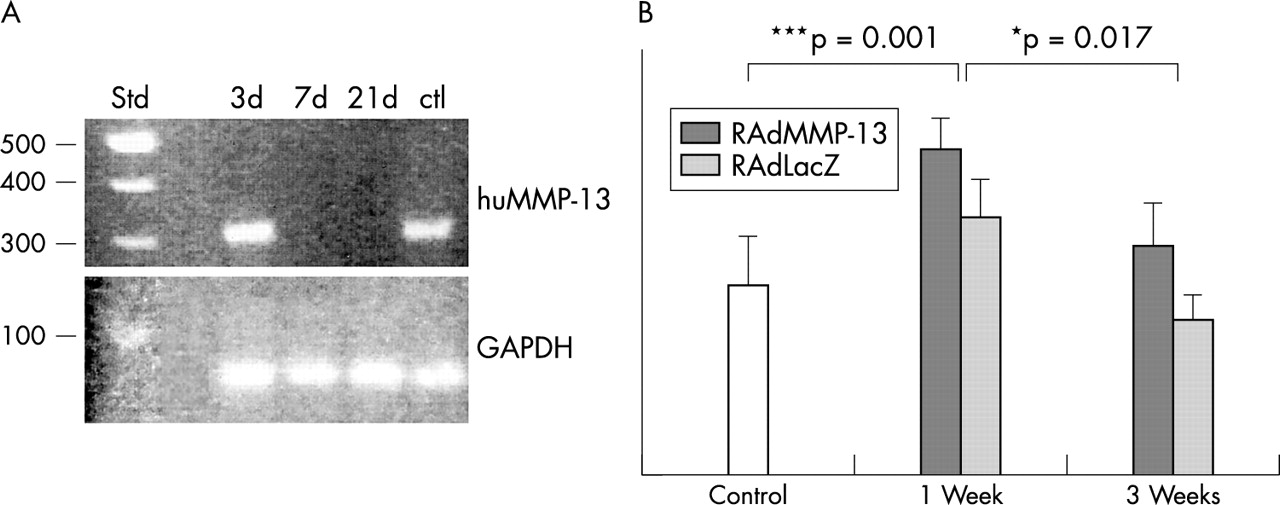
RAdMMP-13 injection increased MMP-13 mRNA production in the knee joint. (A) RT-PCR analysis of human MMP-13 mRNAs in RAdMMP-13 injected knee joints. RNA extracted from a squamous cellular carcinoma sample was used as a positive control. Aliquots of RNA extracted 3, 7, and 21 days after the injection were used for cDNA synthesis by reverse transcriptase reaction and subsequent amplification by PCR. The reaction products were electrophoresed on a 1% agarose gel with molecular weight markers. The expected human MMP-13 derived band of 300 bp is seen migrating close to the 300 bp fragment of the standard. (B) Compiled data from northern analysis of total RNAs extracted from intact, RAdLacZ, or RAdMMP-13 injected knee joints at 1 and 3 weeks after injection (n = 6 at each time) using a probe for murine MMP-13 and 28S ribosomal RNA. The hybridisation intensity was analysed with a phosphoimager and normalised to a constant amount of 28S RNA. Values are mean (SD). Northern analysis: *p<0.05.
Localisation of human and mouse MMP-13 in articular tissues after RAdMMP-13 injection
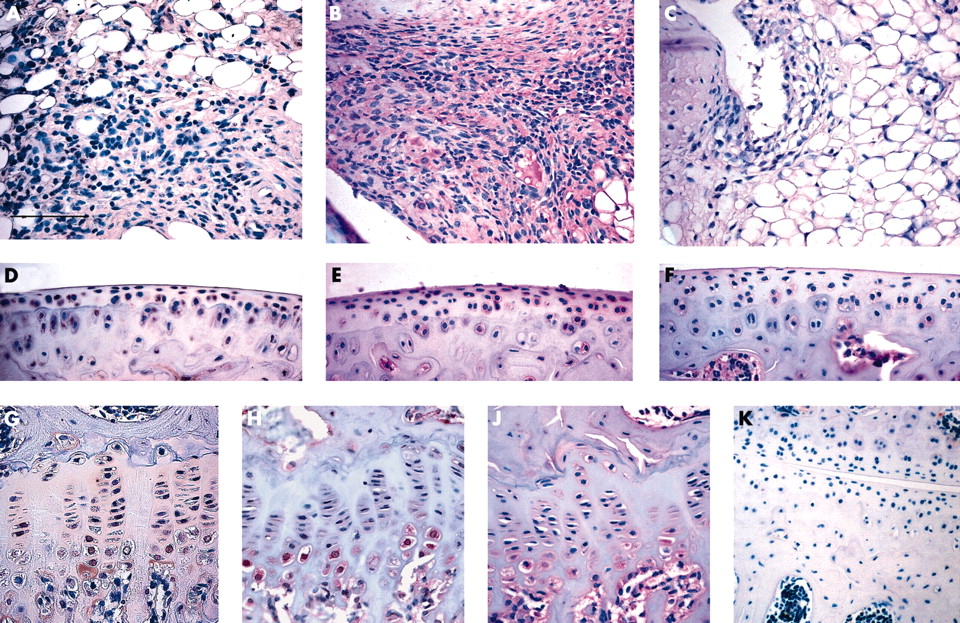
Immunohistochemical localisation of human MMP-13 in articular tissue after a single RAdMMP-13 injection was similar to that of β-galactosidase after RAdLacZ injection. Human MMP-13 was seen in the synovium, predominantly in the fibroblast-like cells of hypertrophic synovium (fig 5A), and in growth plate, especially in the hypertrophic chondrocytes, where staining was seen both intracellularly and pericellularly (fig 5G). In articular cartilage, human MMP-13 production was seen in the pericellular area around the chondrocytes of articular cartilage (fig 5D).

Immunohistochemical localisation of MMP-13 production in the joint. Immunohistochemical localisation of human MMP-13 production in the synovium (A), articular cartilage (D), and growth plate (G) 1 week after injection of RAdMMP-13 using a monoclonal antibody recognising only human MMP-13. Binding of the antibody was detected as a reddish brown precipitate using the avidin-biotin complex method. As a control (K) the monoclonal antibody against human MMP-13 was also applied on a section of uninjected control knee. Localisation of both human and mouse MMP-13 by immunohistochemistry in joint tissues 1 week after injection of RAdMMP-13 (B, E, and H) or in uninjected knee joints (C, F, and J) using polyclonal antibody (B and C synovium, E and F articular cartilage, H and J growth plate). Binding of antibodies was detected using biotin labelled secondary antibody and alkaline phosphatase conjugated streptavidin. The sections were counterstained with haematoxylin. The scale bar in panel A represents 100 µm for panel K and 50 µm for the other panels.
Using another anti-MMP-13 antibody, which recognises both human and murine MMP-13 proteins, MMP-13 was found to be produced throughout the hypertrophic synovium (fig 5B) and also in areas where no staining for human MMP-13 was detected. Production of endogenous MMP-13 was also induced in articular cartilage of RAdMMP-13 injected animals. Whereas only relatively few chondrocytes in the superficial zone of the articular cartilage expressed human MMP-13 after RAdMMP-13 injection, most of these chondrocytes exhibited positive pericellular staining with antibodies detecting both the human and mouse MMP-13 proteins. The staining for this antibody could also be observed more diffusely in the matrix compared with that seen for the antihuman MMP-13 antibody (figs 3D and E). In intact knee joints MMP-13 was detected in hypertrophic chondrocytes of the growth plate and in a few chondrocytes in the uncalcified articular cartilage, but no staining for MMP-13 could be found in the normal synovium. (figs 5C, F, and J).
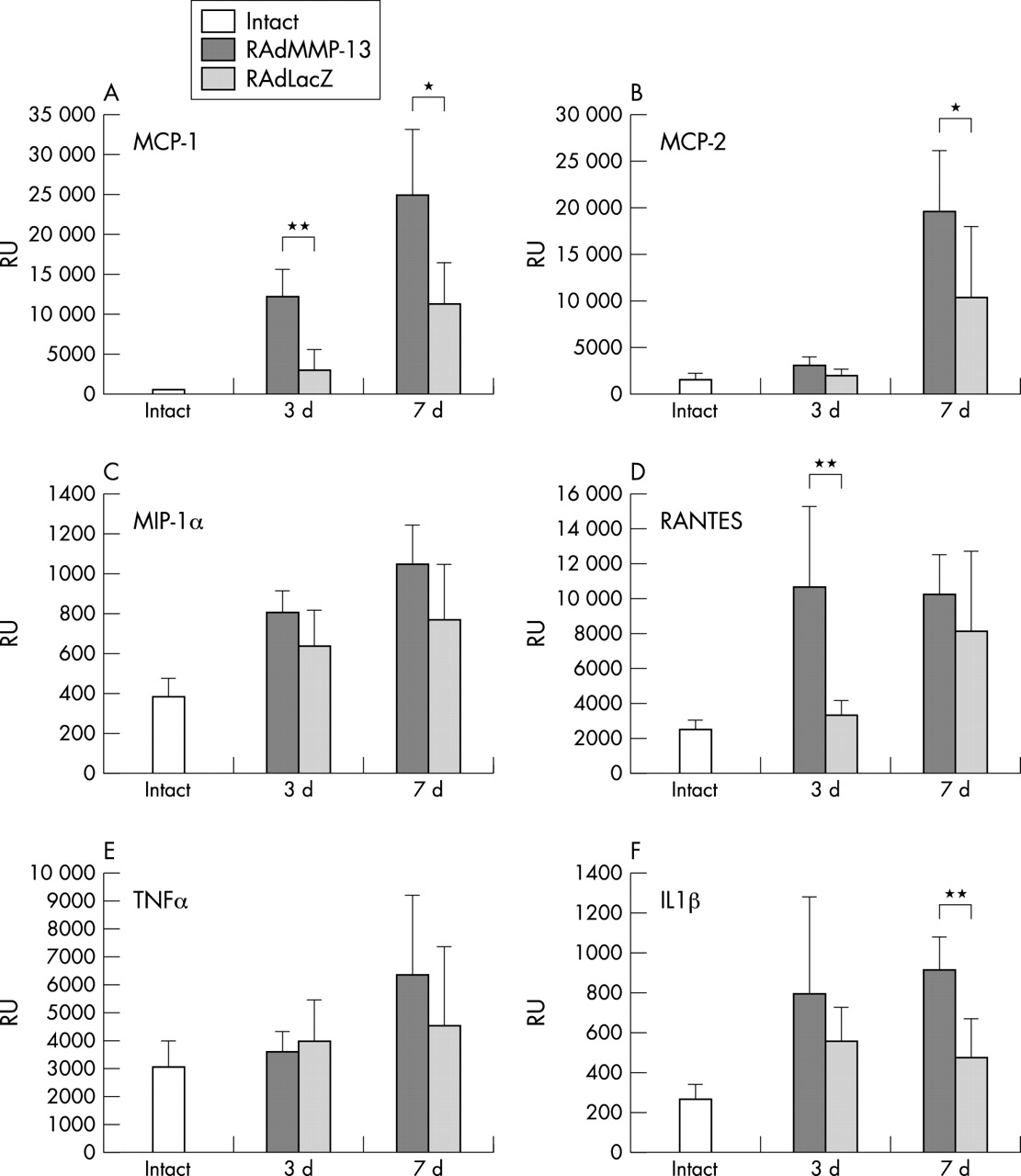
Assessment of inflammatory response with a quantitative RT-PCR method
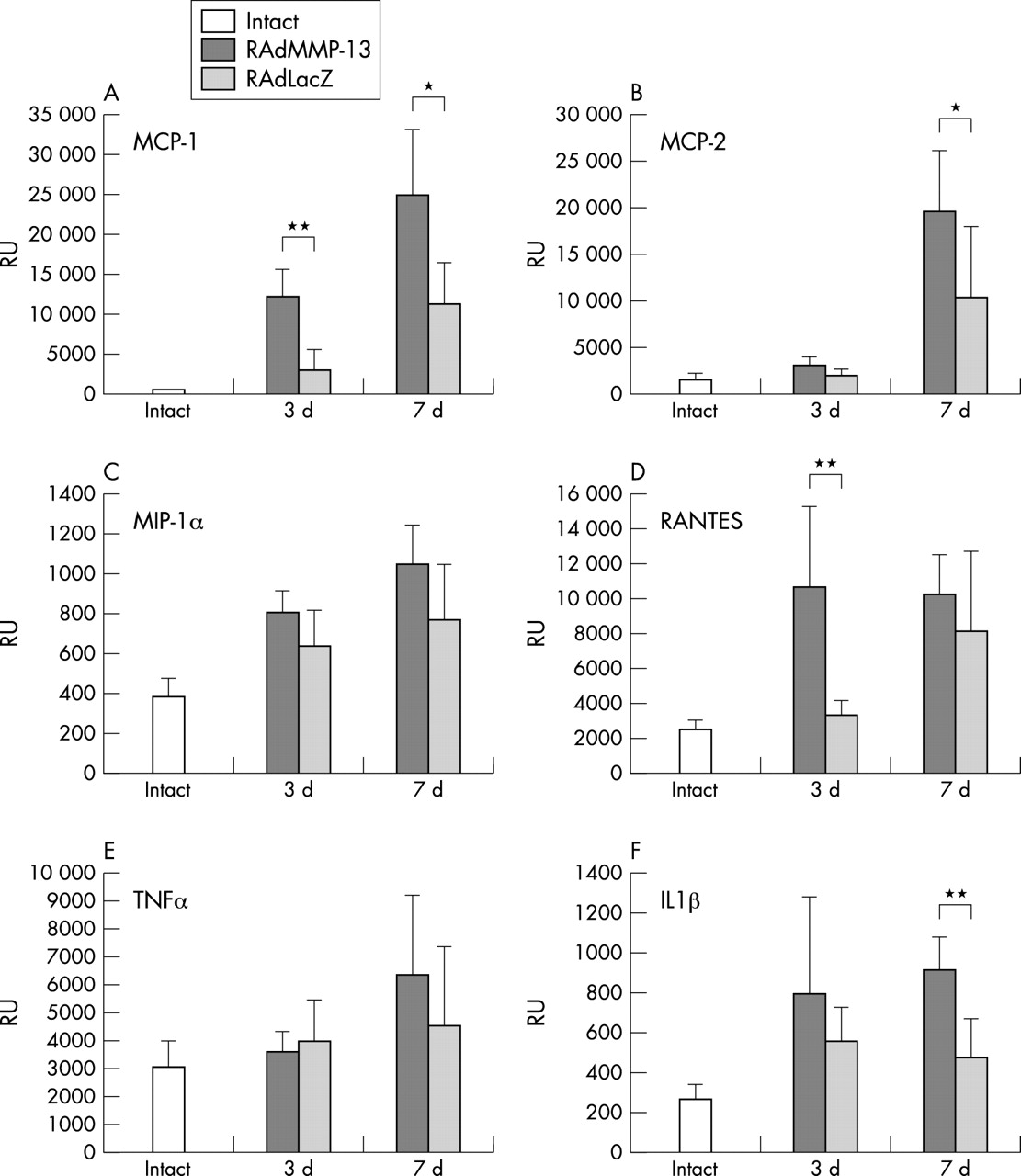
To obtain a quantitative estimate of the inflammatory response to intra-articular RAdMMP-13 injection, a quantitative RT-PCR was used to study the production of selected inflammatory chemokines and cytokines at 3 and 7 days after injection, and in uninjected knee joints (fig 6). Production of MCP-1 and RANTES mRNAs was significantly increased in RAdMMP-13 injected knee joints compared with uninjected and RAdLacZ injected joints (p = 0.0013 and p = 0.0073, respectively) already 3 days after injection (figs 6A and D). IL1β mRNA was also up regulated at 3 days in RAdMMP-13 injected knee joints compared with intact and RAdLacZ injected samples (fig 6F). The mRNA levels of MCP-1, RANTES, and IL1β remained raised in RAdMMP-13 injected animals at 7 days after injection.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of chemokines and cytokines is up regulated after RAdMMP-13 injection Production of mRNAs for (A) MCP-1, (B) MCP-2, (C) MIP-1α, (D) RANTES, (E) TNFα, and (F) IL1β mRNA production was measured in intact, RAdMMP-13, and RAdLacZ injected mouse knee joints (n = 5–6 per group) using a quantitative Taqman RT-PCR method. The results were normalised for a constant amount of 18S ribosomal RNA. For statistical analyses a Wilcoxson matched pair test was used: *p<0.05, **p<0.005 significance of mRNA levels in RAdMMP-13 treated knee joints over those injected with RAdLacZ.
The level of MIP-1α mRNA was increased at 3 and 7 days after injection compared with non-injected knee samples, but the increase was of the same magnitude also in the RAdLacZ injected knees (fig 6C). The mRNA level of MCP-2 was markedly up regulated in RAdMMP-13 injected knee joints only at 7 days after injection (fig 6B). Interestingly, no marked up regulation in TNFα mRNA production could be seen in either group at 3 or 7 days after the injection (fig 6E).
DISCUSSION
In this study we report that a single injection of RAdMMP-13 into the knee joints of mice results in a transient synovitis, characterised by recruitment of inflammatory cells and an increase in intra-articular production of many different cytokines and chemokines, followed by synovial hyperplasia and pannus formation. These findings are in agreement with earlier observations of increased levels of many MMPs in the synovial fluid and tissue of arthritic patients, and the ability of MMP-13 to degrade the major constituents of articular cartilage.15,27
We chose to use adenoviral gene transfer in order to elucidate the role of MMP-13 in the development of arthritic disease, because adenoviral vectors have been shown to provide efficient transient expression in joint tissues.28,29 However, the well documented immunogenicity of adenoviruses clearly hinders their use in clinical trials. The strength of the cell mediated immune response to adenovirus, which eventually shuts down the expression of any gene delivered, varies between animals and strains used and depends also on the amounts of virus used.29,30 Initially, we tested different dosages of the RAdMMP-13 virus for the intra-articular injections. For the final experiments the lowest amount of the virus with which the biological effect could be achieved was used to minimise the possible immune reaction against adenovirus or the transgene itself. The doses used here were 10–100 times lower than those previously reported to cause inflammation after injection.31 The relatively small amount of the virus may in part also explain why the expression of human MMP-13 mRNA was transient and could only be detected by the RT-PCR method. We chose to use the human MMP-13 in the experiments to be able to distinguish between endogenous production of murine MMP-13 and the expression of the adenovirally delivered gene. Because the homology between the human and murine MMP-13 is 86% and the sequence of the catalytic region differs by only one amino acid, it is reasonable to assume that the biological effects of human MMP-13 and murine MMP-13 are quite similar. Furthermore, the ability of this construct to mediate a collagenolytic response has been well established using melanoma cells.31
Our results showed that the synovium was the most abundant source of adenovirus mediated transgene expression, but expression of marker gene LacZ as well as MMP-13 was also detected in articular cartilage chondrocytes. As this was a consistent finding with all knees analysed it indicated an efficient transfer of adenovirus from synovial cavity to the growth plate. Similar observations have been made in rabbit and guinea pig models.28,32 As far as we know, the expression of adenovirally delivered transgene in the growth plate after intra-articular injection of adenovirus has not been reported previously, and is especially interesting because the interactions between the synovial cavity and the growth plate remain poorly understood. Based on our northern hybridisation data, some basal MMP-13 expression also occurs in the normal knee joint. Thus it is difficult to explain why a relatively small increase in the total expression of MMP-13 results in such a strongly reactive phenotype. In normal joints, MMP-13 expression has been observed in osteoblastic cells and the hypertrophic chondrocytes of the growth plate,33 whereas no MMP-13 expression has been reported in normal synovium. As the adenovirally delivered MMP-13 is mainly produced by synoviocytes, it is likely that this synovial activation of MMP-13 production has a key role in triggering the inflammatory cascade seen in our model.
In this model no massive cartilage erosion was seen, although superficial fibrillation of meniscal and articular cartilage was detected in some animals injected with RAdMMP-13. In a transgenic mouse model expressing an active mutant of human MMP-13 under type II collagen promoter, consistent expression of active human MMP-13 by articular chondrocytes leads to an osteoarthritic-like phenotype with marked cartilage degradation.21 Lack of marked cartilage destruction in our model is probably due to inability to achieve sufficiently strong and lasting expression of MMP-13 by using adenoviral gene transfer. The transient nature of inflammatory synovitis after MMP-13 expression may in part be due to the ability of joint tissues to “neutralise” the amount of MMP-13 produced in the joint after adenoviral gene transfer.34,35 It seems obvious that persistent long term MMP-13 activity in the joint is required for cartilage degradation, because intra-articular injections of recombinant MMP-13 protein also did not result in any marked cartilage erosion.36 Interestingly, also, in a well characterised Del1 mouse model for osteoarthritis, no production of MMP-13 could be seen at sites of superficial fibrillation of articular cartilage, which may imply that MMP-13 does not participate in the initiation of cartilage degradation.37 It has also been proposed that the specific receptor mediating MMP-13 internalisation has a role in cell signalling, and overexpression of MMP-13 might cause cellular malfunction through this mechanism.38 Thus the mechanism whereby intra-articular overexpression of MMP-13 induces synovial inflammation is probably not dependent on cartilage degradation. It is, however, possible that although no massive cartilage erosion was seen, the proteolytic activity of MMP-13 resulted in release of cleavage products of type II collagen or other extracellular matrix molecules into synovial fluid, which evoked the inflammatory response.
Recently, the role of MMPs in chemokine activation has been increasingly highlighted. For example, MMP-3 has been shown to be required for generation of macrophage chemoattractant and subsequent macrophage infiltration in a model of herniated disc resorption.6 It has been shown that local stimulation of collagenase expression, by lipopolysaccharide endotoxin injection into joints, induces inflammatory cytokines.39 Rat MMP-13 has been shown to release basic FGF from heparan sulphate proteoglycans, especially perlecan.40 Basic FGF stimulates fibroblast activation and matrix deposition, which leads to synovial fibrosis and hyperplasia. Besides controlling the bioavailability of different growth factors by releasing them from bound proteins, many MMPs also regulate the activity of several cytokines—for example, IL1β41 and TNFα.42 MMP-13 has been shown to be capable of cleaving and activating MCP-1 and stromal derived factor-1.43,44 The in vivo function of MMP-13 in chemokine shedding is, however, yet to be elucidated.
In our model intra-articular overproduction of MMP-13 led, nevertheless, to increased production of many inflammatory cytokines and chemokines with different expression profiles. Some up regulation of cytokine and chemokine production was also seen in the animals injected with RAdLacZ, reflecting the inflammatory response to the adenovirus. However, compared with the uninjected control group the increases in most cases remained relatively small and clearly distinguishable from the MMP-13 induced inflammatory reaction. The up regulation of MCP-1 and RANTES expression in the RAdMMP-13 treated group is in accordance with findings in human rheumatoid and reactive arthritis.45 RANTES production is also increased in osteoarthritic cartilage and has been speculated to have a role in chondrocyte activation and thus contribute to cartilage degradation.46 In the present model, the mRNA levels of IL1β—considered to be the major cytokine promoting the cartilage destruction in arthritides—were significantly up regulated at day 7 after the injections.
A surprising finding in the present model is the lack of responsiveness of TNFα expression despite the massive synovitis. Indeed, the biological proinflammatory effect of TNFα in inflammatory models has been suggested to result from post-transcriptional modifications and protein activation more than from transcriptional activation.47 In a recent study, gene expression profiling of murine collagen induced arthritis also failed to show regulation of TNFα expression.48
Inhibition of the proteolytic activity of MMPs provides an attractive option for a new anti-arthritic treatment. However, the substantial side effects—for example, joint contractures,49 have limited the use of broad acting MMP inhibitors for treatment of arthritis. Hence identification of the MMPs responsible for joint destruction and inflammation is essential in order to develop more targeted inhibitors. The results presented here provide evidence that MMP-13 may serve as a new target for anti-inflammatory treatment in synovitis.
Acknowledgments
The authors are grateful to Tuula Oivanen, Merja Lakkisto, and Marjo Hakkarainen and to the personnel of the animal facilities for expert technical assistance.
This study was supported by grants from the Academy of Finland (projects 52940 and 45996), Sigrid Jusélius Foundation, the Cancer Research Foundation of Finland, Turku University Central Hospital, and by a research contract with Finnish Life and Pension Insurance Companies. KJ is a recipient of a training grant from Turku Graduate School in Biomedical Sciences.