Article Text

Abstract
Objective: Recent studies suggest that crystals of monosodium urate (MSU), deposited in joints of patients with acute gouty arthritis, activate the NACHT domain, leucine-rich repeat and pyrin domain-containing protein (NALP)3 inflammasome. In the present study we have investigated whether production of proinflammatory cytokines by crystals was exacerbated during costimulation with Toll-like receptor (TLR) ligands.
Methods: Mononuclear cells of 22 healthy donors were stimulated by various concentrations of MSU crystals in the absence or presence of lipopolysaccharide (LPS), Pam3Cys and flagellin. Production of tumour necrosis factor α (TNFα), interleukin (IL)1β and IL6, as well as the intracellular concentrations of proIL1β were measured by ELISA. mRNA transcripts of TNFα and IL1β were assessed by real-time PCR. Stimulation experiments were also performed with peripheral blood mononuclear cells (PBMCs) of one patient carrying a NALP3 mutation.
Results: MSU induced a moderate release of IL1β and IL6, but not of TNFα. Urate crystals amplified IL1β production stimulated by the TLR4 ligand LPS, while no synergy was apparent for IL6 production. In addition, no synergy between urate crystals and Pam3Cys (TLR2 ligand) or flagellin (TLR5 ligand) was apparent. The synergy between urate crystals and LPS was directed at the level of the NALP3 inflammasome, as it was present only when active IL1β was measured, but not at the level of IL1 mRNA or proIL1β. The synergy between LPS and MSU crystals ceased to exist in the presence of a caspase 1 inhibitor.
Conclusions: MSU crystals act in synergy with LPS for the induction of enhanced release of IL1β. Increased cleavage of proIL1β by urate-activated caspase 1 is proposed as the underlying mechanism.
Statistics from Altmetric.com
In gout, a chronic inflammatory disorder characterised by exacerbations and remissions, crystals of monosodium urate (MSU) deposited in joints and in synovial membranes lead to inflammation.1 Proinflammatory cytokines, such as interleukin (ΙL)1β, tumour necrosis factor α (TNFα) and IL8, are induced by MSU and orchestrate the inflammation, for instance by the recruitment of inflammatory cells.2
Recently, Martinon et al3 demonstrated that crystals of MSU activate the NACHT domain, leucine-rich repeat and pyrin domain-containing protein (NALP)3 (or cryopyrin) inflammasome. The inflammasomes are intracytoplasmic complexes of proteins that are activated by exogenous stimuli (eg, bacterial components) or endogenous substances (eg, urate). Activation of the inflammasome leads to conversion of procaspase 1 into active caspase 1 (or IL1β-converting enzyme (ICE)). The latter enzyme cleaves proIL1β into its active form, IL1β.4 5
Although the inflammasome concept is mainly based on over-expression studies in cell lines, such as the monocytic THP-1 cell line, a recent experimental study in mice revealed that coadministration of anti-IL1RI monoclonal antibody (mAb) or anti-IL1R antagonist (anakinra) with MSU inhibited peritoneal influx of neutrophils.6 Based on these findings, the authors managed to effectively cure symptoms of acute gouty arthritis in 10 patients with the administration of anakinra.6 However, all the above evidence does not readily explain the paroxysmal character of gout, as patients with gout usually have a chronic urate overload. Conceptually, two mechanisms could be proposed for the generation of exacerbations: either an acute overload of urate induces an inflammatory attack (eg, hyperuricemia induced during cancer chemotherapy); or trivial exposure to a secondary stimulus for cytokine production, in combination with urate overload, is necessary for triggering the attack. In this study we investigated the latter concept in vitro, focussing on a possible synergy between urate and well established ligands for Toll-like receptors (TLRs), the major cellular receptors of the innate host defence system.
MATERIALS AND METHODS
Volunteers
A total of 22 healthy volunteers (14 males, 8 females, mean (SD) age 32.8 (9.8) years) were asked to donate blood. After informed consent, 10 ml of venous blood was collected from the antecubital vein under sterile conditions. Production of cytokines by peripheral mononuclear cells from one patient with Muckle–Wells syndrome carrying the R488K mutation for NALP3 was also investigated.
Preparation of MSU crystals
MSU crystals were prepared according to the method described by Seegmiller et al.7 Briefly, a 0.03 M solution of MSU at a volume of 200 ml was prepared after diluting 1.0 g of uric acid (Sigma, St Louis, Missouri, USA) in 200 ml of sterile water containing 24 g of NaOH. The pH was adjusted to 7.2 after addition of HCl and the solution became pyrogen free after incubation for 6 h at 120°C. The solution was left to cool at room temperature and stored at 4°C. Crystals produced were 5–25 μm long. On each day of experiment, a small amount of the crystals were weighed under sterile conditions for application.
Stimulation of cells
Peripheral blood mononuclear cells (PBMCs) were isolated after gradient centrifugation of heparinised whole blood over Ficoll (Amersham Biosciences, Uppsala, Sweden) for 20 min at 1400 g. After three washings in ice-cold phosphate buffered saline pH 7.2 (B Braun Melsungen AG, Melsungen, Germany), PBMCs were counted in a Buerker chamber with trypan blue exclusion of dead cells. They were then diluted in RPMI 1640 enriched with 10 mM of l-glutamine, 10 μg/ml of gentamicin and 10 mM of pyruvate and suspended in wells of a 96-well plate (Greiner, Alphen aan den Rijn, The Netherlands). The final volume per well was 200 μl with a density of 2×106 cells/ml.
PBMCs were exposed to concentrations of MSU crystals ranging from 0.1 to 1000 μg/ml for 24 h at 37°C in 5% CO2. All experiments were run in duplicate, with cells of at least two donors per day of experiment, using cells stimulated by 1 ng/ml of lipopolysaccharide (LPS) of Escherichia coli as a positive control. At the end of incubation, plates were centrifuged and supernatants were collected and stored at −20°C until the cytokine assays were performed.
Potential LPS contamination of crystals was investigated by incubation of urate crystals for 30 min at 37°C in 5% CO2 with 2 μg/ml of polymyxin B, or incubation of cells with 100 ng/ml of LPS of Bartonella quintana, a potent TLR4 inhibitor,8 before exposing the cells to MSU crystals. No TLR4-dependent activity was detected (data not shown), demonstrating that our MSU crystal preparation was LPS free.
To investigate the potential capacity of MSU crystals to amplify the cytokine production induced by TLR stimulation, the following TLR ligands were added to PBMC cultures in addition to crystals: 0.1 and 10 ng/ml of E coli LPS (strain O55:B5, Sigma, extra-purified by the method described by Hirschfeld et al);9 0.1 and 1 μg/ml of Pam3Cys (EMC microcollections, Tübingen, Germany); and 0.5 and 5 μg/ml of flagellin of Salmonella typhimurium (InvivoGen, San Diego, California, USA).
To investigate the role of caspase 1, experiments were performed in the absence or presence of 5 μmol/litre of the caspase 1 inhibitor (ICE-i) Ac-Tyr-Val-Ala-Asp-2,6-dimethylbezoyloxymethylketone (YVAD). YVAD was purchased from Alexis Biochemicals (San Diego, California, USA) and solubilised in dimethylsulfoxide (DMSO) at 10 mg/ml.
To measure intracellular proIL1β, 200 μl of RPMI 1640 were added into each well after plate centrifugation and removal of supernatants. After three cycles of freeze/thaw to lyse the cells, the lysates were collected and kept frozen until assay.
Measurement of cytokines
Concentrations of TNFα, IL1β and IL6 in supernatants were estimated in duplicate by an enzyme immunoabsorbent assay (Pelikine; CLB, Amsterdam, The Netherlands). The lowest limits of detection were 40 pg/ml for TNFα; 20 pg/ml for IL1β and 16 pg/ml for IL6. ProIL1β was also estimated in duplicate by an enzyme immunoabsorbent assay (R&D Systems, Minneapolis, Minnesota, USA); the lowest detection limit was 234 pg/ml.
Quantitative PCR for mRNA expression of TNFα and IL1β
PBMCs were stimulated, as stated above, with or without 1 ng/ml of LPS of E coli and 10 and 100 μg/ml of MSU crystals, or a combination thereof. After 4 h of incubation at 37°C in 5% CO2 and plate centrifugation, the cell pellet was lysed with 400 μl of Trizol (Invitrogen, Karlsruhe, Germany) and kept at −80°C until extraction of RNA.
RNA was extracted with chloroform and gradient centrifugation for 15 min at 4°C and 12 000 g followed by treatment for 30 min at 37°C with 0.04 U/μl of DNAase (Ambion, Applied Biosystems, Nieuwerkerk a/d Ijssel, The Netherlands). RNA was recognised after 3% agarose gel electrophoresis and ethidium bromide staining. RNA (1.5 μg; Pharmacia Biotech photometer; Pharmacia Biotech, Piscataway, New Jersey, USA) was applied for the production of cDNA using 0.4 mM of dNTPs (Invitrogen), 1 U of RNA-sin (Invitrogen), 10 mM dithiothreitol (DTT) (Invitrogen) and 5× of the reverse transcriptase buffer in a Mastercycler 5330 apparatus using appropriate blanks (Eppendorf, Cambridge, UK). After an initial incubation step of 10 min at 65°C, 1 μU of reverse transcriptase (Invitrogen) was added followed by three steps: 10 min at 25°C, 50 min at 42°C and 15 min at 70°C. cDNA was kept at −80°C until assayed.
Expression of mRNA was tested by the iCycler system (BioRad, Philadelphia, Pennsylvania, USA) using 1 μl of cDNA, 0.1 mg/ml of sense and antisense primers, 3 mM of MgCl2 (Invitrogen), 0.25 mM of dNTPs (Invitrogen), 10× buffer and 1 mM of Taq polymerase with SYBR-Gr as a fluorochrome per reaction tube. Primer sequences were: for TNFα sense 5′-TGGCCCAGGCAGTCAGA-3′ and antisense 5′-GGTTTGCTACAACATGGGCTACA-3′, for IL1β sense 5′-GCCCTAAACAGATGAAGTGCT C-3′ and antisense 5′-GAACCAGCATCTTCCTCAG-3′ and for β2-microglobulin sense 5′-ATGAGTATGCCTGCCGTGTG-3′ and antisense 5′-CCAAATGCGGCATCTTCAAAC-3′. After an initial denaturation step for 10 min at 95°C, 34 cycles were performed. Each cycle consisted of three steps; denaturation for 30 s at 95°C; annealing for 30 s at 72°C; and elongation for 30 s at 95°C. Amplification was followed by a melting curve; appropriate blanks were applied. The PCR product was recognised after 3% agarose gel electrophoresis and ethidium bromide staining. Quantitative results were expressed as defined by the Pfaffl equation10 using the efficiency of a standard curve created with known cDNA.
Statistical analysis
Results were expressed as medians and 95% CIs. Percentage changes of the production of cytokines by the interaction of MSU crystals and LPS compared to single LPS without/with ICE inhibitor were estimated. Comparisons were performed by the Wilcoxon rank sum test. Any p value below 0.05 was considered significant.
RESULTS
MSU crystal induction of proinflammatory cytokines
The release of IL1β, IL6 and TNFα after stimulation of PBMCs of four healthy donors by various concentrations of MSU crystals are shown in fig 1. Concentrations of urate crystals below 10 μg/ml do not induce sizeable amounts of IL1β, IL6 and TNFα.

MSU crystals amplify LPS-induced production of IL1β
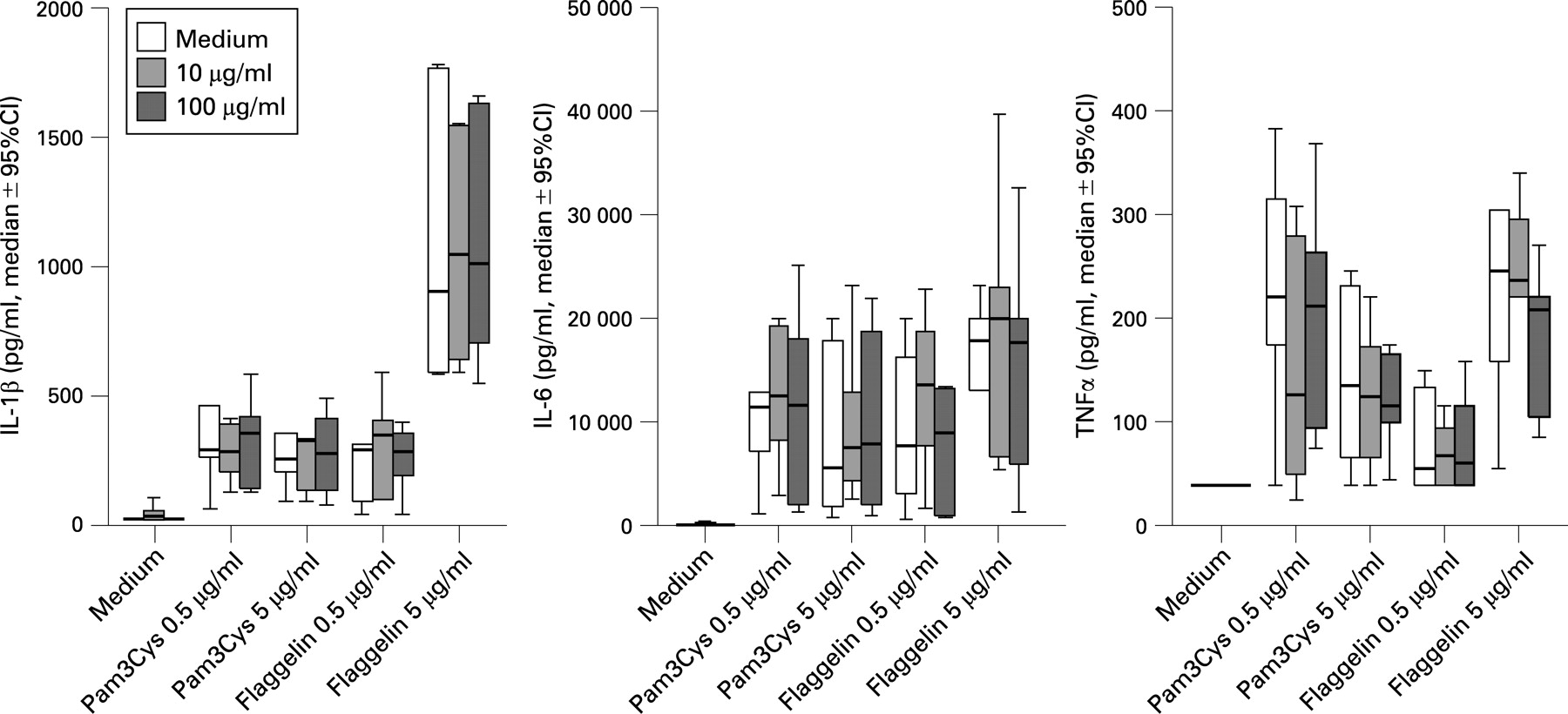
The next set of experiments dealt with the question whether MSU crystals would synergise with a secondary stimulus such as a TLR ligand for the induction of cytokines. Compared with MSU and LPS alone, the combination of LPS with MSU crystals induced a significant increase in the release of IL1β, arguing in favour of an interaction with the TLR4 pathway (fig 2). A similar synergy, although less strong, was also found for TNFα, but not for IL6 release (fig 2). By contrast, the addition of the TLR2 ligand Pam3Cys and the TLR5 ligand flagellin of S typhimurium did not change cytokine production (fig 3).
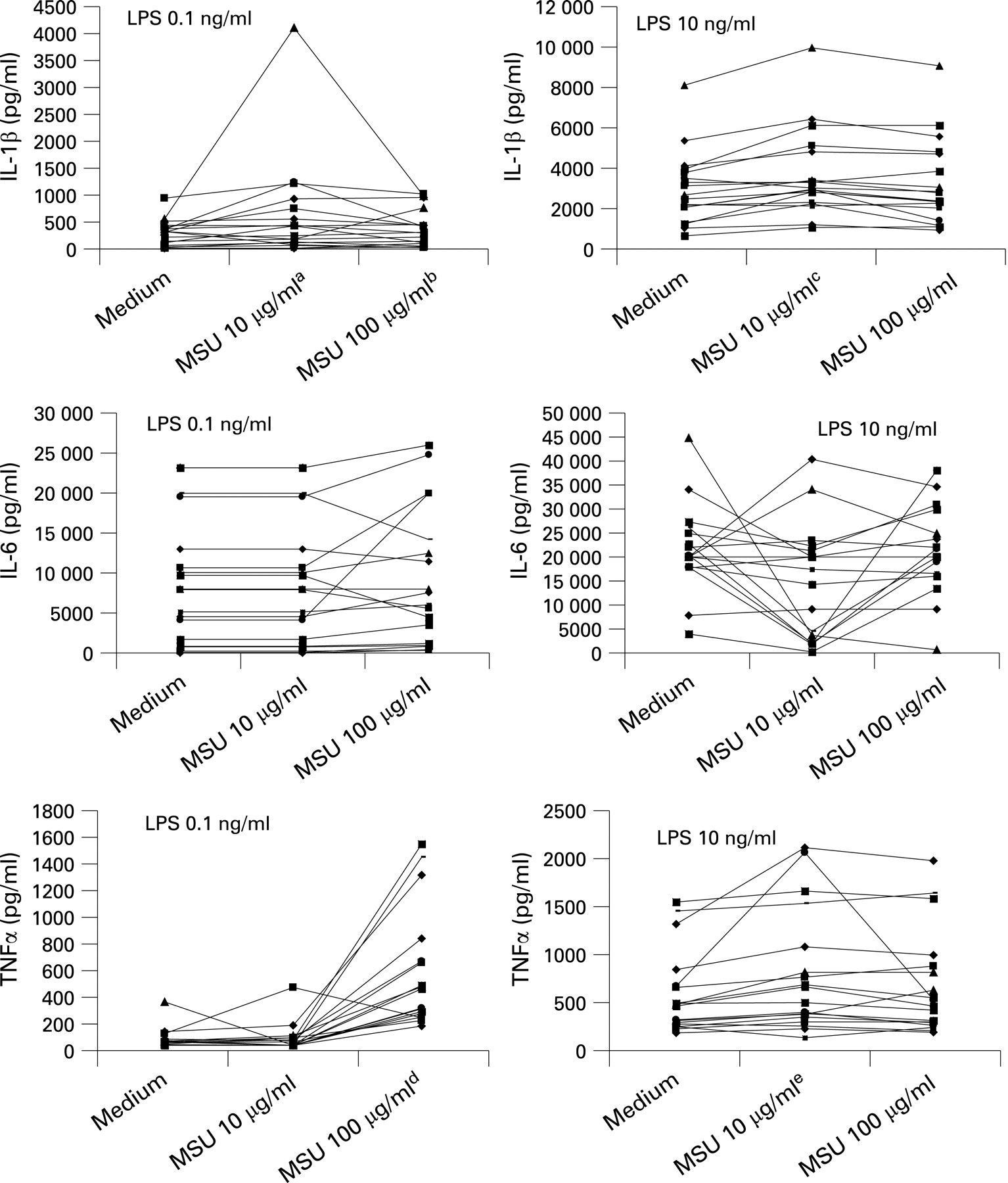
The role of caspase 1 activation for the synergism between MSU crystals and TLR4 ligation
We hypothesised that the synergy between LPS and urate crystals might be mediated through two possible mechanisms: either at the transcriptional level by an increased induction of the mRNA for proIL1β (and proIL1β production), or by enhancing the processing of proIL1 into mature IL1 through caspase 1 activation. The quantitative assessment of mRNA transcripts of IL1β and TNFα after stimulation with LPS and MSU crystals, as given in table 1, shows that the addition of MSU to LPS does not lead to a significant change in mRNA for either IL1 β or TNFα. Along the same line of reasoning, urate crystals did not lead to detectable production of proIL1β, and MSU crystals did not amplify the production of proIL1β induced by LPS (fig 4).

The issue of caspase 1 activation by MSU was addressed by exposing the cells to the caspase 1 inhibitor YVAD. As shown in fig 5, the increase of IL1β released by the interaction of LPS and MSU crystals ceased to exist in the presence of YVAD. The addition of YVAD did not affect the release of IL6 and TNFα.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cytokine release induce by MSU crystals in a patient with NALP3 mutations
Recently it has been proposed that MSU crystals induce caspase 1 activation through its recognition by NALP3,3 and individuals with Muckle–Wells syndrome bearing NALP3 mutations have autoinflammatory reactions through enhanced IL1β release.11 We investigated the response of a patient with Muckle–Wells syndrome to MSU crystals. PBMCs isolated from the patient with Muckle–Wells syndrome carrying the R488K mutation for NALP3 were cultured either with medium, LPS, MSU crystals, or the combination. Our hypothesis was that this patient’s cells would release more IL1 β when exposed to LPS, because of enhanced caspase 1 activity. For the same reason, we would expect to see little if any effect of urate crystals in the cells of the patient with NALP3, due to the already activated inflammasome. In line with this hypothesis, the unstimulated cultures of the patient’s PBMCs did not produce more IL1β than control cells. IL1β produced after stimulation with 0.1 ng/ml of LPS was 540 pg/ml (ie, considerably higher than that of the control (25 pg/ml)). IL1β released after stimulation with 10 μg/ml of MSU was 158 pg/ml compared to 20 pg/ml of the control. Costimulation with LPS did not enhance IL1β production yielding 660 pg/ml of IL1β. Respective intracellular proIL1β of PBMCs of the patient after stimulation with LPS, with urate crystals and after costimulation with LPS and urate crystals was 1490 pg/ml, 510 pg/ml and 1600 pg/ml.
DISCUSSION
In the present study we show that MSU crystals are able to induce production of IL1β and IL6 by PBMCs in vitro, albeit only at high concentrations (between 100 and 1000 μg/ml MSU). Interestingly, and in accordance with the concept that the inflammatory effects of MSU crystals are due to the activation of the inflammasome, there was little TNFα production induced by MSU crystals. Moreover, the concentrations of IL1β and IL6 induced by MSU were much lower than those achieved by stimulation with LPS and other TLR ligands. In this respect, our data are in accordance with those obtained by others using either primary monocytes or a human monocytic cell line.3 12
The modest capacity of MSU to induce cytokines, as well as the notion that many patients experience attacks of gout despite a chronic overload of uric acid, led us to the hypothesis that for a vigorous cytokine response in patients with gout, a second stimulus is needed. Indeed, we found that the cellular response to uric acid is strongly enhanced by LPS. The above observation suggests that in the patient with (asymptomatic) chronic hyperuricemia, attacks may be precipitated by trivial TLR4 stimuli, either exogenous, eg, LPS during transient subclinical endotoxinaemia, or by endogenous TLR4 agonists such as fibronectin, hyaluronic acid and heat-shock proteins.13 14
The low stimulation of cytokines by MSU alone and the strong synergy with LPS has additional implications. Thus, studies reporting strong induction of cytokines by MSU alone should be interpreted with caution if the LPS contamination was not excluded, as even minor LPS contamination could lead to erroneous results.
From our data derived from IL1β mRNA and proIL1β measurements, as well as the experiments employing caspase 1 inhibition, it can be concluded that the synergy between LPS and MSU reflects a two-stage process, in which LPS leads to activation of mRNA transcription and production of proIL1β, while exposure to MSU induce additional synergistic activation of caspase 1. Our results are consistent with those showing that stimulation of human monocytes with MSU crystals induced production of IL18. IL18 production induced by MSU crystals was inhibited after addition of a caspase 1 inhibitor,12 which is to be expected, as secretion of IL18 also requires cleavage of proIL18 by caspase 1.5 Interestingly, caspase 1 inhibition had only partial effects on the production of IL1β. The only partial effects of the caspase 1 inhibitor may be on the one hand due to the presence of certain amounts of proIL1β in the supernatant, and on the other due to the presence of other enzymes capable of processing proIL1β, such as proteinase-3 or elastases.15
Other mechanisms could be proposed to explain the synergistic effect between LPS and MSU crystals on the release of IL1β. These mechanisms are based on the chemical properties of MSU crystals, which have a negatively charged, highly reactive surface allowing them to bind to at least 25 different serum proteins.16 Among these are the binding of MSU crystals to the peroxisome proliferator-activated receptor γ,16 to CD14 receptor on the cell membrane of monocytes17 or to the TLR2 and TLR4 receptors.18 However, in all these studies MSU crystals have been applied at concentrations ranging between 500 and 1000 μg/ml, which are much greater than 10 μg/ml (ie, the concentration that showed optimal synergy with LPS in the present study and is relevant for the physiological concentrations found in the joints of patients with gout).19
The ex-vivo observations in the cells of the patient with Muckle–Wells syndrome and NALP3 mutations corroborate the in-vitro results. Patients with such mutations overproduce IL1β through chronic activation of the inflammasome, which is considered the mechanism behind the autoinflammatory nature of this syndrome.20 Our patient did exhibit overproduction of IL1β after stimulation with LPS, consistent with an increased cleavage of proIL1β induced by LPS. However, the exposure of his cells to urate crystals did not further augment the IL1β production, which is consistent with an already activated NALP3 inflammasome.
Median concentration of MSU crystals in the joints of patients with hyperuricemia is 7.5 crystals/400× power field.19 When considering the molecular weight of MSU, then the latter concentration assessed after microscopy is estimated to be close 50 μg/ml, ie, within the range of concentrations applied in the present study. These calculations render probable our hypothesis for a need of a second stimulus of TLR4 to elicit exacerbations of gout.
In conclusion, our study provides evidence that MSU crystals act in synergy with LPS, a ligand for TLR4, and thereby enhance release of IL1β. That synergy is mediated through activation of caspase 1 achieved when MSU crystals stimulate the NALP3 inflammasome. Through this mechanism, the released concentrations of IL1β are much higher than those achieved by MSU crystals stimulation alone, supporting the hypothesis that attacks of acute exacerbation of gout necessitate the presence of a secondary stimulus in addition to MSU crystals. Further research is required to confirm if this is the underlying mechanism responsible for the exacerbations of gout in patients with this disorder.
REFERENCES
Footnotes
Competing interests: None.
Ethics approval: Ethics approval was obtained.