Article Text

Abstract
OBJECTIVE To elucidate factors involved in the arthritogenicity of bacterial cell walls.
METHODS For characterisation of an arthritogenic Eubacterium aerofaciens cell wall, peptidoglycan-polysaccharide (PG-PS) polymers were isolated by removing cell wall associated proteins (CWPs), PG and PS moieties were separated, and an attempt was made to de-O-acetylate PG-PS. The cell wall ofE limosum was used as a non-arthritogenic control. The chemical composition of these cell wall preparations was analysed by gas chromatography-mass spectrometry. Also, their ability to resist lysozyme degradation and to sustain experimental chronic arthritis was tested.
RESULTS The observations made with the cell wall of E aerofaciens, an anaerobic habitant of the human intestine, were compared with those reported from a pathogenicStreptococcus, showing that in both strains a complex consisting of PG-PS is required for the induction of chronic arthritis. The PS moiety most probably protects PG from enzyme degradation, allowing prolonged tissue persistence and leading to the chronic synovial inflammation. CWPs attached to PG-PS are not necessary for this function. O-Acetylation of PG, which is required for arthritogenicity of the streptococcal cell wall, seems not to be present in the arthritogenic E aerofaciens PG or only occurs to a small degree; attempts to de-O-acylate the E aerofaciens cell wall did not affect its arthritogenicity or lysozyme resistance.
CONCLUSION The results obtained indicate that the source of bacterial cell wall plays no part in the chemical or structural requirements for PG to induce chronic cell wall arthritis in the rats; the chemical structure of the PG moiety is decisive.
- intestinal flora
- rheumatoid arthritis
- peptidoglycan-polysaccharide
- cell wall induced arthritis
Statistics from Altmetric.com
The intestinal flora of patients with newly developed rheumatoid arthritis (RA) is significantly different from that of non-RA controls.1 ,2 This difference is primarily due to anaerobic bacterial species—namely, to Eubacterium aerofaciens.1 On the other hand, a single intraperitoneal (IP) injection of cell walls isolated from this Gram positive bacterium, a normal habitant of the human gut, induces a chronic arthritis in Lewis rats, whereas cell walls from a relatedE limosum are non-arthritogenic.3 ,4
The Gram positive bacterial cell wall skeleton mainly contains peptidoglycan (PG), polysaccharide (PS), and the cell wall associated proteins (CWPs) (fig 1). The CWPs in the Gram positive cell wall extend from the plasma membrane through the PG and PS layers to the cell surface and are covalently attached to the PG.5 ,6 They interact with the extracellular environment and contribute to the development of bacterial infections. CWPs ofStaphylococcus aureus interact with specific host molecules either by promoting attachment to the host tissues or by preventing the phagocytic clearance.7 CWPs ofMycobacterium leprae are the major stimulants of the cell mediated immunity,8 and CWPs ofM tuberculosis are known to play a part in establishing protective immunity.9 The M protein of the group A streptococcal CWPs is a major virulence factor with superantigenic activity.10 Group A streptococcal M protein has shared epitopes with the articular cartilage and synovium,11 and systemic administration of streptococcal cell wall induces a chronic, destructive arthritis in susceptible rat strains.12

Gram positive bacterial cell wall contains mainly peptidoglycan (PG), polysaccharide (PS), and cell wall associated proteins (CWPs). The CWPs outside the PS and PG layers can be removed by proteolytic enzymes, whereas the proteins inside the PG layer are difficult to remove without breaking the PG structure. PS binds covalently to the PG layer, and protects PG polymers from the degrading enzymes. PG consists of several layers (up to 70) of sugar chains composed of N-acetylglucosamine and N-acetylmuramic acid alternately linked each other. Short peptides bound to the N-acetylmuramic acid (muramyl peptides) connect the sugar chains. PG and the muramyl peptides possess multiple biological activities.
The bacterial cell wall PG-PS complex can initiate and sustain a chronic destructive inflammation.13 PG-PS isolated from human intestinal flora bacteria can induce chronic arthritis in rats.14 ,15 The arthritogenic ability of cell wall constituents is dependent on resistance to biodegradation and consequent persistence in the tissues.13 The PS moiety covalently bound to PG is essential for the PG-PS complexes to persist and induce chronic inflammation. Streptococcal PG-PS did not sustain chronic arthritis when the PS moiety was removed,16 and PS alone induced only mild oedema in the joints.17 Another structural basis for streptococcal arthritogenicity is the presence of an acetyl moiety at the C-6 hydroxyl group ofN-acetylmuramyl residues of PG, producing the corresponding 2,6-diacetylmuramyl PG (O-acetylated PG).18 Gonococcal O-acetylated PG is more resistant to enzyme degradation than non-O-acetylated PG,19 and subsequently persists in the tissues longer, leading to chronic arthritis.20 De-O-acylated group A streptococcal PG-PS induced less severe chronic arthritis than native PG-PS.21 In this study we aimed at defining the arthritogenic component(s) of the E aerofaciens cell wall to determine whether the same requirements are valid for a representative of the normal human gut flora and a pathogenic Streptococcus.
Methods
BACTERIA
Eubacterium aerofaciens ATCC 25986 was obtained from the Culture Collection, University of Gothenburg, Sweden, with a nomination CCUG 28087. E limosum ATCC 8486 was purchased from the American Type Culture Collection, Rockville, MD, USA. Both strains were grown overnight under strictly anaerobic conditions at 37°C in BBL Schaedler Broth (Becton Dickinson Microbiology Systems, Cockeysville, MD, USA) to the late logarithmic phase. They were characterised by 16S rDNA sequence analysis as described by Jalava et al.22
CRUDE CELL WALL PREPARATION
Bacterial crude cell walls were isolated as described previously.23 Briefly, the cells were broken with glass beads (diameter 0.45–0.50 mm) in an MSK cell homogeniser (B Braun Melsungen AG, Melsungen, Germany). The cell walls were collected by centrifugation, treated with deoxyribonuclease I (1 mg/g wet weight), ribonuclease A (10 mg/g wet weight), and trypsin (20 mg/g wet weight), washed, and sonicated in an ice bath for 75 minutes (Branson Sonifier, Smith Kline Co, Danbury, Conn., USA). The sonicated cell wall suspension was centrifuged at 10 000 × g, 4°C, for 20 minutes. The supernatant was centrifuged by ultracentrifugation (Sorvall Ultracentrifuge OTD65B, rotor 60 Ti, Du Pont Co, Wilmington, USA) at 100 000 × g, 4°C, for 60 minutes. The pellet containing the cell wall was suspended in water. For chemical analysis the cell walls were dialysed in membrane (MWCO 3500, Spectrum, CA, USA) against distilled water. For IP injection into rats the cell wall suspension was sterilised by heating at 90°C for 30 minutes.24 The sterility was checked by cultures on agar plates at 37°C and at room temperature under aerobic and anaerobic conditions; no bacterial growth was detected after two days of culture. The endotoxin tests by E-TOXATE (Sigma Chemical Co, MO, USA) were also found to be negative. These procedures were applied for all cell wall preparations used.
PG-PS POLYMERS
To obtain PG-PS polymers, the CWPs were removed; the crude cell walls were treated with proteolytic enzymes, protease (Subtilisin Carlsberg, 5 mg/g wet weight), proteinase K (2 mg/g wet weight), pepsin (20 mg/g wet weight), and papain (5 mg/g wet weight). All enzymes were purchased from Sigma Chemical Co, MO, USA. After lyophilisation the cell walls were extracted with chloroform-methanol (2:1 vol/vol, 40 mg/ml) at room temperature for four hours to remove lipoteichoic acid or glycosyl glycerides.16 After washing once with water and dialysis against distilled water, the cell walls were treated by 8 M lithium chloride at room temperature for one hour to remove any non-covalently bound CWPs.25 The cell walls were collected by ultracentrifugation and washed twice with distilled water. They were extracted with 0.1 M EDTA at room temperature for one hour to remove any divalent cation stabilised oligomeric proteins.26After ultracentrifugation and washing twice with distilled water the cell walls were suspended in acetone at room temperature for one hour and further mixed at 4°C for 30 hours to remove possible contaminating proteins and endotoxin.27 ,28 The cell walls were collected by ultracentrifugation, washed three times with distilled water, and dialysed against distilled water. The preparations thus obtained are PG-PS complexes as shown by gas chromatography-mass spectrometry (GC-MS) analysis.
PS AND SOLUBLE PG POLYMERS
PG and PS polymers were isolated from E aerofaciens ATCC 25986 as described previously forLactobacillus.29 Briefly, the lyophilised cell walls were extracted with 10% trichloroacetic acid at 60°C for four hours. The suspension was centrifuged at 38 800 ×g at 4°C for 30 minutes and washed once with distilled water. The supernatant contained the PS fraction, and the pellet the PG fraction. The soluble PG was obtained as described previously.24 The PG fraction was lyophilised and suspended in water at 10 mg/ml and sonicated (Branson Sonifier) for 180 minutes (18 × 10-minute cycles with breaks for cooling between the cycles) in an ice bath. The remaining insoluble PG fraction was removed by centrifugation at 10 000 ×g at 4°C for 30 minutes. The supernatant (soluble PG fraction) was collected and dialysed against distilled water for 48 hours.
DE-O-ACYLATION OF CELL WALL
The treatment of the cell walls for de-O-acylation was done as described previously.21 Briefly, the cell walls were lyophilised, suspended in 0.05 M sodium hydroxide at 37°C for four hours. After washing three times with distilled water, the cell walls were fragmented by sonication and subsequently separated by differential centrifugation as described above for the cell wall preparation.
GAS CHROMATOGRAPHY-MASS SPECTROMETRY (GC-MS)
The chemical composition of the cell wall preparations was analysed by GC-MS as described previously.30 The derivatised molecules were ionised by the electron impact method and analysed in the selected ion monitoring mode using single positive ions at a mass to charge ratio (m/z). Sugars were analysed as alditol acetate derivatives with the same ions as monitored by Gilbartet al,30 except m/z 289 for rhamnose. Fucose was used as internal standard for rhamnose, allose for glucose/galactose, andN-methyl-d-glucamine for muramic acid. Amino acids were analysed as butyl heptafluorobutyl derivatives with the same ions as used by Gilbartet al.30 l-Norleucine was used as internal standard for glycine, serine, alanine, leucine, cysteine, threonine, isoleucine, and valine; l-methionine for phenylalanine, glutamic acid/glutamine, proline, ornithine, lysine, aspartic acid/asparagine, and tyrosine; l-tryptophan for cystine, diaminopimelic acid, histidine, and arginine. One microlitre of the derivative was injected in the pulsed splitless mode and analysed by a gas chromatograph (model HP 5890A; Hewlett-Packard, Wilmington, DE) equipped with a fused silica capillary column (SE-54; Nordian Instruments, Helsinki) and coupled directly with a TRIO-1 mass spectrometer (VG Instruments, Manchester, UK). The oven temperature for sugars was started at 50°C, programmed to 270°C at a rate of 10°C/min, and held for one minute; finally, the oven was heated to 290°C for five minutes. For amino acids the oven temperature was started at 85°C, programmed to 280°C at a rate of 10°C/min, and held for one minute; finally, the oven was heated to 290°C for five minutes.
ANIMALS AND INDUCTION OF ARTHRITIS
All the animal experiments were done in concordance with the national and international laws and policies (Order No 1076/85, Finland; EEC Council Directive 86/609, OJL 358, Dec 1987). Inbred pathogen-free female Lewis rats weighing approximately 150 g were purchased from Harlan Sprague Dawley, Inc, Indianapolis, IN, USA. The animals were kept in Macrolon III cages with disposable filter tops (Scanbur, Denmark) and given autoclaved standard diet and water; all handling was performed in a laminar-flow hood. Arthritis was induced by a single IP injection of the cell wall preparations suspended in sterile phosphate buffered saline (PBS) on day zero. Control rats were injected with an equal volume of sterile PBS. To monitor development of arthritis, each limb was assigned a score of 0–4, based on the degree of erythema, oedema, painfulness, and functional disorder of the ankle and metatarsal joints (wrist and metacarpal joints) and judged by two independent observers as described previously.31 The arthritis score used in the “Results” is the sum of the score of each limb. The animals were observed for at least four weeks. Rats were killed under anaesthesia by Metofane (Pitman-Moore, Inc, Washington Crossing, NJ, USA).
SENSITIVITY TO LYSOZYME
The sensitivity of different cell wall preparations to lysozyme was tested as described previously.21 Suspensions of cell wall preparations (4 mg/ml) in 0.1 M sodium acetate buffer (pH 5.0) were incubated with lysozyme (400 μg/ml) at 37°C. The optical density of the suspension was measured at wavelength 560 nm after five and 24 hours' incubation.
STATISTICAL ANALYSIS
Differences of arthritis development between the experimental groups were analysed with the non-parametric Mann-Whitney U test. Differences were considered significant at p<0.05.
Results
IDENTIFICATION OF BACTERIA
The gene coding for 16S ribosomal RNA is generally used to identify bacterial strains. To confirm the identity of the bacterial strains used in this study the genes coding for this region were partially sequenced (320–452 base pairs). Both forE aerofaciens and for E limosum, a 100% identity with respective sequences in the GenBank data base was confirmed. In addition, a 72.6% identity between the strains of E aerofaciens andE limosum was obtained.
ANALYSIS OF CELL WALL PREPARATION BY GC-MS
All bacterial cell wall preparations were analysed by GC-MS before injection into the rats (table 1). The total amount of protein amino acids is 2.2% in the E aerofacienscrude cell wall, and less than 0.6% in its PG-PS polymer, in which the amount of individual protein amino acids is less than 0.1%. The amount of protein amino acids is 12.6% in the E limosum crude cell wall, and less than 0.7% in its PG-PS polymer. Both bacterial PSs contain rhamnose and glucose/galactose.E aerofaciens PS also containsN-acetylglucosamine.N-Acetylgalactosamine is present in theE limosum cell wall and missing in theE aerofaciens cell wall. PG of both cell walls contains the amino sugarsN-acetylmuramic acid andN-acetylglucosamine. ArthritogenicE aerofaciens has four PG amino acids: alanine, glutamic acid, lysine, and aspartic acid. Non-arthritogenicE limosum cell wall has five PG amino acids: alanine, glutamic acid, lysine, ornithine, and serine (table 1). These findings are in concert with the previously reported PG types—A4α for E aerofaciens and B2α forE limosum.23
Gas chromatography-mass spectrometry analysis of the cell wall preparations
INDUCTION OF ARTHRITIS
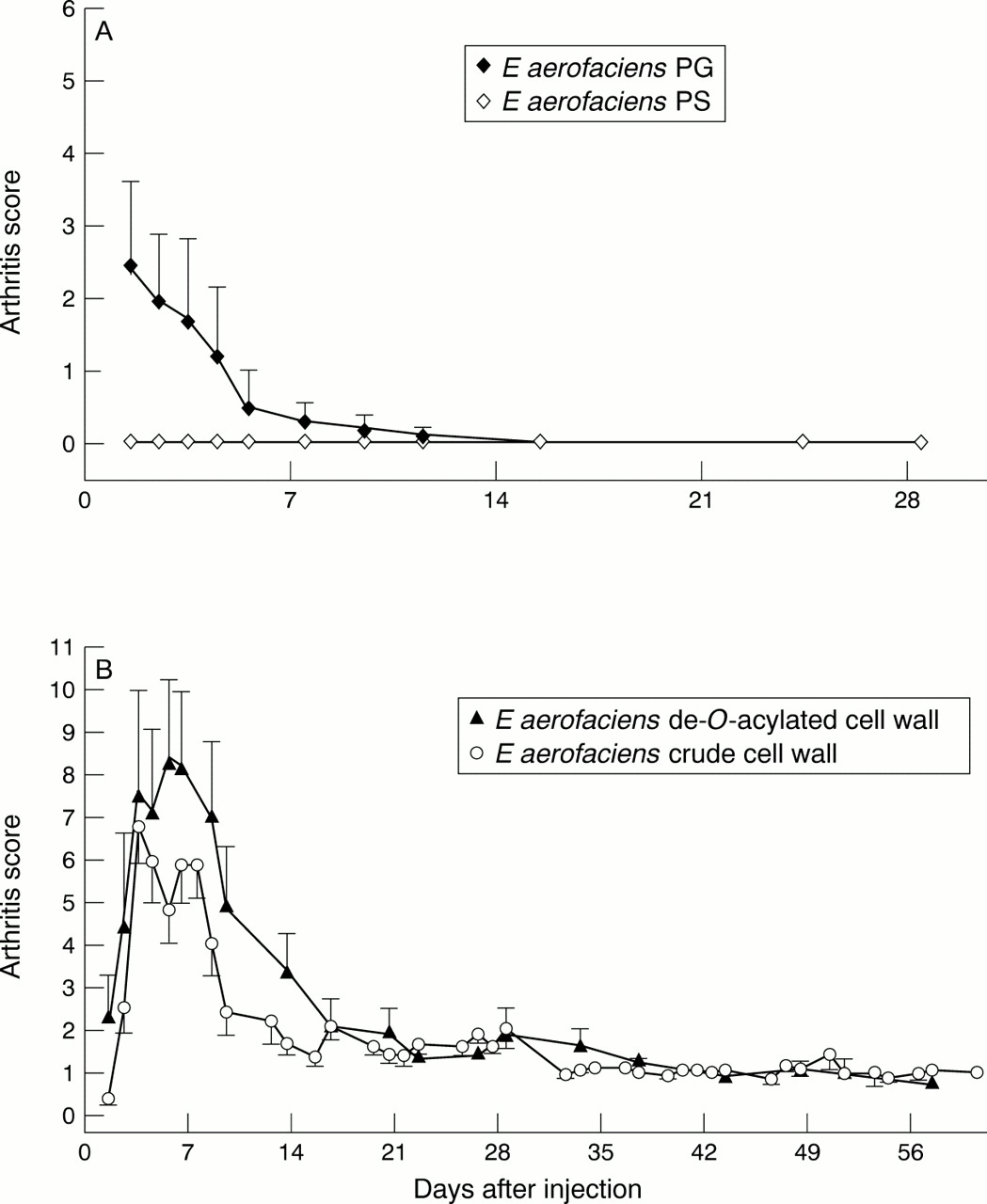
For induction of arthritis a single IP injection of the cell wall components was applied. E limosum PG-PS appeared non-arthritogenic, similar to the crude cell wall ofE limosum (fig 2B). A high dose (200 μg/g body weight) of PG-PS isolated from the arthritogenicE aerofaciens crude cell wall was toxic, leading to death within three days (data not shown here), probably owing to the high PG content. When a lower dose (150 μg/g body weight of rat) was used, E aerofaciens PG-PS caused a significantly more severe chronic arthritis than the equivalent dose of the crude cell wall (fig 2A), suggesting that the PG-PS complex possesses the structure decisive for arthritogenicity. However, after the PS moiety was removed, the PG alone induced only acute arthritis. Likewise, PS alone did not induce any arthritis (fig 3A). Further, attempts to de-O-acetylate theE aerofaciens cell wall did not affect its arthritogenicity (fig 3B). None of the rats injected with PBS alone developed arthritis.

(A) A single intraperitoneal (IP) injection of E aerofaciens peptidoglycan-polysaccharide (PG-PS; 150 μg/g body weight of rat) induced a more severe chronic arthritis than that induced with a crude cell wall (140 μg/g body weight of rat) (p<0.02 by non-parametric Mann-Whitney U test). Each black symbol represents a mean (SEM) of six rats. Each white symbol represents a mean (SEM) of 11 rats. (B) Rats injected with E limosum PG-PS (200 μg/g body weight of rat) IP or with the crude cell wall (200 μg/g body weight of rat) developed only mild acute arthritis. Each black symbol represents a mean (SEM) of eight rats. Each white symbol represents a mean (SEM) of six to 12 rats.
{kind=link}
{kind=link}
{kind=link}
(A) Rats injected with E aerofaciens peptidoglycan (PG; 33 μg/g body weight of rat) intraperitoneally (IP) deprived of polysaccharide (PS), developed only acute arthritis. Rats injected with E aerofaciens PS alone (122 μg/g body weight of rat) did not develop any arthritis. Each symbol represents a mean (SEM) of five rats. (B) Rats injected with E aerofaciens de-O-acylated cell wall (140 μg/g body weight of rat) IP developed arthritis similar to that seen after injection of the crude cell wall (140 μg/g body weight of rat). Each black symbol represents a mean (SEM) of six rats. Each white symbol represents a mean (SEM) of 11 rats.
DEGRADATION OF BACTERIAL CELL WALL PREPARATIONS BY LYSOZYME
E aerofaciens crude cell wall was found to be resistant to degradation by lysozyme; only 20.7% was degraded after 24 hours' treatment with this enzyme (table 2). In contrast,E limosum crude cell wall was susceptible to lysozyme, with 58.5% degradation after 24 hours. De-O-acetylation of E aerofaciens cell wall did not affect its susceptibility to lysozyme. E aerofaciens PG-PS showed the same degree of lysozyme resistance as the crude cell wall, whereas PG alone (after removal of PS) was rapidly degraded by lysozyme; 75.9% was already degraded after five hours (table 2).
Degradation of bacterial cell wall preparations by lysozyme
Discussion
It has been shown that synovial tissue of patients with RA may harbour traces of bacterial PG.32 The results presented in this paper, obtained with a normal bacterial habitant (E aerofaciens) of the human intestine, indicate that the PG moiety of the bacterial cell wall contains a structure decisive for induction of chronic arthritis in rats. They also show that the PG-PS complex is necessary for the arthritis induction; the PS moiety most probably protects PG from enzyme digestion through steric hindrance, allowing prolonged tissue persistence and leading to the chronic synovial inflammation. Similar conclusions have previously been reached in studies on streptococcal cell wall arthritis.16 Up to this point, our observations indicate that the same principles apply for the chemical structure, determining arthritogenicity here also, independently of the bacterial species used. However, in contrast with the streptococcal cell wall, in which the O-acetylation is known to be associated with the arthritogenicity and resistance against biodegradation,21 O-acetylation does not seem to be a prerequisite for the arthritogenicity or biodegradation of the E aerofaciens cell wall. Our findings suggest thatO-acetylation does not occur inE aerofaciens cell wall or only occurs to a small degree. The gonococcal cell wall seems to resemble the streptococcal cell wall in this respect becauseO-acetylation is required for its arthritogenicity.19 ,20
Lysozyme susceptibility is usually thought to be associated with arthritogenicity of the cell wall. Almost all bacterial strains studied for their ability to induce cell wall arthritis have been found to be lysozyme resistant; this includes species ofLactobacillus,Streptococcus,Bifidobacterium, andEubacterium.33 The only exception is the E limosum cell wall, which is non-arthritogenic and which has been reported to be lysozyme resistant.3 However, our results obtained by using the same strain of E limosum indicate both non-arthritogenicity and lysozyme sensitivity (table 2). Furthermore, because PG is degraded in vivo by other enzymes also (for example,N-acetylmuramyl-l-alanine amidase, N-acetylglucosaminidase, and others),19 ,34 ,35 it is unlikely that lysozyme resistance alone would be a decisive requirement for the ability of a cell wall to induce chronic arthritis.
It has so far remained undetermined whether CWPs attached to PG-PS contribute or are required for induction of chronic arthritis. Our results indicate that their presence is not necessary for arthritis induction. Removal of CWPs by proteolytic enzymes resulted in PG-PS which induced an even more severe chronic arthritis than that seen after injection of an equivalent amount of the crude cell wall (fig2A). These findings parallel those showing that genetic mutation of the streptococcal M protein attached to PG does not change arthritogenicity of the cell wall.36 Otherwise, results from GS-MS analyses (table 1) show very little, if any, further information about chemical or structural requirements for arthritogenicity. For instance, the presence of rhamnose does not correlate with this ability, even though such a correlation has previously been suggested.37
In a previous study on four Eubacteriumstrains it was shown that the chemical structure of PG present in the bacterial cell walls is decisive in determining arthritogenicity/non-arthritogenicity.23 The present results, based on characterisation of PG-PS complexes, further strengthen that conclusion by indicating that the source of a cell wall plays no part in the chemical or structural requirements of PG arthritogenicity; it does not matter whether the cell wall is isolated from a normal habitant of the human intestine or from a pathogenic strain of Streptococcus. The structure of PG is decisive. Studies with other bacterial species—for example, withClostridia andBifidobacteria, are needed to elucidate further the detailed structural determinants of the PG arthritogenicity.
Acknowledgments
We thank Leena Kivistö, Marja-Riitta Teräsjärvi, Karolina Karlson, and Heli Niittymäki for excellent technical assistance, Jari Jalava for 16S rDNA sequence analysis, and Yong Zhang for help in evaluating the arthritis.
References
Footnotes
This work was supported by EVO of Turku University Central Hospital.