Article Text

Abstract
OBJECTIVE To analyse the functional activity of the plasminogen activators urokinase (uPA) and tissue type plasminogen activator (tPA) in human synovial membrane, and to compare the pattern of expression between normal, osteoarthritic, and rheumatoid synovium. The molecular mechanisms underlying differences in PA activities between normal and pathological synovial tissues have been further examined.
METHODS Synovial membranes from seven normal (N) subjects, 14 osteoarthritis (OA), and 10 rheumatoid arthritis (RA) patients were analysed for plasminogen activator activity by conventional zymography and in situ zymography on tissue sections. The tissue distribution of uPA, tPA, uPA receptor (uPAR), and plasminogen activator inhibitor type-1 (PAI-1) was studied by immunohistochemistry. uPA, tPA, uPAR, and PAI-1 mRNA values and mRNA distribution were assessed by northern blot and in situ hybridisations respectively.
RESULTS All normal and most OA synovial tissues expressed predominantly tPA catalysed proteolytic activity mainly associated to the synovial vasculature. In some OA, tPA activity was expressed together with variable amounts of uPA mediated activity. By contrast, most RA synovial tissues exhibited considerably increased uPA activity over the proliferative lining areas, while tPA activity was reduced when compared with N and OA synovial tissues. This increase in uPA activity was associated with increased levels of uPA antigen and its corresponding mRNA, which were localised over the synovial proliferative lining areas. In addition, in RA tissues, expression of the specific uPA receptor (uPAR) and of the plasminogen activator inhibitor-type 1 (PAI-1) were also increased.
CONCLUSION Taken together, these results show an alteration of the PA/plasmin system in RA synovial tissues, resulting in increased uPA catalytic activity that may play a part in tissue destruction in RA.
- plasminogen activation
- urokinase activity
- rheumatoid arthritis
- osteoarthritis
Statistics from Altmetric.com
Joint damage with cartilage and bone destruction is a common feature of arthritic disorders.1-4 Increased extracellular proteolysis involving serine and metalloproteases is thought to play a critical part in these destructive processes.5 The serine proteases plasminogen activators (PAs) have been studied intensively in the degradation of extracellular matrices in the context of physiological and pathological tissue remodeling and cell migration events such as ovulation, mammary gland involution or tumour cell invasion.6 Two types of PAs have been identified in mammals, urokinase (uPA) and tissue type PA (tPA), which catalyse the conversion of the zymogen plasminogen into plasmin. Plasmin, a protease of broad specificity, has the capacity to degrade most of the extracellular proteins including proteoglycans, directly or indirectly through the activation of procollagenase into collagenase.7 The activity of PAs is controlled by natural inhibitors (PAIs), which include PAI- l, the main PAI in the circulation.8 Another important step in the regulation of PA activity involves binding of tPA to fibrin, leading to enhanced catalytic activity of the enzyme, and binding of uPA to a specific, high affinity cell surface receptor, uPAR9; binding of uPA to uPAR increases the rate of plasminogen activation10 and greatly enhances extracellular matrix degradation and ceIl invasion in vitro and in vivo.11 ,12
Clinical and experimental evidence suggest a contribution of the PA system to joint pathogenesis in osteoarthritis (OA) and rheumatoid arthritis (RA). In synovial fluid from RA patients, antigenic values of uPA and PAI- l were increased compared with plasma values13–15 or with normal knee synovial fluid16 while those of tPA were decreased13–15or unchanged.16 In one study, the enhanced uPA and decreased tPA antigenic activities measured in synovial fluid have been associated with the clinical severity of RA.17 Clinical improvement of patients with OA or RA was observed after intra-articular injection of urinary trypsin inhibitor, which was shown to inhibit uPA.18 ,19 Immunohistological studies on synovia of OA and RA patients showed an increase of uPA, uPAR, and PAI in RA tissues whereas tPA values were comparable between the different patients.20 ,21 Several cells present in the joint (for example, mononuclear and polymorphonuclear cells, synovial fibroblasts, endothelial cells, and chondrocytes) have been shown to express PAs and their inhibitors whose production can be modulated by locally generated cytokines.22 ,23 Altogether, these results suggest that in arthritides local synovial production of some components of the PA system, which accumulate subsequently in synovial fluid, could promote degradation of joint cartilage and bone through PA mediated proteolytic activity.
To date, however, the published results do not provide a functional picture of PA mediated proteolytic activity in OA and RA tissues, as they relied exclusively on antigenic assays. Measurement of net PA proteolytic activity is particularly crucial in these tissues because of the concomitant expression of PA enzymes and their inhibitors.21 Moreover, most of the studies did not evaluate the PA system in normal joints, thus precluding any conclusion on its contribution in the pathophysiology of the joints. This prompted us to analyse directly PA enzymatic activities as well as expression of individual components of the PA system by the combined use of histological zymography, immunohistology, and mRNA hybridisation in normal synovial tissues and in synovial tissues of patients suffering from advanced OA or RA.
Methods
TISSUE SAMPLING AND HISTOLOGICAL EXAMINATION
Specimens of synovial tissue from 14 OA and 10 RA patients undergoing joint surgery of the knee or the hip for advanced disease, were obtained from the Department of Orthopaedics (CHUV, Lausanne). OA patients were diagnosed by clinical and radiological criteria24 and RA patients fulfilled at least four of the seven American Rheumatism Association revised criteria for RA.25 Seven normal synovial tissues were obtained at necropsy (Institute of Pathology, Lausanne) from people with no history of joint disease. All tissues were cut in small pieces and immediately frozen in precooled hexane and stored at −70°C until use. Sections (10 μm) of synovial tissue samples were analysed after staining with haematoxylin and eosin. Only tissue samples that histologically showed a synovial lining cell layer were used in this study.
ENZYMATIC ASSAYS
Histological zymographies were performed on 10 μm cryostat sections as described elsewhere.26 ,27 Briefly, sections were overlaid with 130 μl of a mixture containing 2% non-fat dry milk, 0.9% agar, and 40 μg/ml of purified human plasminogen in PBS (with 0.9 mM Ca2+ and 1 mM Mg2+). Control experiments were carried out with overlay mixtures from which plasminogen was omitted. To distinguish uPA from tPA, amiloride, a specific inhibitor of uPA enzymatic activity,28 was added in the overlay mixture at 1 mM final concentration; the slides were then incubated in a humid chamber at 37°C for one to three hours and photographs taken using dark ground illumination.
Tissue protein extracts were analysed by SDS-PAGE zymographies as described before. 26 Briefly, after SDS-PAGE of the samples, the gel was washed in Triton X-100 and layered over a casein underlay containing the same mixture as described above for in situ zymographies.
TISSUE PROTEIN EXTRACTS PREPARATION
Ten to twenty 30 μm cryostat sections of synovial tissue were scraped with a scalpel and homogenised in 50 mM TRIS-HC1 pH 7.5, containing 110 mM NaCl, 10 mM EDTA, 5 mM iodoacetamide, and 0.1% NP-40. The homogenate was centrifuged at 4000 g for 10 minutes at 4°C and the supernatant stored at −20°C. Protein content of the tissular extracts was measured by the method of Bradford using BSA as a standard.
ANTIBODIES
Murine monoclonal antibodies against uPA (ref 394), uPAR (ref 3936), goat polyclonal antibodies against tPA (ref 387) and PAI-1 (ref 395G) were purchased from American Diagnostica Inc (Greenwich, CT, USA). Monoclonal antibodies against CD31 (ref M0823) were obtained from Dako A/S (Denmark).
IMMUNOHISTOCHEMISTRY
Immunohistochemistry was performed on air dried 5 μm cryostat tissue sections stored at −20°C and then fixed for 10 minutes in acetone at 4°C before use. Each slide was incubated for 30 minutes with 10% normal human serum and 10% normal serum of the species in which the secondary biotinylated antibody has been raised. Slides were then overlayed with the primary antibody overnight at 4°C. Bound antibodies were visualised by the avidin-biotin-peroxydase complex (Vectastain Elite ABC kit, Vector Laboratories, Burlingame, CA, USA). All incubations were carried out in a humidified chamber. The colour was developed by 3-3’-diaminobenzidine (DAB, Sigma) containing 0.01% hydrogen peroxide. After being extensively washed in water, each slide was counterstained with Papanicolaou’s Harris solution (Merck) and further dehydrated in graded alcohol and mounted. To determine the specificity of the staining, controls were done using, as primary antibodies, isotype matched mouse immunoglobulins of irrelevant specificity, or purified preimmnune goat IgG. An incubation in which the first antibody was omitted served as a negative control.
PLASMID CONSTRUCTIONS AND IN VITRO TRANSCRIPTION
Plasmid constructions for uPA antisense and sense probes (a 610-bp EcoRI/PstI fragment), as well as uPAR (a 584-bp BamHI/BamHI fragment), tPA (a 614-bp BglII/EcoRI fragment), and PAI-1 (a 1.4 Kbp EcoRI/BgLII) antisense probes were prepared exactly as described previously.27 Linearised plasmids were used as templates for bacteriophage RNA polymerase in the presence of32P-labelled UTP (400 Ci/mmol) as described elsewhere.29 The specific activity of the RNA probes is approximately 3 × 108 cpm/μg.
RNA ANALYSES
In situ hybridisations were carried out on 5 μm cryostat tissue sections, which were hybridised to 32P-labelled probes and revealed by film autoradiography as described previously.23Total RNA extractions from frozen synovial tissue specimens were performed using the Trizol reagent (GibcoBRL), according to the manufacturer’s instructions and the amount of RNA was quantified spectrophotometrically. Twelve μg/lane of total RNA were analysed by northern blot as described elsewhere.29 Filters were exposed to Kodak XAR-5 films at −80°C between intensifying screens. Relative mRNA values were determined by laser densitometric analysis, normalising the values to an internal standard (HT1080 mRNA) loaded on each northern blot.
STATISTICAL CALCULATIONS
The Mann-Whitney test for unpaired variable (two tailed) was used to compare differences between groups. p Values < 0.05 were considered significant.
Results
PA ACTIVITIES EXPRESSED BY NORMAL, OA, AND RA SYNOVIAL TISSUES
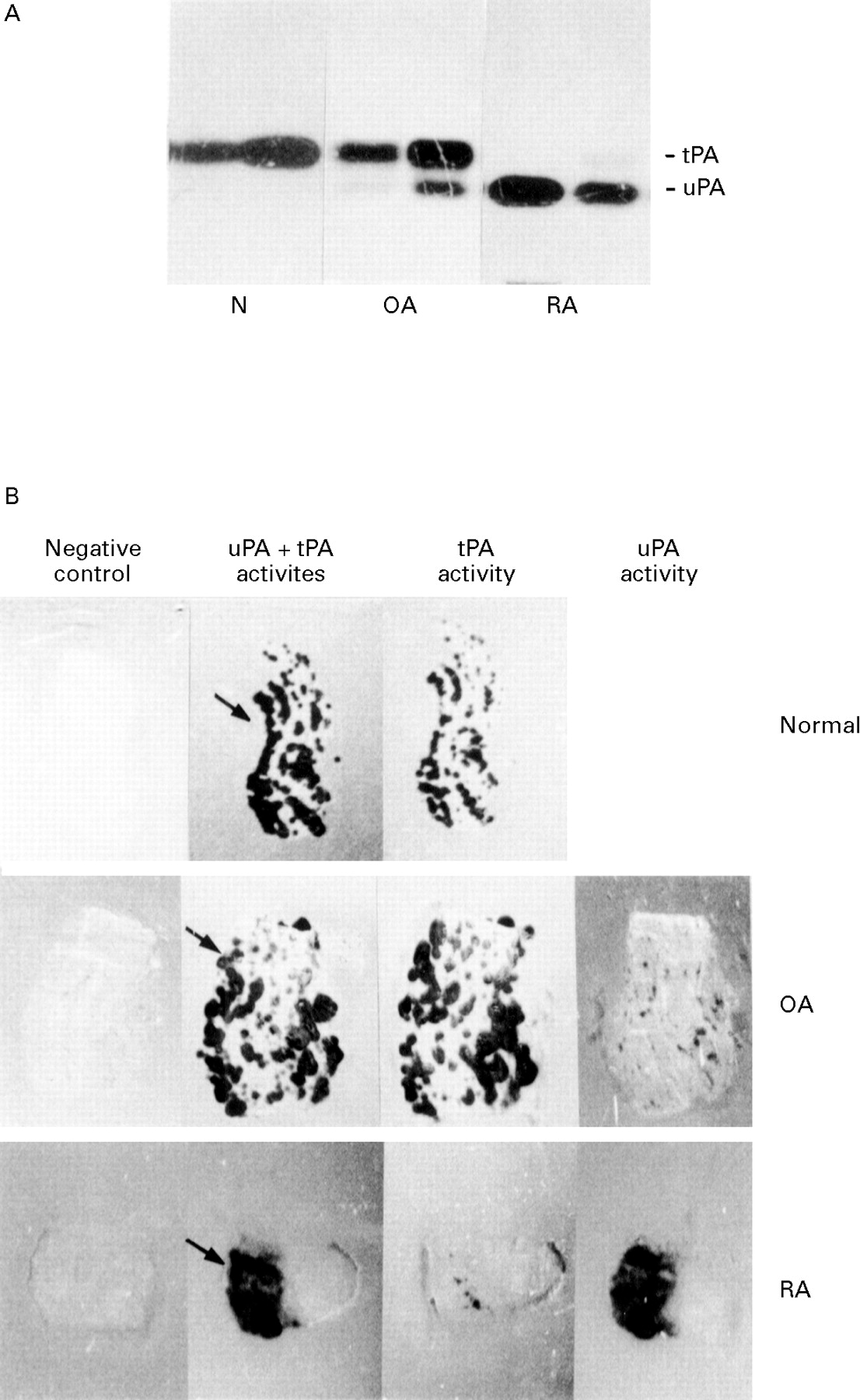
Tissue extracts were prepared from seven N, 14 OA, and 10 RA synovial tissues, and analysed by zymography (fig 1A). This permitted clear distinction between uPA and tPA mediated activities, which migrated at 55 and 70 kDa respectively. Normal synovial tissues expressed exclusively tPA activity. OA synovial tissues expressed predominantly tPA activity together with variable amounts of uPA activity. By contrast, uPA activity was detectable in all RA synovial tissues and represented the major PA activity in six of 10 tissues. In addition in these RA tissues, when detectable, tPA activity levels were always reduced when compared with those measured in N and OA synovial tissues. In situ zymograms (fig 1B) performed on synovial tissue sections revealed in all specimens analysed (31 of 31) areas of plasminogen dependent caseinolysis after one to three hours of incubation at 37°C. No lysis was observed in the absence of plasminogen even after a longer period of incubation. In N and OA tissues, these areas of plasminogen dependent caseinolysis were scattered throughout the entire section, usually in a punctate pattern (second column). Addition of amiloride, an inhibitor of uPA emzymatic activity in the overlay mixture, did not affect these areas of caseinolysis (third column), whereas addition of anti-tPA antibodies abolished the caseinolytic activity (not shown for N, fourth column for OA), indicating that the observed caseinolysis was caused by tPA. From a microscopic examination of serial frozen sections of N and OA synovial tissues stained by immunohistochemistry using an antibody against endothelial cells (CD31), we could deduce that the observed tPA activity was detected mainly in association with endothelial cells (not shown). In RA tissue specimens, areas of caseinolysis were present over the synovial proliferative lining area (second column). This caseinolytic activity was almost totally inhibited by the addition of amiloride (third column) but was not modified by anti-tPA antibodies (fourth column), indicating that the catalytic activity associated with RA tissues was mediated mainly by uPA. In some RA tissues additional areas of caseinolysis related to tPA (that is, not modified by amiloride) were observed in the deep layer of the synovial membrane (see fig 4B) The results obtained by zymographies of tissue protein extracts corroborated the results obtained by in situ zymographies on tissue sections and are summarised in table1.

PA activities expressed by N, OA, and RA synovial tissues. (A) Zymographic analysis. Equal amounts of proteins (5 μg), from N synovial tissue extracts or from synovial tissue extracts of OA and RA patients were analysed by SDS-PAGE zymographies as outlined in the Methods section. Two representative samples from two different subjects are shown for each group. (B) Histological zymograms on sections of normal synovial tissue and of synovial tissue from OA and RA patients. No lysis was observed in the absence of plasminogen (negative control, first column). Caseinolytic areas appear, in the presence of plasminogen, under dark ground illumination, black on the figure (uPA +tPA activities, second column). Lysis zones on N and OA synovial tissue correspond to tPA, as they were not inhibited by amiloride (tPA activity, third column), but by antibodies against tPA (uPA activity, fourth column). RA tissues displayed caseinolytic areas that were almost totally inhibited by amiloride, but only slightly inhibited by anti-tPA antibodies. Arrows denote the localisation of the synovial lining layer, facing the synovial fluid. Photographs of zymograms were taken after two to three hours’ incubation at 37°C. Original magnification × 3.

Visualisation of PA activities and mRNAs on tissue sections of synovial membranes from OA and RA patients. (A) OA synovial tissue. Top row: in situ zymographies. OA histological zymograms show amiloride resistant PA enzymatic activity corresponding to tPA. Bottom row: in situ hybridisations. Hybridisation of consecutive sections show an accumulation of uPA mRNA mainly in the synovial lining layer whereas no uPAR mRNA is detected. (B) RA synovial tissue. Top row: in situ zymographies. RA histological zymograms show both amiloride resistant PA activity resulting from tPA, in the deep lining layer, and amiloride sensitive PA activity corresponding to uPA, associated to the lining border and the synovial proliferative lining area. Bottom row: in situ hybridisations. Hybridisation of consecutive sections show an accumulation of uPA and uPAR mRNAs mainly in the synovial lining layer. Control hybridisation was performed with the uPA sense probe. Autoradiograms of in situ hybridisations were exposed for two weeks at room temperature. Arrows denote the localisation of the synovial lining layer, facing the synovial fluid. Original magnification × 2.
PA activities in normal synovial tissues and synovial tissues of OA and RA patients
ANTIGENIC LOCALISATION OF uPA,uPAR, tPA, AND PAI-1
Immunohistochemical analysis of uPA,uPAR, tPA, and PAI-1 was performed on three N, four OA, and four RA synovial tissues. Negative controls (omission of the first specific antibody or replacement of the first specific antibody by an irrelevant antibody) gave no signal at all. In normal synovial tissues, no positivity or only weak signals were observed using anti-uPA antibodies. By contrast, uPA immunoreactivity in OA was moderate to abundant, mainly localised to the synovial lining cell area. In two of four OA synovial tissues, some interstitial fibroblasts and endothelial cells were also uPA positive. An even more abundant uPA immunoreactivity was found in all the synovial RA biopsy specimens examined localised to the lining cells and interstitial cells (fibroblasts and mononuclear cells) and associated with most of the blood vessels.
uPAR was not detectable in any of the normal synovial tissues and was weakly present in OA synovial tissues, but when detected (in two of four OA tissues), its expression was restricted to the synovial lining cells. By contrast, expression of uPAR was found in all RA synovial tissues, mainly associated to the lining cells, but also to some mononuclear cells in the deep layers. Figure 2 shows representative examples of immunohistochemical staining of uPA and uPAR.
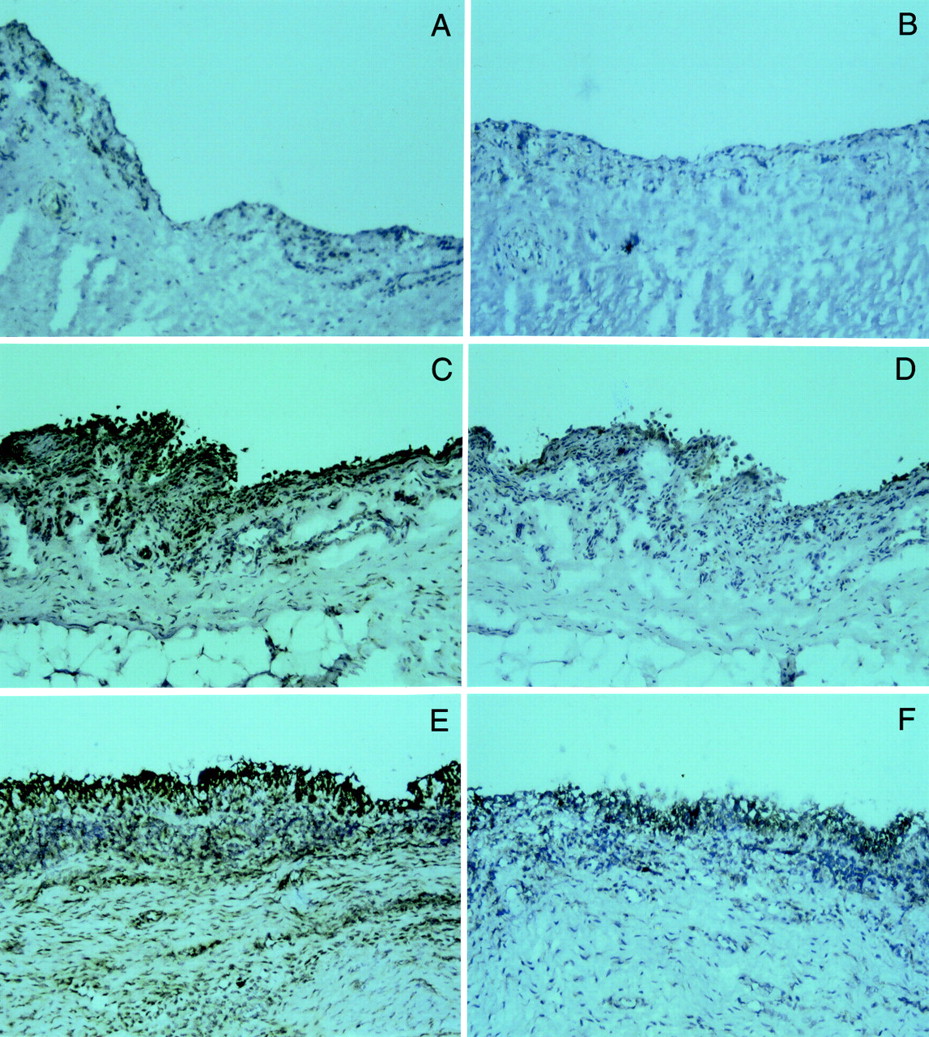
Immunohistochemical localisation of uPA and uPAR antigens in N, OA, and RA synovial tissues. Cryostat sections of N (A, B), OA (C, D), and RA (E, F) synovial tissues were incubated with monoclonal antibodies against uPA (left panels, A, C, E) or uPAR (right panels, B, D, F). Bound antibodies were visualised by the avidin-biotin-peroxidase complex method and colour was developed by 3-3’-diaminobenzidine, giving a dark brown signal over the blue-purple counterstaining colour. Original magnification × 100.
In all synovial tissues tPA and PAI-1 immunoreactivity was found in endothelial cells. In addition, a variable expression of tPA and PAI-1, from moderate to strong, was detected in the lining cell area. No apparent difference in distribution was observed in the three groups (N, OA, and RA synovial tissues).
Table 2 summarises the results of the microscopic evaluation of the immunohistochemical staining.
Immunohistochemical staining of components of the plasminogen activation system in normal synovia and synovia of OA and RA patients
uPA, uPAR, tPA, ANDPAI- 1 mRNA EXPRESSION IN NORMAL AND PATHOLOGICAL SYNOVIAL TISSUES
The presence of mRNA for the different components of the PA system was first analysed by northern blot hybridisations (fig 3A). In all samples analysed, single sized uPA, tPA, and uPAR mRNA transcripts, co-migrating with the corresponding mRNA of a human fibrosarcoma cell line (HT 1080, C, last lane of the northern blots) were detected, thus ruling out the presence of alternatively spliced mRNAs (as previously described for uPAR mRNA in gastric mucosa30). As expected from the histological zymograms and immunohistochemistry results, uPA mRNA levels were increased in OA and RA tissues compared with N, although this increase in uPA mRNA levels did not reach statistical differences. By contrast, tPA mRNA levels were significantly decreased about twofold in OA and threefold in RA tissues. The induced uPAR expression detected by immunohistochemistry in RA tissues compared with the very low or undetectable values in OA or N tissues, seemed to be accounted for by increased uPAR mRNA steady state levels in RA tissues (fourfold increase in RA compared with OA and N tissues). Finally we examined also the levels of PAI-1 mRNA. Although pronounced inter-individual variations were observed, PAI-1 mRNA amounts were significantly increased in OA and RA by about eightfold and 18-fold respectively. To identify the tissular distribution of the uPA, tPA, uPAR, and PAI-1 mRNAs, tissue sections of OA and RA were hybridised to the corresponding 32P cRNA probes. In situ zymographies were performed on adjacent sections to localise the corresponding PA activity. In OA tissues, a focal accumulation of uPA mRNA was observed along the synovial lining layer (fig 4A, arrowed). The negative control used was the sense uPA mRNA probe, which gave only backgrounds levels of hybridisation. The localised uPA mRNA was not coupled to detectable uPA mediated caseinolytic activity, as there was no lytic activity along the synovial lining (fig 4A). No signal was detected for uPAR (fig 4A) nor tPA and PAI-1 mRNA (not shown); the failure to detect these transcripts is probably because of a lower copy number of these mRNAs compared with uPA mRNA (as suggested by the differential exposure times of the corresponding northern blots). In contrast, in RA synovial tissues, the distribution of uPA and uPAR mRNA appeared to be similar, these mRNA being mainly localised to the synovial lining layer area (fig 4B, arrowed). uPA and uPAR mRNA accumulations were coupled to a corresponding increase in uPA mediated catalytic activity, as evidenced on histological zymograms by the thin dark line of lytic activity (amiloride sensitive) at the membrane border and by the adjacent areas of lytic activity (amiloride sensitive) in the synovial lining area. In addition a zone of tPA mediated caseinolytic activity was located in the deep layer of the synovial membrane. As for OA, no tPA or PAI-1 mRNA could be detected (not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Comparison of uPA, uPAR, tPA, and PAI-1 mRNA concentrations in N and pathological synovial tissues. (A) Northern blot analysis. Total RNA (12 μg/lane) extracted from normal synovial tissue or from synovial tissue of OA and RA patients was hybridised with the corresponding 32P-cRNA probes. Three representative different samples from three different subjects are shown in each group. A positive control, total mRNA from the human fibrosarcoma HT1080, known to produce uPA, uPAR, tPA, and PAI-1 was included as a control in all northern blots (C, last lane). Exposure times were four hours for the uPA probe, 16 hours for the uPAR probe, and 40 hours for tPA and PAI-1 probes. (B) Densitometric analysis. Laser densitometry was used to quantify mRNA levels seen on northern blots. Results are expressed as % of mRNA concentration measured in normal synovia (mean (SEM), n=6–11). * p<0.05 as determined by Mann-Whitney test.
Discussion
Our investigations show that plasmin mediated proteolytic balance is severely disturbed in human synovial tissues affected by destructive joint diseases (OA and RA), corroborating and extending previous findings mainly based on antigenic levels of uPA, tPA uPAR, and PAI-1 in OA and RA synovial fluids and tissues.15-21 We show here that in normal synovial tissue, the major source of plasmin formation was vascular and resulted from tPA catalysed proteolytic activity produced by endothelial cells. This activity was reduced in arthritic tissues. This could be attributed to the concomitant and opposite regulation of PAI-1, a specific inhibitor of tPA, and tPA itself, in the same cells (that is, endothelial and some synovial lining cells), as evidenced by increased PAI-1 and decreased tPA mRNA levels.
Another striking change we observed in plasmin mediated proteolytic activity in arthritic tissues was the overexpression of uPA mediated enzymatic activity, especially in rheumatoid arthritis tissues. In RA synovial tissue, we have localised, by in situ hybridisation and immunohistochemical studies, uPA and uPAR mRNA and the corresponding antigens to synovial lining cells. In this tissue, up modulation of uPA and its receptor was coupled to a corresponding increase in uPA activity. By contrast, in OA synovial tissue, although we demonstrated the expression of uPA mRNA and its antigen, this expression was not coupled to the presence of uPA activity. This uncoupling could be explained by the undetectable or very low levels of uPAR expression in OA synovial tissues (as shown by in situ hybridisation fig 4A and immunohistochemistry fig 2D). Thus, in the absence of significant amounts of uPAR, uPA catalytic activity would be reduced compared with that of receptor bound uPA10 and, in addition, preferentially secreted in the synovial fluid.
Though the mechanisms underlying dysregulations of uPA, uPAR, and PAI-1 gene expression remain undefined, the local presence of proinflammatory cytokines may account for modulations in uPA, uPAR, and PAI-1 production; proinflammatory cytokines are known to accumulate in RA synovial tissues, and have been shown to modulate PAs production by various synovial cells in vitro.22 In that context, major perturbations of PA mediated proteolytic balance were observed in RA synovial tissues, which exhibit pronounced infiltrations by inflammatory cells and increased cytokines concentrations,1 ,2 ,4 while moderate perturbations of PA mediated proteolysis were detected in OA specimens that are associated with limited histological signs of inflammation.3 ,4
RA is characterised by the proliferation of fibroblasts and synovial cells, constituting the pannus that invade the periarticular cartilage and bone. The rheumatoid pannus is therefore thought to mimic the proliferative and invasive features of neoplasia.31 Indeed, arthritic tissues share molecular features with malignant tumours: changed repertoires of gene expression analogous to those seen in malignant tissues have been found in RA tissues. For example, constitutive expression of c-fos and c-jun has been demonstrated in RA specimens and this expression was maintained in RA synoviocytes through multiples cell passages, in the absence of an inflammatory stimulus.32 In addition, the expression of the ets-2 nuclear oncogene was increased in RA tissues.32Interestingly, several known genes involved in the degradation of the extracellular matrix, such as those encoding stromelysin I, collagenase, and uPA, contain AP-1 (which consists of the heterodimer fos/jun proteins) and ets binding sites in their regulatory flanking regions.33 Thus, the increase of uPA mRNA accumulation observed in RA could be partly accounted for by constitutive AP-1 and ets-2 expression.
The overproduction of components of the PA system—that is, uPA, uPAR, and PAI-1— and the decrease of tPA values in RA tissues, is also reminiscent of findings made in tumoral tissues. Numerous studies have reported increased uPA production in a whole variety of neoplasia and correlations have been drawn between increased uPA expression and invasive properties.27 ,34-36 Conversely, reduced amounts of tPA are frequently encountered in tumoral specimens when compared with their non-malignant counterparts.27 ,34-36
The functional repercussions of a changed plasmin mediated proteolytic balance in arthritic tissues remain to be elucidated. As postulated for malignant tissues, increased uPA mediated proteolytic activity may mediate the degradation of articular tissues and promote joint destruction. In addition, PAs seem to fulfil a wider spectrum of activities than previously suspected. For instance, plasmin has been shown to activate and mobilise latent forms of growth/angiogenic factors.37 ,38 Excessive plasmin formation may consequently influence indirectly the growth and differentiation of the various cellular constituents of the arthritic process. Finally, a role for uPA in neutrophils, macrophages and CD4+ lymphocytes recruitment has been reported in an in vivo model of pulmonary inflammation.39This uPA effect could be, at least partly, independent of uPA proteolytic activity.40 Other uPA effects, observed upon binding of uPA to its receptor and independent of uPA catalytic activity have been reported, such as increased cell adhesion and proliferation.41 ,42 Such uPA effects could be relevant to RA pathogenesis.
The availability of animals genetically deficient in PAs43and plasminogen44 offers the means to clarify the pathogenic contribution of the plasmin mediated proteolytic balance in arthritic processes. Ultimately, it may help design novel therapeutic strategies for these debilitating disorders.
Acknowledgments
We would like to thank Veronique Sciretta for her skilful technical assistance and Dr Mark Ibberson for his help in the preparation of the manuscript.
This work was supported by a grant from the Jean and Linette Warnery foundation and a donation by the Loterie Romande.