Article Text

Abstract
Chronic inflammatory processes are based on a sustained and tightly regulated communication network among different cells types. This network comprises extracellular mediators such as cytokines, chemokines and matrix-degrading proteases, which orchestrate the participation of cells in the chronic inflammatory process. The mirrors of this outside communication world are intracellular transcription factor pathways, which shuttle information about inflammatory stimuli to the cell nucleus. This review examines the function of one key signal transduction pathway of inflammation—the p38 mitogen-activated protein kinases (p38MAPK). The signalling pathway is considered as crucial for the induction and maintenance of chronic inflammation, and its components thus emerge as interesting molecular targets of small molecule inhibitors for controlling inflammation. This review not only summarises the current knowledge of activation, regulation and function of the p38MAPK pathway but also examines the role of this pathway in clinical disease. It gives an overview of current evidence of p38MAPK activation in inflammatory arthritis and elaborates the key molecular determinants which contribute to p38MAPK activation in joint disease.
Statistics from Altmetric.com
Communication between plasma membrane receptors and the nucleus allows cells to respond to environmental danger signals. Rapid and adequate transduction of this information is critical for appropriate cell reactions and survival. Intracellular messengers act as couriers of information from the outside world to the genes inside the nucleus. Without such molecules, genes would be “blind”, like a person sitting in a room with no windows and doors, entirely deprived from external information. In real life, however, genes must be turned on quickly upon environmental changes.
One of these key sensors of cellular stress is the mitogen-activated protein kinase p38 (p38MAPK), which links inflammatory as well as environmental stress to transcription factors, which then bind to DNA and turn on the target genes.1 2 Transduction is accomplished by a cascade of activation steps involving sequential kinases linking the plasma membrane level with the transcription factor level (fig 1). p38MAPK is the most downstream signalling step before the transcription factor level and part of a greater family of proteins, the MAPKs, which share similar organisation structure. Whereas, p38MAPK and c-Jun N-terminal kinase (JNK) are mainly regulated by extracellular stress factors, the third pathway, extracellular signal-related kinases (ERK) is preferentially a target for mitogenic stimuli. In this review, we will focus on the p38MAPK cascade. Other papers have previously described the other two MAPK pathways, ERK und JNK, in rheumatic disease.3 4

INDUCTION OF THE P38MAPK PATHWAY
p38MAPK is not a single protein but comprises four different isoforms termed p38MAPKα, -β, -γ and -δ, which will be later discussed in more detail. Importantly, all isoforms are serine–threonine protein kinases that share the common phosphorylation motif TGY. Upon activation p38MAPK faces dual phosphorylation at threonine 180 and tyrosine 182.5 Inflammatory stimuli, such as lipopolysaccharide (LPS), tumour necrosis factor (TNF) and interleukin-1 (IL1) are the major inducers of p38MAPK. The fact that LPS induces p38MAPK led to the first description of this molecule, because p38MAPK was originally identified as an LPS-activated gene.5 Thus initial research focused on the clinical role of p38MAPK in septic shock, by virtue of its role in LPS-mediated induction of inflammatory cytokines such as TNF.5 6 TNF itself also activates p38MAPK by engaging type I TNF receptor.7 8 Downstream activation of p38MAPK then allows TNF to transduce its inflammatory message to the target organ—for example, the synovial membrane. TNF-mediated activation is also relevant in vivo because systemic TNF overexpression in mice leads to activation of p38MAPK in the inflamed joints.8 Moreover, this kinase not only integrates inflammatory stimuli but also signals heat stress, osmotic shock, ultraviolet light and cytotoxic chemicals.1 2 The activity of p38MAPK is tightly regulated by phosphatases such as mitogen-activated protein kinase phosphatase-1 (MKP-1), dephosphorylating p38MAPK.9 Interestingly, MKP-1 is strongly upregulated by glucocorticoids, suggesting that part of the anti-inflammatory properties of these drugs is based on p38MAPK inhibition.10 11 Regulation of MKP-1 by glucocorticoids has also been demonstrated in synovial fibroblasts from patients with rheumatoid arthritis (RA).12
In view of the presence of several different stress factors in the synovium, activation of the p38MAPK pathway in joints affected by RA is conceivable. Typically, proinflammatory cytokines are chronically increased in RA, which appears to be critical for p38MAPK activation in human RA.13
FIRST STEP: ACTIVATION OF MAP KINASE KINASE KINASES (MAPKKK)
Plasma membrane receptors are linked to the most upstream kinase (mitogen-activated protein kinase kinase kinase, MAPKKK or MEKK) in the by small GTPases of the ρ family, such as Ras, Rac, ρ or Cdc42.14 GTPases such as ρ and Ras appear to be involved in chronic synovial inflammation because both regulate synovial fibroblast proliferation.15 16 Moreover, inhibition of Ras also ameliorates adjuvant arthritis of rats.17 Ras is expressed in the synovium of RA; however, whether Ras is mutated in RA is controversial.18 The interaction between small GTPases and a MAPKKK leads to activation of a three-step kinase cascade that starts with the phosphorylation of a MAPKKK. The exact mechanism that selects a specific MAPKKK for phosphorylation of the p38MAPK pathway is unclear. In some cases the integration of signals is facilitated by the formation of signalling complexes that contain two to three kinases in series along with a structural protein. Some MEKKs such as MEKK1-4, preferentially activate downstream MAPKK, MKK4 and MKK7, which are important for the activation of JNK but not p38MAPK signalling, although in some cases MKK4 can phosphorylate p38MAPK.19–21 Among the four MEKKs, only MEKK2 protein expression is highly abundant in synovial tissue isolated from patients with RA, whereas cultivated synoviocytes express abundant MEKK1 and MEKK2 (fig 2).22 Importantly, MEKK1, MEKK2 (and TAK1) activate the JNK pathway in cultured synoviocytes as determined by in vitro kinase assay.22 One MEKK, that can only weakly affect p38MAPK, MEKK3 is only modestly expressed in the synovium. In contrast, ASK1 (also termed MEKK5), an MAPKKK that is stimulated by apoptotic stimuli is not only a strong inducing MAPKKK for p38MAPK23 but is also easily detected in the synovium of RA.22 TAK1 and MTK1 are other MAPKKKs that can induce the p38MAPK pathway by activating downstream MKK3 and MKK6.24 25 Protein and mRNA expression of TAK1 has been demonstrated in synovial tissue and cultured synoviocytes of patients with RA.22 Among the second family of MAPKKK, the mixed lineage kinases (MLKs), MLK3 and dual leucine zipper-bearing kinase (DLK)26 can activate p38MAPK. However, MLK3 is minimally expressed in the synovium of RA and for DLK it is unknown.

SECOND STEP: ACTIVATION OF MAPK KINASES (MAPKK)
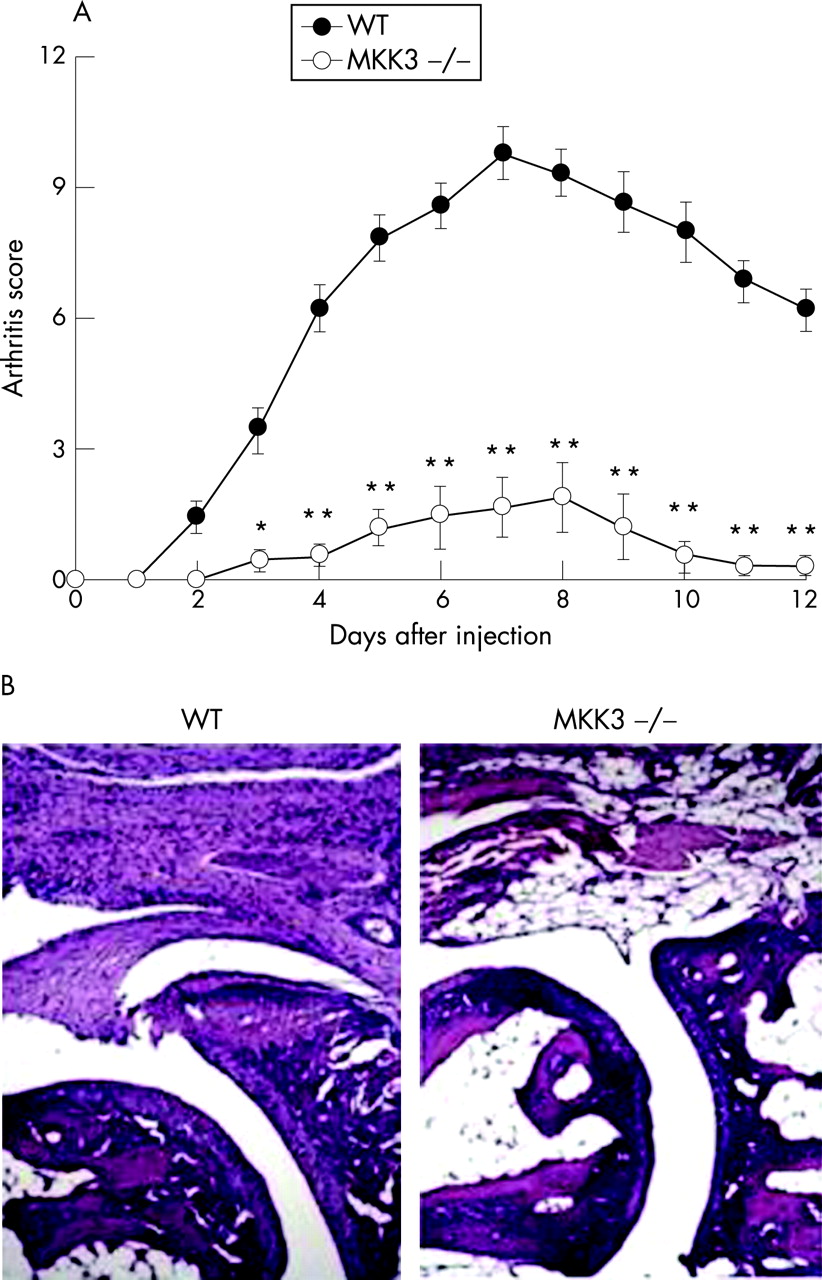
Activation of p38MAPK is accomplished by mitogen-activated protein kinases kinases (MKKs), that phosphorylate p38MAPK at amino acid residues 180 and182. MKKs are themselves subject to phosphorylation by upstream kinases (MAP3Ks), through threonine and serine residues at a specific domain of the MKK.27 Not all MKKs target p38MAPK but MKK3 and MKK6 are specifically important for the activation of p38MAPK. Studies from MKK3−/− mice and MKK6−/− mice have shown that each of them is required for full activation of p38MAPK in cells.28 MKK3 and MKK6 share an 82% amino acid homology and expression varies with respect to tissue- and cell type.29 30 Preferential activation of either activate MKK3 or MKK6 has been described: Thus osmotic stress leads to MKK6 activation, whereas p38MAPK activation in synovial fibroblasts is preferentially triggered by MKK3 when cells are exposed to proinflammatory cytokines like TNF or IL1.31 32 Of interest the MKK3−/− synoviocytes and mice responded normally to LPS challenge. These data suggest that a more favourable balance between suppression of host defence and synovial inflammation can be achieved by targeting upstream kinases. In line with an increased activation of p38MAPK in synovial tissue of RA, MKK3 and MKK6 are also activated in the joint of patients with RA.33 Phospho-MKK expression is found mainly in the synovial lining layer, which is the subcompartment in which p38MAPK activation also is preferentially found. In contrast, synovial tissue of osteoarthritis (OA) shows only very limited phosphorylation of MKK3/6, although both MKKs can be detected in non-phosphorylated forms. Cell types in the inflamed joint that exhibit activation of MKKs are synovial fibroblasts and macrophages.33 The type of MKK which ultimately leads to activation of p38MAPK and subsequent expression of proinflammatory mediators like IL1 or IL6 as well as matrix metalloproteinases depends on the initial trigger as well as on the cell types involved. Thus, TNF-mediated activation of p38MAPK in synovial fibroblasts is predominantly MKK3-dependent, whereas stimulation by IL1 or LPS does not absolutely require MKK3 and rather involves other mechanisms such as MKK6.31 Interestingly, passive induction of KBRN arthritis in MKK3-deficient mice results in significantly less disease severity, suggesting that MKK3-induced activation of p38MAPK is a critical step in chronic arthritis (fig 3).31
THIRD STEP: ACTIVATION OF THE P38 MAPK
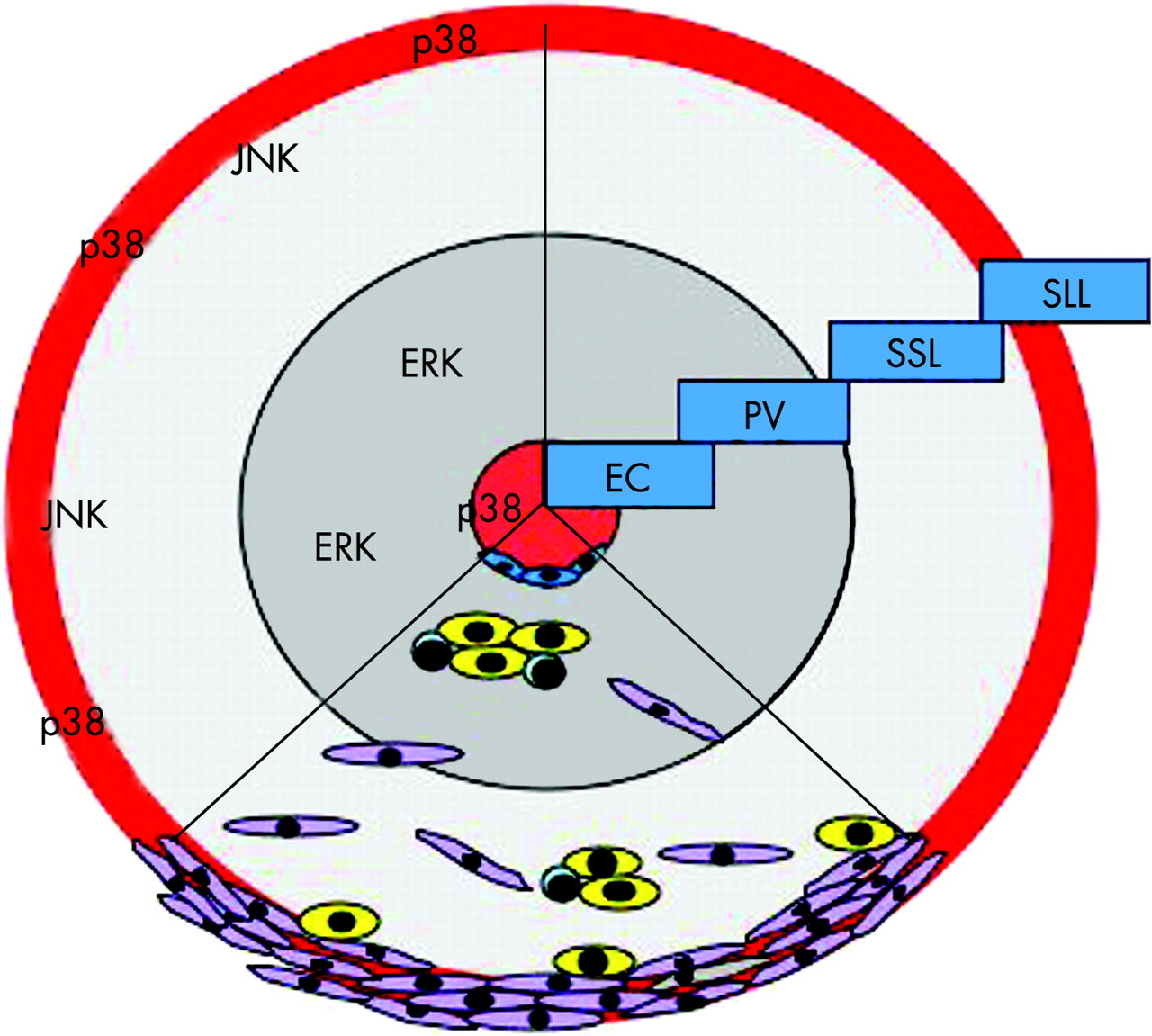
p38MAPK is strongly activated in the synovial membrane of RA, but not in synovial membranes of patients with degenerative joint disease.4 Roughly one-third of cells in the inflamed synovial membrane activate the p38MAPK pathway. Active p38MAPK is predominantly expressed in the endothelium of synovial microvessels and in the cells of the lining layer. These two regions are of significant importance when considering the necessity of transendothelial migration by blood-derived cells in the course of inflammation as well as the role of the lining layer in the formation of the destructive pannus (fig 4).

Apart from endothelial cells, macrophages and synovial fibroblasts are the major cell types exhibiting p38MAPK activation, which on the hand is only scarcely observed in lymphocytes.4 Similarly, activation of p38MAPK in the synovial tissue of animal models of arthritis is primarily associated with macrophages, synovial fibroblasts and endothelial cells. Osteoclasts also express activated p38MAPK at synovial invasion sites into bone in animal models of arthritis.34 TNF, IL1 and IL6 are potent inducers of p38MAPK in fibroblast-like synoviocytes.4
ISOFORMS OF P38MAPK
So far, four different isoforms of p38MAPK have been described and knowledge of differential expression and activation of these isoforms in disease is limited.35 36 Stimulating cells with LPS, leading to the discovery of the α-isoform of p38MAPK (also termed SAPK2a), is involved in the synthesis of the proinflammatory cytokines TNF and IL1.5 Later, the β-isoform of p38MAPK (also termed SAPK2b) has been described, which shares a 75% structural homology with p38MAPKα and is also activated by proinflammatory cytokines as well as environmental stress.37 Investigations in myocytes later unravelled a third isoform, p38MAPKγ (also termed SAPK3 or ERK6), which shares a 63% structural homology with p38MAPKα.38 39 And, finally, p38MAPKδ (also termed SAPK4) has been determined as a fourth isoform of p38MAPK, sharing 57% homology with p38MAPKα and which, like all other forms of p38MAPKs, responds to inflammatory and toxic environmental triggers.40 41
All four p38MAPK members share activation by some but not all upstream kinases (eg, MKK6)42–44 and common molecular targets of p38MAPKs like the transcription factor ATF2.35 37–44 Proinflammatory cytokines like TNF and IL1 have the ability to stimulate all four different isoforms.5 37–41 Thus, p38MAPK isoforms share several key functions. However, there are also differences among the isoforms with respect to (a) mode of activation; (b) regulation and inhibition; (c) substrate specificity; and (d) function, which underlie differences in the expression pattern of p38MAPK isoforms.
Selective activation is known for ρ GTPase, which activates the only the γ-isoform.14 As stated above ρ is important for the proliferative capacity of synovial cells.15 Activation of the δ-isoform has been described for L1-retrotransposable elements in fibroblasts of patients with RA.45 Activation can also differ among the cell type: thus, vascular endothelial growth factor (VEGF) activates α-isoform in endothelial cells; T-cell receptor signalling induces the α- but not the β-isoform in lymphocytes; and LPS enhances the α- and δ-isoform in granulocytes.46–48 Furthermore, there is selectivity in upstream kinases described. MKK3, for instance, does not activate the β-isoform.43 Finally, hypoxia mainly triggers activation of the α- and γ-isoforms, which is interesting since hypoxia-induced genes like HIF are abundantly expressed in the synovium.49
A second matter of isoform selectivity is the regulation and inhibition of p38MAPK. Thus, the most widespread class of synthetic p38MAPK inhibitors are pyridinyl imidazoles, which inhibit the p38MAPK α- and β-isoforms but not the γ- and δ-isoforms.37–40 These compounds are very effective in animal models of arthritis. Recent data indicate that arthritis is not attenuated in p38MAPKβ−/− mice, suggesting that activation of the α-isoform is most important.50 It is unclear whether compounds of different structures, such as triaminotriazine anilines, bicyclic pyrazoles, benzimidazoles or phenylmethanonenes have a different substrate pattern.51–54 Similarly, endogenous p38MAPK inhibitors such as MKP1 or MKP7 do not affect all isoforms but fail to inhibit p38MAPKγ as well as p38MAPKγ and -δ, respectively.55 56 Expression of such endogenous inhibitors might be critical in the physiological regulation of inflammation. Moreover, these molecules are substrates of glucocorticoids, thereby mediating part of their anti-inflammatory effects.10 Finally, also, specific inhibition of the β-isoform is known by histone deacetylase.57
Variable substrate specificity is another form of isoform-selective function. Whereas all isoforms affect ATF2, MAPKAP2, which is actually another kinase, is only phosphorylated by the α- and β-isoforms.35 37–40 42 43 MAPKAP2 activation has been demonstrated in synovial fibroblast-like cells.58 On the other hand, stathmin, a protein involved in the dynamics of microtubules, is a substrate for the γ- and δ-isoforms.59 Interestingly, stathmin expression is seen in blood vessels, where it potentially participates in endothelial migration and the organisation of smooth muscle cells.59 Furthermore, specific substrates for the γ-isoform are scaffolding proteins containing PDZ motifs, which are involved in membrane trafficking and sorting of receptors, ion channels and other signalling proteins for endothelial and smooth muscle cell.60
p38MAPK isoform selectivity can also be based on the differences in the function of the various isoforms and can vary depending on the cell lineage. Thus, p38MAPKα specifically induces the synthesis of proteases such as stromelysin (MMP3) or collagenase-1 (MMP1), which are important for mediating cartilage damage in RA.61 Moreover, the α-isoform can engage proteins in cell cycle arrest, such as p21, a CDK inhibitor. p38MAPKβ functions as a survival protein, inducing heat shock protein 70, and a potent anti-apoptotic factor induced in the synovial membrane of patients with RA.62 Maintaining cell survival is considered to be a key feature of p38MAPKβ activation.62 63 Conversely, p38MAPKγ is involved in myocyte differentiation, regulating proteins such as syntrophin, a synapse-associated protein.64 In contrast, p38MAPKδ is important for differentiation of keratinocytes as well as microtubule organisation, which might be important in the organisation of synovial microvessels.59 65
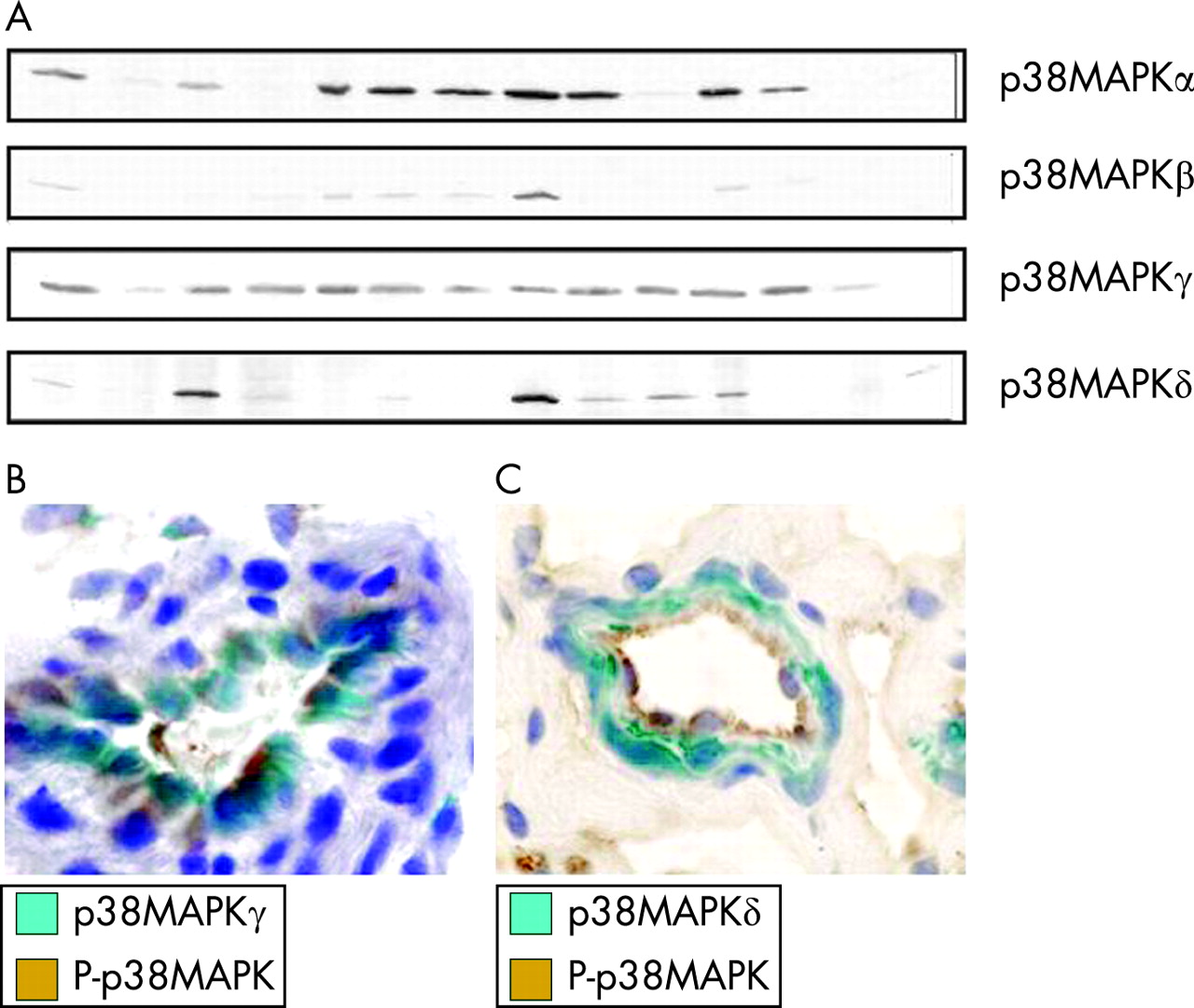
Recently, expression and activation of all four p38MAPK isoforms has been assessed in RA synovial tissue (fig 5). Among the four different isoforms of p38MAPK, the α- and γ-isoforms are the ones most abundantly expressed in the inflamed synovial tissue of patients with RA. As shown for the activated form of p38MAPK, expression of each of the isoforms except the δ-isoform is virtually confined to endothelium and the synovial lining layer, the metabolically most active compartment in the synovium.66 The sublining layer including the perivascular regions, which is located in between the lining and the synovial microvessels, is almost negative for each of the p38MAPK isoforms but does express other MAPK kinase families such as ERK and JNK, as demonstrated previously.4 Endothelial p38MAPK expression is dominated by the γ-isoform but also relevant amounts of p38MAPKα and -β are found. Only p38MAPKδ is not found in the endothelium but in myofibrocytes within the wall of synovial microvessels. Importantly, expression of p38MAPKδ has also been observed at sites of tissue destruction in patients with RA.45 Within the lining layer, the α- and γ-isoforms dominate in macrophages, the β- and γ-isoform in fibroblasts and the δ-isoform in granulocytes. Synovial lymphocytes express little or no p38MAPK. With respect to differential activation of p38MAPK isoforms it was observed that the α- and γ-isoforms are the ones that are predominantly activated in the inflamed tissue in vivo, whereas activation of p38MAPKβ is not common and phospho-p38MAPKδ is not found in the synovial membrane. Whether other sites such as the invasion front, where p38MAPKδ expression has been observed, or the bone marrow next to synovial inflammation, show a different activation pattern remains to be determined.
TARGET MOLECULES DOWNSTREAM OF P38MAPK
p38MAK is involved in regulation of Hsp27 and MAPKAP2 and several transcription factors including ATF2, STAT1, the Max/Myc complex, MEF2, ELK1 and indirectly CREB via activation of MSK1.35 37–40 42–44 Interaction with these transcription factors is a crucial determinant of cellular responses after p38MAPK activation. One of the key target molecules of p38MAPK is MAPKAP2, which can be phosphorylated by p38MAPKα and -β. MAPKAP2 is essential for the biosynthesis of TNF upon LPS challenge.67 MAPKAP2 activation has been observed in synovial fibroblasts upon stimulation of the p38MAPK pathway.58 Blockade of p38MAPK in arthritis models in vivo leads to reduced activation of the downstream target MAPKAP2, suggesting that MAPKAP2 is a key downstream target molecule of p38MAPK in inflammatory joint disease.68 Interestingly, MAPKAP2-deficient mice are resistant to collagen-induced arthritis.69
ROLE OF P38MAPK IN SYNOVIAL INFLAMMATION
The mechanisms of gene regulation are complex and can include both transcriptional and translational events as well as alterations in mRNA stability. One major group of target genes for p38MAPK activation are proinflammatory cytokines such as TNF, IL1 and IL6.5 6 Regulation of these inflammatory mediators is thus a major function of p38MAPK, which influences the balance of pro- and anti-inflammatory mechanisms and ultimately determines whether an environmental trigger leads to a physiological self-limited inflammatory response or causes chronic inflammatory disease. Selective blockade of TNF, IL1 or IL 6 can control disease by inhibiting joint inflammation and structural damage, indicating that an imbalanced cytokine production is a key event in RA.70 The regulatory role of p38MAPK on cytokine production as well as the role of p38MAPKs as important intracellular messengers of the same cytokines provide the rationale to develop specific synthetic inhibitors. Such compounds indeed have anti-inflammatory properties in experimental animal models of arthritis and have faced difficulties in early-phase clinical trials, mainly owing to toxicity.67 71–76 Inhibition of p38MAPK in animal models leads to reduced inflammation, which correlates with reduced expression of cytokines such as IL1, IL6 and RANKL.67 Upon inhibition of p38MAPK, formation of synovial inflammatory tissue is generally reduced and no disproportional effect on specific cell lineages was noted; rather, the quantity of synovial cellularity was significantly reduced. This points to a general inhibition of synovial inflammation by p38MAPK blockade rather than specific effects on certain cell types, and suggests that p38MAPK activation is a critical factor determining severity of synovial inflammation. In contrast to pharmacological inhibition of p38MAPK, which probably affects more than one isoform of p38MAPK, the selective genetic deletion of the β-isoform is not sufficient to inhibit experimental arthritis.50
Other functions of p38MAPK besides regulating proinflammatory cytokine expression probably also regulate synovitis. The role of p38MAPK in the control of cell cycle and apoptosis,77 the regulation of oxidative processes by inducing nitric oxide synthethase induction71 78 and neutrophil activation79 are examples of such activities. The relative contributions of each are not known (fig 6).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
ROLE OF P38MAPK IN CARTILAGE DAMAGE
Cartilage damage is a hallmark of RA. It is based on increased proteoglycan loss as well as attachment and invasion of inflammatory tissue into the cartilage, which leads to its structural disintegration. Production of matrix metalloproteinases (MMPs) by synovial tissue appears to be a key prerequisite for synovial tissue to invade and destroy cartilage.80 81 Synthesis of MMPs is regulated through multiple MAPK families, including p38MAPK, suggesting that blockade of p38MAPK might have structural benefit in arthritis.82 83
Indeed, data from animal models of arthritis suggest that p38MAPK activation might be especially important for the destructive features of arthritis since p38 inhibitors strongly reduce cartilage degradation.67 It is uncertain, whether this effect is directly through p38MAPK-dependent regulation of MMP expression or an indirect effect due to lower expression of proinflammatory cytokines, especially IL1, which is a key inducer of MMPs (fig 6).
ROLE OF P38MAPK IN INFLAMMATORY BONE LOSS
Inflammatory bone destruction is also a central component of RA, as evident from the formation of local bone erosions in RA, which lead to structural damage, changed joint architecture and loss of joint function. Bone damage in arthritis results from formation of osteoclasts, which are derived from monocyte precursors and resorb bone.84–86 Formation of osteoclasts is modulated by the stimulation of haematopoietic precursor cells with cytokines—that is, RANKL and TNFα, which results in p38MAPK phosphorylation. Activation of p38MAPK is thus important for osteoclast formation.87 88 This is in line with the dramatic reduction of mature osteoclasts and osteoclast precursors in the synovial tissue of experimental arthritis after p38MAPK blockade.67 As a consequence, tissue invasion into juxta-articular bone can be effectively blocked by the use of p38MAPK inhibitors. In contrast, deregulation of p38MAPK signalling, such as found it is found in CD44−/− mice when challenged with TNFα, leads to increased osteoclast formation and increased bone resorption.34 The resulting osteoclasts are increased in number and size owing to a decreased expression of MKP-1, which is a major regulatory molecule of the p38MAPK (fig 6).34
ROLE OF P38MAPK IN ENDOTHELIAL FUNCTION DURING INFLAMMATION
Activation of p38MAPK in synovial microvessels may reflect autocrine, paracrine or even endocrine activation by proinflammatory cytokines. VEGF also makes use of p38MAPK to communicate mitogenic stimuli to endothelial cells, which are essential for the synovial microvessel proliferation in the newly formed inflammatory tissue.89 Activation of p38MAPK is critical for several different functions of endothelial cells such as (a) chemoattraction; (b) vasodilatation; and (c) angiogenesis. Chemoattraction is a particularly critical event in synovial inflammation, since a majority of cells in the inflamed synovium migrate from the blood stream and have passed the endothelial barrier. p38MAPK regulates adhesion molecule expression, such as E-selectin and VCAM-1 on endothelial cells, which regulates rolling and adhesion of leucocytes on the endothelium before transmigrating to the inflamed tissue.90 91 Also, molecules involved in chemoattraction such as MCP-1 are regulated by p38MAPK.92 Vasodilatation is the second key response of the endothelium and depends on p38MAPK. Nitric oxide (NO) synthetase is a central determinant of vasodilatation through regulating the formation of NO and a target gene of p38MAPK. NO-induced vasodilatation of blood vessels enables increased blood flow within the inflamed tissue and facilitates the trafficking of leucocytes to these sites. Thirdly, p38MAPK is involved in the signalling of angiogenic stimuli as outlined above. Angiogenic effects mediated by p38MAPK depend at least in part of the activation of ETS1 transcription factor (fig 6).
SUMMARY
The “cytokine era” of rheumatic disease is closely linked to a more detailed insight into the signalling processes involved in these diseases. Without engaging these molecules, cytokines would be unable to exert their proinflammatory, regulatory, mitogenic or chemoattractive properties that participate in chronic inflammation. p38MAPK is one of three MAPK cascades which are key signalling components of inflammatory disease. Over the past years there has been greatly increased insight into the expression, regulation and pathophysiological function of the components of the p38MAPK signalling cascade in inflammatory joint disease. Current data suggest that stress kinases govern a variety of processes in the synovial tissue of the inflamed joint. These new insights are important to judge in the rational selection of therapeutic targets in order to inhibit inflammation and tissue destruction in rheumatic diseases.
REFERENCES
Footnotes
Competing interests: None declared.
Funding: Supported by the SFB423 of the Deutsche Furschungsgemein-Schaft.