Article Text

Abstract
OBJECTIVE Neutrophil elastase accounts for the ability of some fresh rheumatoid synovial fluids to degrade cartilage matrix in vitro. The aim of this study was to determine if enzyme activity could result from depletion of synovial fluid inhibitors or protection of the enzyme from inhibition.
METHODS The ability of synovial fluids to inhibit porcine pancreatic elastase was investigated together with chemical pretreatments capable of inactivating alpha1 protease inhibitor (α1PI) or preventing formation of alpha2 macroglobulin (α2M) elastase complexes. Subsequently, complexes of human neutrophil elastase with α2M were prepared and applied to frozen sections of cartilage. Proteoglycan loss was quantified by alcian blue staining and scanning and integrating microdensitometry. Parallel studies were carried out using a low molecular weight chromogenic elastase substrate. The effects of α1PI and SF on these systems were investigated. Finally, synovial fluids were subjected to gel filtration and the fractions assayed for elastase activity. High molecular weight fractions were pooled, concentrated, and tested for their ability to degrade cartilage sections.
RESULTS All synovial fluids reduced the activity of porcine pancreatic elastase, the inhibition mainly being attributable to α1PI, whereas remaining activity resulted from complexes of elastase with α2M. Complexes of human neutrophil elastase with α2M were shown to cause proteoglycan degradation in frozen sections of human articular cartilage. Alpha1PI prevented α2M elastase complexes from degrading cartilage but not the chromogenic substrate. The data suggested that α1PI does not inhibit elastase bound to α2M but sterically hinders the complex. However, only one of five synovial fluids was able to completely block the actions of α2M elastase complexes against cartilage. Gel filtration of rheumatoid synovial fluids showed elastase and cartilage degrading activity to be associated with fractions that contained α2M, and not with fractions expected to contain free enzyme.
CONCLUSIONS The data suggest that synovial fluid α2M elastase complexes can degrade cartilage matrix in rheumatoid arthritis.
- alpha1 antitrypsin
- alpha2 macroglobulin
- glycosaminoglycans
- neutrophil elastase
- proteoglycans
- synovial fluid
Statistics from Altmetric.com
- alpha1 antitrypsin
- alpha2 macroglobulin
- glycosaminoglycans
- neutrophil elastase
- proteoglycans
- synovial fluid
The cellular pathology of rheumatoid arthritis (RA) is interesting in that the synovial tissue is infiltrated by chronic inflammatory cells whereas the synovial fluid often contains an abundance of neutrophils,1 which typically characterise acute inflammation. The role of the neutrophil in contributing to joint destruction in RA is unknown. In vitro, neutrophil elastase is capable of degrading cartilage proteoglycan2 ,3 and is the main mechanism by which activated neutrophils degrade dead cartilage matrix4 and may have a role in the initial disruption of the superficial layer of articular cartilage in RA.5Reactive oxygen species from activated neutrophils seem to modulate elastase activity through oxidative inactivation of an endogenous elastase inhibitor.6
Up to 50% of fresh cell free RA synovial fluids cause glycosaminoglycan (GAG) loss from sections of human articular cartilage.7 Partial protection against cartilage GAG loss was consistently seen in the presence of a low molecular weight neutrophil elastase inhibitor (ONO-5046) but not with inhibitors of other classes of endopeptidases. This seems to be a direct proteolytic effect of synovial fluid because dead cartilage matrix was used. Cartilage degrading activity of synovial fluids was largely lost on frozen storage.
The major elastase binding proteins in synovial fluid are alpha1 protease inhibitor (α1PI) (also termed alpha1 antitrypsin) and alpha2 macroglobulin (α2M). Alpha1PI in particular has been shown to be sensitive to inactivation by reactive oxygen species and proteases with inactive molecules being recovered from RA synovial fluid.8 Therefore, one explanation for the ability of RA synovial fluids to proteolytically degrade cartilage matrix may simply be attributable to the depletion of elastase inhibitors. An alternative explanation is that elastase forms a complex with an elastase binding protein and that this complex can remain active in the presence of functional elastase inhibitors. This study is aimed at determining which of these explanations is valid.
Methods
CHEMICALS
Alpha2M, α1PI and human neutrophil elastase were from Calbiochem (Novabiochem (UK) Ltd, Nottingham, UK). All remaining chemicals were from Sigma Chemical Company (Poole, Dorset, UK).
ELASTASE ASSAY9
0.2 M TRIS/HCl buffer at pH 8.0 was supplemented with 0.01M CaCl2 and 0.01% Brij 35. Methoxy-succinyl-Ala-Ala-Pro-Val-p-nitroanilide was prepared at 100 mM in N-methyl-2-pyrrolidone and diluted with buffer such that its final concentration in the assay was 1 mM. Assays were carried out in 96 well flat bottomed microtitre plates. Samples and substrate were incubated at 25°C for 30 minutes and then the reaction stopped by the addition of 50% aqueous acetic acid. Releasedp-nitroanilide was quantified at 405 nm using a Biotek EL 310 plate reader. Results were expressed in units of activity, one unit defined as the release of 1 mmolp-nitroanilide from the substrate per minute at 37°C.
SYNOVIAL FLUIDS
Synovial fluids were obtained from the knee joints of RA patients fulfilling the American Rheumatism Association’s 1987 diagnostic criteria9 with clinical indication for joint aspiration and were collected into 10 mM EDTA to prevent clotting and the possible confounding effects of metalloproteases. Cells were removed by centrifugation at 2000 g for 10 minutes and the supernatants were used fresh or stored at −70°C as stated in individual experiments.
ELASTASE BINDING CAPACITY OF SYNOVIAL FLUIDS
Synovial fluids that had been stored at −70°C were used. These were untreated or treated for four hours at room temperature with methylamine (200 mM) or chloramine T (200 mM). Unreacted chloramine T in the sample was reacted with 400 mM methionine to prevent interference with the subsequent elastase assay. A sequential treatment with chloramine T and methionine was also used. After treatments, synovial fluid samples were doubly diluted in a 96 well flat bottomed microtitre plate. Porcine pancreatic elastase was added to give a final concentration per well of 1000 U/ml. Control wells received an equal volume of buffer. Elastase activity was measured as above.
MICROASSAY OF CARTILAGE PROTEOGLYCAN LOSS
Human cartilage samples were obtained after death from adults within six hours of death and were removed from macroscopically normal areas of the non-weight bearing posterior surface of the femoral condyle. Blocks of 3 mm3 were cut, snap frozen by precipitate immersion in n-hexane at −80°C and stored in liquid nitrogen before use. Cryostat sections were cut at 4 μm and mounted onto 12 spot Teflon coated microscope slides that had previously been treated with 10% poly-L-lysine as a section adhesive. Sections were then subjected to incubation as described below. After the incubation, sections were fixed with 10% formal saline and sulphated GAGs were stained with alcian blue (8GX, 0.05% in 0.025 M acetate buffer containing 0.5 M MgCl2) over night. Dye content was quantified with a Vickers M85 scanning and integrating micro densitometer using a × 40 objective, a wave length of 550 nm, a band width of 20 nm, a mask diameter of 15 μm, and scanning spot size of 0.2 μm. For each section, clear field readings (background) and clear field readings with an imposed neutral density of 1.0 (ND1) were taken. Six integrated readings were taken from areas of cartilage matrix free of lacunae. From these values, the mean integrated extinction (MIE) was calculated as (mean integrated absorbance−background)/ND1. Results were expressed as GAG loss relative to free elastase or mean (SEM) percentage GAG loss.
PREPARATION OF ELASTASE α2M COMPLEXES
Human neutrophil elastase (68 nM) and 106 nM α2M (a molar ratio of 1:1.5) were incubated in 0.1 M TRIS/HCl buffer (pH 7.6) supplemented with 0.01 M CaCl2 at 37○C for 30 minutes. The absence of free elastase was confirmed by size exclusion chromatography (below).
SIZE EXCLUSION CHROMATOGRAPHY OF SYNOVIAL FLUIDS
A 40 × 1.8 cm column was packed with Sephadex G75 (Pharmacia, Milton Keynes, UK) and equilibrated with 50 mM TRIS/HCl buffer pH 7.6 supplemented with 0.14 M NaCl and 0.02% sodium azide. The linear flow rate was 12 cm/h and 1.8 ml fractions were collected. A mixture of carbonic anhydrase (29 kDa) and thyroglobulin (800 kDa) was used to calibrate the column. Fresh synovial fluid supernatants were treated with hyaluronidase (30 U/ml for 30 minutes at 37°C) to reduce viscosity. The hyaluronidase was shown to be free of elastolytic activity. Samples of 1 ml synovial fluid were applied to the column. The fractions were tested for elastase activity by measuring the release of p-nitroaniline from methoxy-succinyl-Ala-Ala-Pro-Val-p-nitroanilide over 24 hours. Fractions to be concentrated were pooled as appropriate and their volumes reduced to 1 ml by positive pressure dialysis against 40% polyethylene glycol.
Results
ELASTASE BINDING CAPACITY OF RA SYNOVIAL FLUIDS
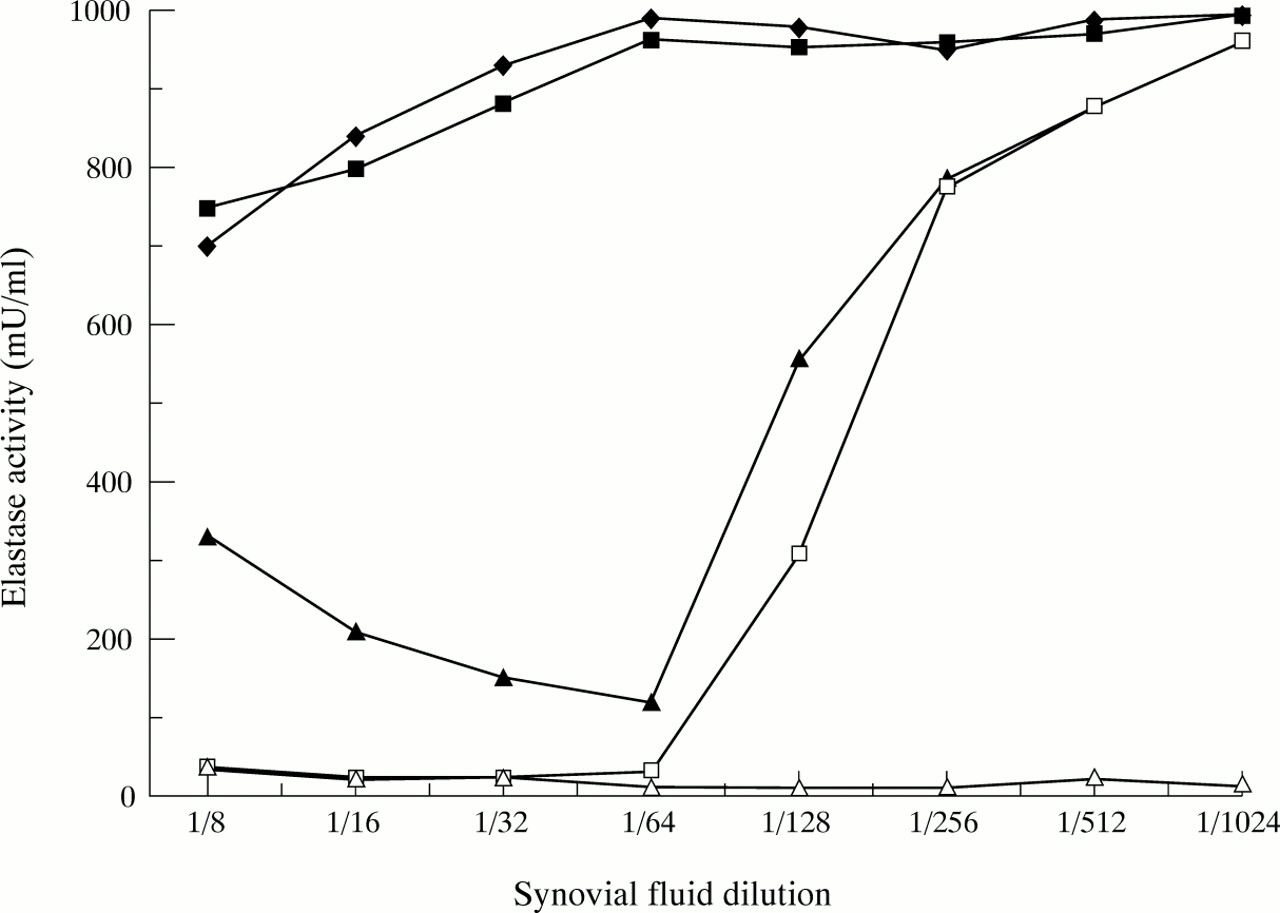
Twelve RA synovial fluid supernatants were examined with similar results. Figure 1 shows a typical experiment. RA synovial fluid supernatants that had been stored at −70°C were shown to have no appreciable elastase activity over a 30 minute incubation period with substrate. All synovial fluids, irrespective of whether they were initially able to degrade cartilage sections, were capable of reducing but not totally inhibiting the activity of added elastase. However, as the synovial fluid was diluted, its ability to reduce the activity of added elastase was first increased and then rapidly diminished. Pretreatment of synovial fluid with chloramine T to oxidatively inactivate α1PI11 largely removed the elastase inhibitory capacity of the synovial fluid. When synovial fluids were pretreated with methylamine, a process that renders α2M incapable of binding proteases,12 the ability of synovial fluids to inhibit elastase was increased. Table 1shows data for the lowest dilution of synovial fluids (1/8) in more detail. There were statistically significant differences in the ability of non-treated synovial fluid aliquots and those pretreated with methylamine or chloramine T to inhibit the activity of added elastase.

The elastase binding capacity of RA synovial fluid. Incubations were for 30 minutes at 37°C in the presence of methoxy-succinyl-ala-ala-pro-val-pNA (a chromogenic substrate). Open triangles = elastase activity of untreated synovial fluid. All other determinations were carried out with the addition of 1000 mU/ml porcine pancreatic elastase to each synovial fluid dilution. Solid triangles = elastase + synovial fluid. Open squares = elastase + synovial fluid pretreated with methylamine. Solid diamonds = elastase + synovial fluid pretreated with chloramine T. Solid squares = elastase + synovial fluid pretreated with both methylamine and chloramine T.
Elastase activity of RA synovial fluids at a 1/8 dilution after addition of 1000 mU/ml porcine pancreatic elastase showing the effects of methylamine or chloramine T pretreatment
Human neutrophil elastase was shown to degrade proteoglycan from frozen sections of cartilage matrix. In preliminary experiments, α2M was added to elastase up to a mol ratio of 1.5:1 without significant effect on the ability of elastase to degrade cartilage. In subsequent experiments complexes were always formed in the ratio of 1.5:1. Free elastase and complexed elastase were subjected to gel filtration and the fractions assayed for elastase activity. The free enzyme eluted in fractions consistent with its molecular weight of 29 kDa whereas the complexed enzyme eluted with the void volume. There was no elastase activity in the fractions that would have been expected to contain any free enzyme (data not shown).
Addition of α1PI (2 μM) to free human neutrophil elastase (68 nM) caused total inhibition of enzyme activity against the chromogenic substrate and cartilage matrix (fig 2). If the elastase was first complexed to α2M, before the addition of α1PI, the activity of the enzyme was greatly diminished against the chromogenic substrate and no significant activity was seen against cartilage matrix. The complexes retained 13% of the activity when α1PI were not present. A similar result was achieved when these combinations were applied to cartilage sections.

The effects of α1PI (2 μM) on the ability of free human neutrophil elastase (NE) (68 nM) or elastase in the presence of α2M (106 nM) to cleave either a low molecular weight chromogenic substrate or degrade cartilage matrix. To compare the two assays, data have been expressed as activity ratios (activity of elastase α2M complexes/activity of free enzyme). Mean (SEM), n=5.
Synovial fluid supernatants that were shown to lack elastase activity over a 30 minute incubation with MeO-suc-ala-ala-pro-val-pNa were tested for their effects on the activity of elastase α2M to degrade cartilage matrix. Figure 3 shows that of five synovial fluids tested all reduced the activity of the complexes against cartilage. However, only in the case of one sample was activity against cartilage matrix completely inhibited.

Proteoglycan loss loss from 4 μm cryostat sections of cartilage treated for 24 hours with 68 nM human neutrophil elastase (NE) in the presence of 106 nM α2M. Also shown are the effects of five RA synovial fluids supernatants at 40% (v/v) on this degradation. Means (SEM), n=5.
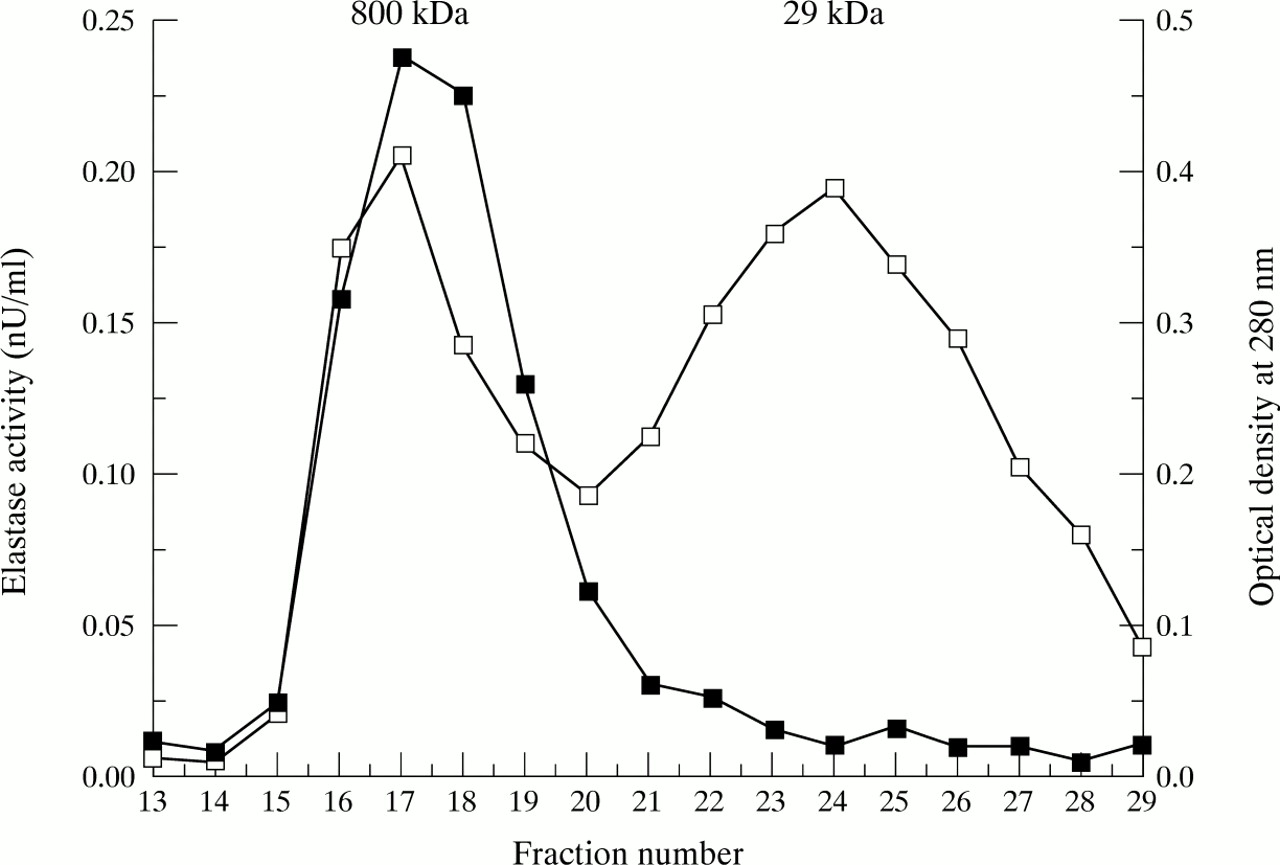
Figure 4 shows gel filtration of a fresh RA synovial fluid that was capable of degrading cartilage matrix by an elastase dependent mechanism. Elastase activity was associated with those fractions that would contain species of 800 kDa (the void volume) and not those fractions that would be expected to contain free enzyme (29 kDa). When the high molecular weight fractions (fractions 15–20) were pooled and concentrated to the same volume applied to the column, they were able to account for the degradative activity of the whole synovial fluid (percentage (SEM) GAG loss pooled fractions 42 (5); synovial fluid 36 (4)).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Gel filtration of RA synovial fluid (solid squares) showing absorbance of fractions at 405 nm after 24 hour incubation with methoxy-succinyl-ala-ala-pro-val-pNA. Superimposed (open squares) is the gel filtration of a mixture of thyroglobulin (800 kDa) and carbonic anhydrase (29 kDa) for calibration purposes.
Discussion
In vitro, neutrophil elastase has been shown to degrade cartilage proteoglycan and collagen. Nevertheless, in RA, degradation of cartilage matrix by synovial fluid neutrophil elastase has been considered unlikely because of a putative excess of protease inhibitors, principally α1PI and α2M.13 However, the oxidative and proteolytic attack of these proteins may render them ineffective in binding neutrophil elastase thereby allowing enzyme activity to be expressed.
In this study we have developed a simple assay for the elastase binding capacity of RA synovial fluid. The assay is functional, as compared with immunoreactive, and therefore indicates the level of active elastase binding proteins. All synovial fluids tested, irrespective of whether they were capable of degrading sections of cartilage, were shown to be capable of reducing the activity of added elastase. Paradoxically, the ability to reduce elastase activity initially increased with dilution before being lost at high dilution (fig 1). The simplest explanation for this result is to suppose two elastase binding proteins one of which is in excess and inactivates the enzyme and the other that allows the enzyme to remain active. Diluting the fluid would allow less enzyme to bind to the second protein and therefore more enzyme to be inactivated. Further dilution would result in increased enzyme activity as a proportion of the enzyme became free.
The major elastase inhibitor of synovial fluid was shown to be α1PI because oxidative inactivation with chloramine T12 greatly reduced the elastase inhibitory capacity. Those synovial fluids that were degrading cartilage through an elastase dependent mechanism were therefore doing so in the presence of an excess of active α1PI. Elastase added to high concentrations of synovial fluid was totally inhibited when the synovial fluid was pretreated with methylamine, a process that renders α2M incapable of binding proteases.13 This indicated that the untreated synovial fluid contained active α2M and that the elastase α2M complex remained active against the chromogenic substrate used in the assay.
Although considered a protease inhibitor, proteases bound to α2M retain activity against low molecular weight substrates.14 By contrast, macromolecular substrates are generally protected from degradation. However, activity against a specific polypeptide15 and protein substrates14 ,16 ,17 has been observed. The reason why some α2M protease complexes can degrade macromolecular substrates is unclear but may be because of inadequate trapping of large proteases (for example, plasmin) or a greater freedom of movement of proteases that are not covalently bound to α2M. Human neutrophil elastase does not bind covalently to α2M,18 in addition, the resulting complexes are unstable because the trapped elastase is able to cause autolysis of the complex.19 If this autolysis were to occur in the presence of α1PI then the released enzyme would be inactivated. Elastase α2M complexes provide a rational explanation for the ability of RA synovial fluids to degrade cartilage matrix and for the loss of such activity on storage. We therefore tested the ability of elastase α2M complexes to degrade cartilage matrix or to cleave a low molecular weight chromogenic substrate.
Elastase α2M complexes were shown to be capable of degrading cartilage proteoglycan (fig 2). This degradation could not be attributed to contaminants in the commercial preparations used because α2M alone had no degradative effect on cartilage and only a selective elastase inhibitor (ONO-5046) was able to block degradation by elastase or elastase α2M complexes (data not shown). The complexes used were in a mol ratio of α2M to elastase of 1.5:1. The ultrastructure of α2M shows the molecule to be a twin trap20 capable of binding two elastase molecules so a 1.5 molar excess is theoretically more than is necessary to bind all the free elastase. This was confirmed with gel filtration experiments that indicated that there was no detectable free elastase in the complex preparations.
Complexed enzyme was always less active against cartilage proteoglycan than against the low molecular weight substrate. This may be attributable to a proportion of the complexes containing two elastase molecules. Because of the polar orientation of such complexed enzymes, only one would be expected to engage cartilage at any one time. Therefore activity against cartilage will be less than activity against a low molecular weight soluble substrate.
If elastase α2M complexes in synovial fluid can degrade cartilage matrix they must be functional in the presence of α1PI. Our data already indicated that complexes were active in synovial fluid against a low molecular weight substrate (fig1), and other workers have shown protease α2M complexes to express proteolytic activity in the presence of high molecular weight inhibitors.21 We were therefore surprised to find that α1PI reduced the ability of elastase α 2 M complexes to cleave the low molecular weight chromogenic substrate and prevented complexes from degrading cartilage (fig 2). However, reduction in enzyme activity towards the chromogenic substrate could not be attributed to inhibition of the enzyme since the α1PI concentration was more than enough to completely inhibit the free enzyme. A more likely explanation is that α1PI became bound to the complexes thereby sterically hindering access to the chromogenic substrate and completely blocking access to cartilage matrix. This is entirely plausible given the known ability of α2M to act as a carrier of polypeptides after protease activation.22 In a synovial fluid environment these binding sites could be occupied by a range of molecules with variable effects on the ability of the complexes to degrade high and low molecular weight substrates. When synovial fluids were “spiked” with elastase α2M complexes their activity against cartilage matrix was reduced but not always inhibited (fig 3).
Finally, using gel filtration of a synovial fluid capable of degrading cartilage matrix, we were able to show that elastase activity was only associated with the void volume fractions that contained α2M and not with fractions that would be expected to contain free enzyme (fig 4). When the high molecular weight fractions were pooled and concentrated to the original volume applied to the column, they were able to fully account for the cartilage degrading activity of the whole synovial fluid. The data therefore support a role for elastase α2M complexes in synovial fluid mediated cartilage destruction.
Estimated half times of association in vivo for elastase/α1PI (0.61 ms) and elastase/α2M (7.2 ms) suggest α1PI to have the higher affinity for elastase.23 However, increased binding of elastase to α2M may occur in the pericellular environments of activated neutrophils where release of reactive oxygen species could cause a local inactivation of α1PI. Certainly, complexes of elastase to α2M can be demonstrated in synovial fluids of patients with RA where their concentrations correlate with the synovial fluid leucocyte counts.24 Injection of α2M protease complexes into the knee joints of rabbits causes the development of a synovitis25 and various immunomodulatory activities have been ascribed to α2M protease complexes that may have a role in the pathogenesis of RA.26 ,27 However, direct degradation of joint structures by α2M protease complexes has not previously been described. These results may have implications for other disorders where neutrophil elastase has been implicated, for example adult respiratory distress syndrome and cystic fibrosis.
Acknowledgments
This work was supported by the Arthritis and Rheumatism Council, UK, The Hyal Research Foundation and Ono Pharmaceutical Company, Japan.