Article Text

Abstract
Background: CD4+ T cell responses to the G1 domain of aggrecan in patients with ankylosing spondylitis (AS) were recently reported. Whether such an immune response can be seen in the CD8+ subpopulation has not yet been determined.
Objective: To determine if HLA-B27 restricted G1-specific CD8+ T cells are present in AS and to analyse immunodominant CD8+ T cell epitopes.
Methods: Peripheral blood mononuclear cells of 45 patients with AS were stimulated with overlapping 18-mer peptides covering the whole G1 protein. Results were compared with those for patients with rheumatoid arthritis (RA) and healthy controls. For epitope analysis, G1-specific interferon gamma positive (IFNγ+) T cells were isolated by magnetic activated cell sorting. After in vitro expansion, CD8+ T cells were restimulated with 14 subpools of G1 peptides. T cells responding to G1 peptide subpools were quantified by flow cytometry according to IFNγ secretion. Predicted peptides were subsequently confirmed by stimulation with single peptides.
Results: G1-specific CD8+ T cell responses were found in 29/45 (64%) patients with AS, 18/35 (51%) patients with RA, but not in healthy controls. Five CD8+ T cell epitopes were identified as immunodominant in five patients. However, the T cell response was not HLA-B27 restricted. Nonamer peptides with an HLA-B27 binding motif did not induce a T cell response.
Conclusion: A G1 peptide-specific CD8+ T cell response is present in AS but also in patients with RA. It does not seem to be HLA-B27 restricted. Whether such a response has a role in the pathogenesis of AS needs clarification.
- AA, amino acid
- AS, ankylosing spondylitis
- IFNγ, interferon γ
- IL, interleukin
- MACS, magnetic activated cell sorting
- PB, peripheral blood
- PBMCs, peripheral blood mononuclear cells
- RA, rheumatoid arthritis
- SEB, Staphylococcus enterotoxin B
- TNFα, tumour necrosis factor α
- CD8+ T cells
- aggrecan
- ankylosing spondylitis
- self antigens
Statistics from Altmetric.com
- AA, amino acid
- AS, ankylosing spondylitis
- IFNγ, interferon γ
- IL, interleukin
- MACS, magnetic activated cell sorting
- PB, peripheral blood
- PBMCs, peripheral blood mononuclear cells
- RA, rheumatoid arthritis
- SEB, Staphylococcus enterotoxin B
- TNFα, tumour necrosis factor α
Ankylosing spondylitis (AS) is a common chronic rheumatic disorder1 affecting more men than women and starts normally in the second decade of life. It has a strong association with HLA-B27.2 Mainly based on this HLA association, it has been postulated that AS is a T cell driven disease, but the pathogenesis remains unclear. The best known function of HLA-B27 is the binding of antigenic peptides and the presentation of such peptides to CD8+ T lymphocytes.3 Therefore, the main hypothesis advanced for the association between HLA-B27 and AS is the arthritogenic peptide theory. It states that some B27 subtype alleles, owing to their unique amino acid residues, bind a specific arthritogenic peptide(s) that is recognised by CD8+ T cells,4–7 which then causes a local immunopathology.8
Magnetic resonance imaging studies suggest the presence of osteitis at disease sites in AS and the hypothesis has been proposed that it occurs at the bone/cartilage interphase.9–11 Especially in the early phases of AS, mononuclear cells invade and erode the cartilage at different sites.9,12,13 Based on these findings, it has been proposed recently that the cartilage is the primary target of the immune response in AS.14–16 The aggrecan G1 domain has been implicated as a possible T cell autoantigen in AS and other rheumatic diseases, based on results from animal models and on studies in patients.17–20 Aggrecan, as part of the proteoglycans, forms a major component of the extracellular matrix of cartilage and helps to maintain the three dimensional network of stable cartilage.14,21 A specific CD4+ T cell response to peptides derived from the G1 domain of aggrecan has been found in animal models22,23 and also in patients with AS. We reported such a response in 60% of patients with AS against the whole G1 protein and against a pool of overlapping peptides derived from this G1 protein.24 A CD8 response to G1 derived peptides has not yet been investigated.
PATIENTS AND METHODS
Patients
Peripheral blood (PB) from 45 HLA-B27+ patients with AS, with a mean (SD) age of 32.5 (8) years and a mean (SD) disease duration of 6.7 (7) years, was collected in the outpatient clinic of the Charité Berlin, Campus Benjamin Franklin, Germany. All patients with AS fulfilled the modified 1984 New York criteria for a diagnosis of AS25 and had active disease, as indicated by the presence of at least two of the following three measures: inflammatory back pain, joint effusion, or an increased C reactive protein (CRP) level. All patients were receiving non-steroidal anti-inflammatory drugs. None of these patients received glucosteroids, disease modifying antirheumatic drugs or tumour necrosis factor α (TNFα) blockers at the time of the study or in the weeks before the analysis. Blood was also collected from 35 patients with rheumatoid arthritis (RA; mean (SD) age 52.4 (5) years, mean (SD) disease duration 5.2 (7) years) and from 17 healthy controls (mean (SD) age 38.7(6) years). Patients with RA and controls were not tested for HLA-B27. The patients with RA were treated with different disease modifying antirheumatic drugs and with prednisolone up to a maximal dose of 7.5 mg/day, but none of these patients were treated with TNFα blockers.
To analyse the fine specificity of G1-specific CD8+ T cells five of the 45 patients with AS were further investigated.
G1 derived peptides
G1 peptides were synthesised as described previously.26 Forty six overlapping 18-mer peptides overlapping by 10 amino acids (AAs), which cover all 375 AA residues of the G1 protein, were synthesised.
In vitro stimulation of CD8+ T cells by 18-mer G1 derived peptides
T cells were stimulated in vitro with G1 derived peptides as described before.27 Briefly, 1 ml whole heparinised peripheral blood was stimulated with the whole peptide pool of 46 overlapping 18-mer peptides (5 μg/ml, each peptide) derived from the G1 domain of aggrecan. As a negative control, cells were stimulated in the presence of anti-CD28 without antigen. As a positive control mononuclear cells were stimulated with 5 μg/ml Staphylococcus enterotoxin B (SEB). Brefeldin A (Sigma) was added for the last 4 hours of the stimulation.
Staining for T cell surface markers, intracellular cytokines, and analysis by flow cytometry
T cells were stained after in vitro stimulation as described previously.24,27 Briefly, cells were quadruply stained for CD8 surface markers, CD69 surface markers, and the intracellular cytokines interferon γ (IFNγ) and TNFα. All stainings were performed in FACS Permebealising Solution (Becton Dickinson, Heidelberg, Germany). The following antibodies were used: antihuman CD8 PerCP (clone Leu-3a), anti-CD69 PE (Leu-23) obtained from Becton Dickinson (Heidelberg, Germany), anti-TNFα (Hölzel Diagnostika, Köln, Germany) coupled to FITC (Sigma), and anti-IFNγ coupled to Cy5 (Amersham Pharmacia Biotech, Freiburg, Germany). Positive cells were subsequently quantified by flow cytometry using a FACS Calibur from Becton Dickinson (San Jose, CA) with CellQuest software. After gating on CD8+ T cells, only cytokine positive T cells which were also positive for the early activation surface antigen CD69 were counted.
CD8+ T cells were regarded as positive after antigen-specific stimulation as judged by the percentage of CD69/cytokine double positive cells if at least 0.02% (at least 30 cells) of the gated CD8+ T cells were positive without background staining (stimulation with anti-CD28 without antigen only).27,28 If the background staining was above 10 cells the percentage of T cells positive for intracellular cytokine staining had to be at least three times higher than the background staining to be accepted as positive. CD69 is an early T cell activation marker and is up regulated shortly after stimulation with specific antigens.29
Isolation of G1-specific T cells using IFNγ secretion assay
For isolation of G1 peptide-specific IFNγ+ T cells PB was stimulated in vitro with 5 μg/ml whole G1 peptides in the presence of anti-CD28 (Immunotech, Marseille, France; 1 μg/ml) for 6–8 hours. Live G1 peptide-specific T cells were isolated using the IFNγ secretion assay kit from Miltenyi Biotec (Bergisch-Gladbach, Germany), according to the manufacturer’s instruction, and magnetic activated cell sorting (MACS; Miltenyi, Bergisch Gladbach, Germany) as has been described previously in detail.30 Isolated cells were additionally stained for CD8 surface expression using anti-CD8-Cy5 (Amersham Pharmacia Biotech, Freiburg, Germany) according to the manufacturer’s instructions. Propidium iodide was used at a concentration of 1 μg/ml to exclude dead cells from the analysis of antigen-specific IFNγ secreting cells and avoid false positive signals. Cell samples were analysed on a FACS Calibur flow cytometer (Becton Dickinson, San Jose, California) using CellQuest software (Becton Dickinson). Isolated cells were washed twice in medium and then cultured in medium with 10% autologous serum in one well of a 96 well plate.
Cytometric evaluation of G1 peptide-specific CD8+ T cells expanded in vitro
To evaluate fine specificities, T cells, which were expanded in vitro by adding interleukin (IL) 2 twice a week, were restimulated with G1 peptides, which were used at a final concentration of 5 μg/ml (each peptide). Subpools of peptides were used as described previously by Kern et al.31 Briefly, subpools of peptides contained 5–8 peptides and were created in such a way that each peptide of the entire G1 protein was present in at least two different subpools of peptides. Immunostimulatory peptides were then predicted if two subpools of peptides were able to induce reactive cytokine secretion among G1 peptide-specific CD8+ T cells. This was then confirmed by stimulation with the predicted peptides and control peptides.
After 3–4 weeks of culture without IL2 for at least 5 days, G1 peptide-specific T cells were either further expanded in vitro or CD8+ T cells were isolated by MACS and analysed cytometrically for their fine specificity. Cytometric analysis was carried out by restimulation with subpools of peptides in the presence of autologous peripheral blood mononuclear cells (PBMCs), which were labelled with carboxyfluorescein diacetate succinimidyl ester (CFDA-SE; Molecular Probes, Eugene, Oregon) before restimulation. PBMCs were washed and resuspended in phosphate buffered saline (PBS) to a final density of 2×107/ml. A 0.5 μM staining solution of CFDA-SE (Molecular Probes) in PBS was prepared from a 5 mM stock. The cell suspension was added to the staining solution in a 1:1 ratio. After incubation for 4 minutes at room temperature, cells were washed twice and finally resuspended in medium with 10% autologous serum. PBMCs and T cells were restimulated in a ratio of 5:1 with at least 2×105 T cells for 6 hours. For the last 4 hours the secretion inhibitor Brefeldin A (Sigma) was added. Subsequently, cells were harvested, fixed, and analysed by fluorescence activated cell sorting.
Identification of single immunodominant CD8+ T cell epitopes
Based on the T cell response to the different peptide subpools of peptides of the G1 protein, predicted peptides were subsequently confirmed by stimulation with single peptides.
Nonamer peptides carrying an HLA-B27 binding motif with arginine (R) at position 2 and which were part of the immunodominant 18-mer peptides were also tested for HLA-B27 restriction of this CD8+ T cell response. In parallel, CD4+ T cells response to the same antigens was analysed.
The same stimulation was repeated in the presence of an anti-HLA-B27 antibody (DPC Biermann GmbH, Bad Nauheim, Germany) in one patient (patient 1) in order to define whether the T cell response is HLA-B27 restricted. For this, PB T cells were incubated with the anti-HLA-B27 antibody (40 μg/ml) for 1 hour before antigen was added.
Statistics
A χ2 test was used to compare frequency in different groups; differences were considered to be significant for a two tailed p value of <0.05.
RESULTS
Antigen-specific cytokine secretion by CD8+ T cells in AS and RA
A G1 peptide-specific CD8+ T cell immune response was seen in 64% of patients with AS (fig 1). The median (25th to 75th centile) of IFNγ/CD69+ CD8+ T cells for all patients was 0.07% (0.015–0.82%) compared with stimulation without antigen 0.005% (0–0.01%). The median of IFNγ/CD69+ CD8+ T cells for all patients after SEB stimulation used as a positive control was 4.41% (2.64–6.92%). However, there was also an increased frequency of IFNγ positive, PB derived CD8+ T cells in PB specific for the aggrecan G1 peptides in RA: 51% of the patients with RA responded. None of the healthy controls showed CD8+ T cell response to G1 peptides. The difference between AS or RA and healthy controls was significant (p<0.05), but the small difference between AS and RA was not significant.
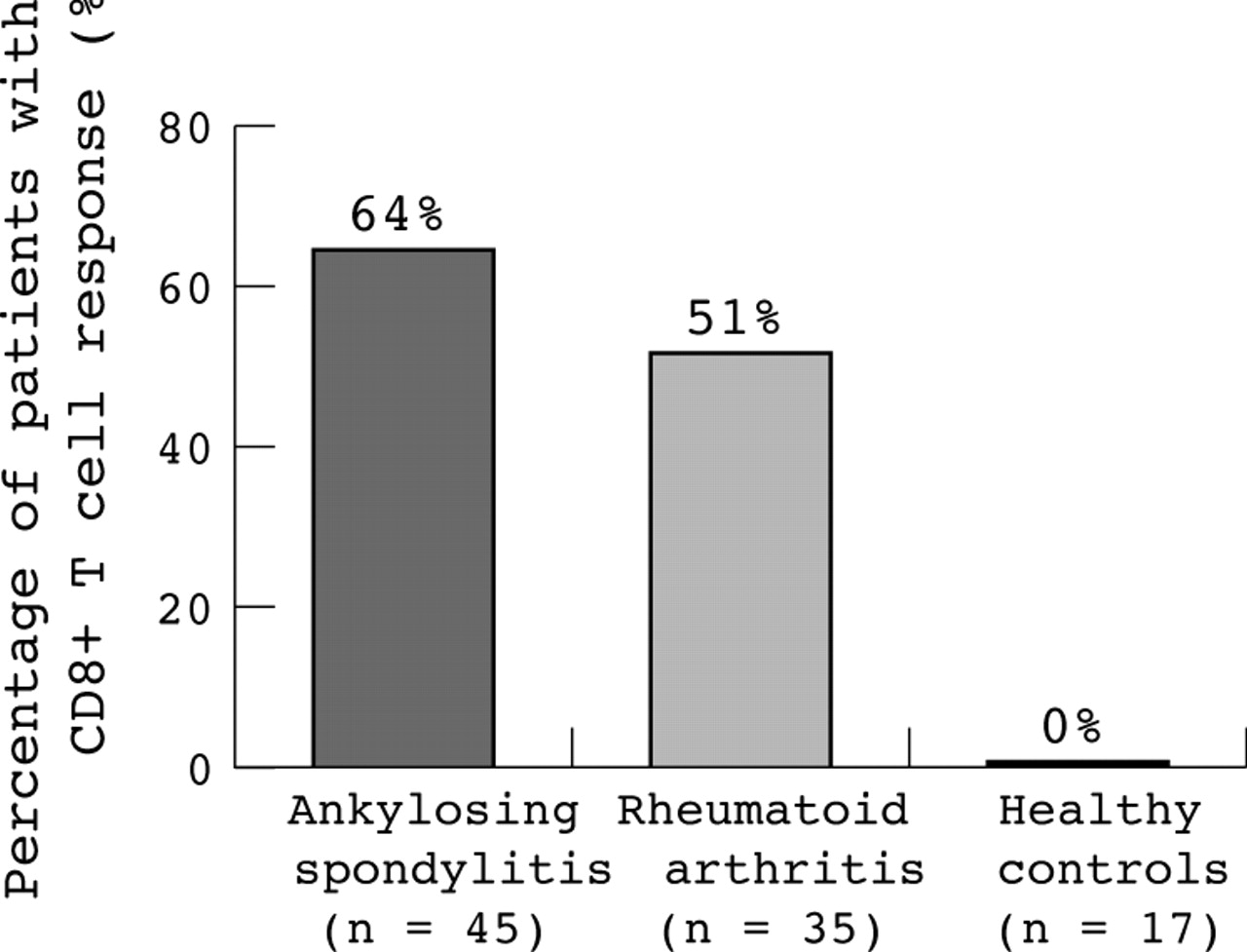
Percentage of patients with AS (64%), with RA (51%), or healthy controls (0%) responding to in vitro stimulation with the whole peptide pool of overlapping G1 peptides of the proteoglycan aggrecan. Response was measured by IFNγ production of CD8+ T cells after antigen-specific stimulation in comparison with stimulation without antigen. For more details see “Patients and methods” section. Analysis was done with whole PB.
Figure 2 shows an example of antigen-specific IFNγ secretion of PB CD8+ T cells of a patient with AS in response to stimulation with the whole peptide pool containing G1 peptides. The stimulation with the whole peptide pool was repeated in five patients altogether with reproducible results (data not shown). Examples of the reproducibility of our results at the single peptide level are shown below (see figs 6 and 7).

Example of an antigen-specific response to the whole peptide pool of aggrecan derived G1 peptides compared with stimulation without antigen in a patient with AS. After staining for T cell surface markers and intracellular cytokines a gate for CD8+ T cells was set. The percentage of IFNγ/CD69 double positive cells of the CD8+ T cell subpopulation is indicated. Stimulation with SEB was used as a positive control.
We also performed intracellular TNFα staining after stimulation with the whole peptide pool. However, owing to higher background staining than with intracellular IFNγ staining these data are not presented further.
Enrichment of G1-specific CD8+ T cells in patients with AS, and characterisation of immunodominant G1 epitopes
Because of the low frequency of G1-specific T cells in peripheral blood, G1 peptide-specific T cells of five patients were enriched by the IFNγ secretion assay for further epitope analysis (table 1). Figure 3 shows an example of one of these five patients. A frequency of 2.8% of G1 peptide-specific IFNγ+ CD8 T cells was detected after 6 hours of stimulation before MACS sorting using staining of the surface IFNγ. Surface IFNγ staining results constantly in a higher percentage of positive cells than with intracellular IFNγ staining, probably because the latter approach implies fixation and permeabilisation of the cells. These cells could then be nicely enriched by MACS sorting for IFNγ+ T cells, resulting in 60.5% antigen-specific CD8+ T cells for the patient shown (range 42.7–60.5% for all five patients) as judged by positive IFNγ staining.
Response of aggrecan G1-specific CD8+ T cell lines after stimulation with subpools of peptides or with single peptides in five patients with AS
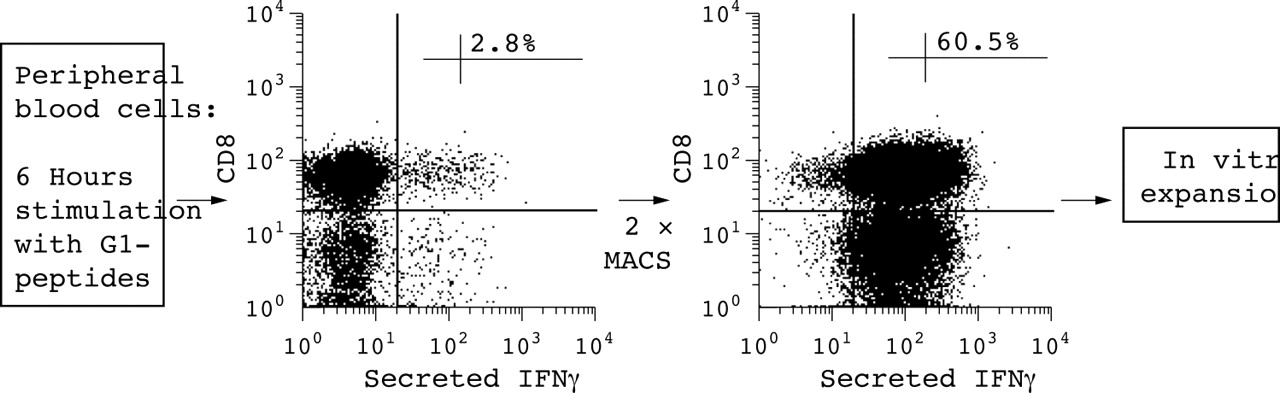
Example of an antigen-specific IFNγ secretion assay after in vitro stimulation with aggrecan G1 peptides compared with stimulation without antigen in a patient with AS. After stimulation, IFNγ+ T cells were isolated twice by MACS. For more details see “Patients and methods” section.
Subsequently, these cells were cultured and expanded for 4 weeks in the presence of IL2 and then used for the identification of immunodominant peptides. The presence of each peptide in two subpools of peptides makes it possible to predict positive peptides, which were subsequently tested. Figure 4A shows an example of this. In this patient (patient 1), four positive subpools of peptides (pool 1, 8, 10, and 11) out of 14 tested were found to be stimulatory for the G1-specific T cell line. Based on the matrix as shown in fig 4A, four predicted single peptides were further tested using this T cell line revealing peptides 16 and 17 as stimulatory in the same patient (fig 4B). When this approach was used one to two immunodominant peptides could be identified in each of the five patients, which were, however, only partly the same. Table 1 shows all the positive subpools of peptides and positive single peptides detected in the five patients.

(A) Example of a patient with AS (patient 1) with an antigen-specific CD8+ T cell response to the subpools of peptides (pools 1, 8, 10, 11) derived from the G1 domain of aggrecan compared with stimulation without antigen. After staining for T cell surface markers and intracellular cytokines a gate for CD8+ T cells was set. The percentage of IFNγ/CD69 double positive cells of the CD8+ T cell subpopulation is indicated. Peptide subpools 1, 8, 10, and 11 were positive, indicating that the crossing peptides 9, 16, 17, and 24 might be stimulatory peptides. (B) Example of a patient with AS (patient 1) with an antigen (Ag)-specific CD8+ T cell response to single peptides 16 and 17, but not to peptides 9 and 24 and control peptide, derived from the G1 domain of aggrecan compared with stimulation without Ag. After staining for T cell surface markers and intracellular cytokines a gate for CD8+ T cells was set. The percentage of IFNγ/CD69 double positive cells of the CD8+ T cell subpopulation is indicated.
We also investigated if the antigen-specific CD8+ T cell response to the 18-mer single peptides 8, 9, and 17 is mediated by a nonamer peptide that is part of the positive 18-mer peptide and carries an HLA-B27 binding motif with an arginine at position 2. No antigen-specific CD8+ T cell response could be detected after stimulation with these nonamer peptides. Figure 5 illustrates this result for stimulation with the nonamer peptide derived from peptide 17.

Example of a patient with AS with an antigen-specific CD8+ T cell response to 18-mer single peptide 17, but not to a nonamer peptide that is part of this 18-mer peptide, derived from the G1 domain of aggrecan, compared with stimulation without antigen (Ag).
After identification of these immunodominant T cell epitopes, PB was taken again from these five patients. When PB was directly stimulated with single peptides of G1 peptide CD8+ T cell epitopes, a clear immune response was found in the CD8+ T cell subpopulation but not in the CD4+ T cell subpopulation, also confirming the specificity of the T cell response. Figure 6 shows the example of patient 1.

Example of 18-mer peptides derived from the G1 domain of aggrecan inducing an antigen-specific CD8+ T cell response but not CD4+ T cell response compared with stimulation without antigen (Ag) in patient 1. After staining for T cell surface markers and intracellular cytokines a gate for CD8+ or CD4+ T cells was set. The percentage of IFNγ/CD69 double positive cells of the CD8+ or CD4+ T cell subpopulation is indicated.
Blockade with an anti-HLA-B27 antibody did not restrict the peripheral CD8+ T cell response when tested in patient 1, indicating that this antigen-specific CD8+ T cell response was not HLA-B27 restricted (fig 7). As a positive control this anti-HLA-B27 antibody was shown to block a CD8+ T cell response of an HLA-B27 restricted T cell line specific for an immunodominant Epstein-Barr virus peptide (EBNA 258–266) (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Antigen-specific CD8+ T cell response to 18-mer single peptides (peptides 16,17) which cannot be blocked by an anti-HLA B27 antibody in patient 1. T cells were taken from PB.
DISCUSSION
In this study we investigated the CD8+ T cell response to the G1 domain of the proteoglycan aggrecan in patients with AS in more detail. As for CD4+ T cells, CD8+ T cells specific for this protein could be detected in AS in more than 50% of patients but not in healthy controls. However, such an immune response is also present in patients with RA, indicating that aggrecan-specific CD8+ T cells are not confined to AS. In view of the observed CD4+ T cell responses in RA,32 osteoarthritis,33 and juvenile idiopathic arthritis,34 these data suggest that such a T cell response can probably be induced by damage of the cartilage, a site which is normally not exposed to blood perfusion and to the immune system.
Thus, these data do not tell us whether the observed CD8+ T cell response is just a bystander effect of cartilage destruction or whether the aggrecan-specific cellular immune response has a causative role. Because of the high HLA-B27 association with AS and because major histocompatibility complex class I molecules present antigens to CD8+ T cells we examined whether the same aggrecan derived CD8 epitopes could be identified in different patients and whether such peptides are presented by HLA-B27. If HLA-B27 restricted peptides are present this could be taken as an argument for their potentially pathogenetic role in AS.
Because the frequency of T cells in PB specific for autoantigens can be expected to be low we first enriched the antigen-specific CD8+ T cells by cytokine secretion technology,28 expanded these T cell lines in vitro, and used these lines for epitope mapping of the whole G1 domain of aggrecan. To our knowledge this is the first time that this technique has been successfully applied for CD8+ T cells specific for autoantigens. After separation of the antigen-specific CD8+ T cells according to their secretion of IFNγ by MACS we achieved an enrichment of about 60% of the G1 aggrecan-specific CD8+ T cells among the whole CD8+ T cell population. An expansion of these lines over 4 weeks was necessary to increase the amount of T cells used for epitope mapping. Interestingly, after 4 weeks the percentage of peptide-specific CD8+ T cells was still as high as about 10%. By using these lines we succeeded in identifying immunodominant peptides from the whole G1 aggrecan molecule. Interestingly, the significant antigen-specific CD8+ T cell response was observed even though we did not use nonameric peptides, but rather, overlapping 18 amino acid-long peptides. A similar good CD8+ T cell response to longer peptides has been described previously and might be explained by processing or trimming of longer peptides before or after binding to major histocompatibility complex class I molecules.35
However, the identified peptides differed among the patients. Furthermore, although some of the peptides (8, 9, and 17) contained an HLA-B27 binding motif (a nonamer peptide with arginine at position 2), these nonamers failed to induce a peptide-specific CD8+ T cell response. When T cell stimulation with these peptides was performed with an HLA-B27 blocking antibody in one patient no inhibition of T cell stimulation occurred. These results—different immunodominant peptides in different patients, the failure of the nonamer peptides to stimulate the G1-specific T cell lines, and the failure of anti-HLA-B27 antibodies to block this T cell response—clearly indicate that these peptides are not presented by HLA-B27.
It cannot yet be decided whether the failure to detect an HLA-B27 restricted T cell response is because we chose the wrong target protein (aggrecan) or whether this failure is an argument in favour of alternative hypotheses36–38 to explain the HLA-B27 association. For the CD8+ T cell response in patients with RA, further experiments have to be performed to investigate the role of these T cells. However, the fact that a CD4+ T cell response specific for aggrecan G1 can also be found in different rheumatic diseases24,32,33 makes a bystander effect also likely for the CD8+ T cell response in RA.
In the past we have applied different approaches for the identification of HLA-B27 restricted epitopes from Yersinia39 and Chlamydia40 in patients with reactive arthritis, a disease related to AS. The technique described here for the identification of antigen-specific CD8+ T cells seems to work even for low CD8+ T cell frequencies, which is important for T cells specific for autoantigens, and may help in the quest to find disease relevant “arthritogenic” peptides in the future. Using this sensitive technique, we have recently started to screen all cartilage derived proteins presented by HLA-B27 to CD8+ T cells.41 Future experiments will show whether T cell responses to cartilage antigens have a causative role in AS and related diseases.
Acknowledgments
This work was supported by grants from the Deutsche Forschungsgemeinschaft DFG: SFB 421, Projekt C1; Ap 82/2-1 and Ap82/2-2; Si 620/6-2.