Article Text

Abstract
Objective: To investigate the correlation between density of nerve fibres and the presence of BDNF+ cells.
Methods: Densities of nerve fibres and BDNF+ cells were detected by quantitative immunohistochemistry in fresh synovial tissue from 52 patients with RA, 59 with OA, and 26 controls (Co). BDNF was also detected by in situ hybridisation.
Results: Sympathetic nerve fibre density was similar in Co and OA but markedly reduced in RA (p = 0.002), whereas density of substance P positive (SP+) sensory nerve fibres was lower in OA than in Co and RA (p = 0.002). The ratio of sympathetic/SP+ sensory nerve fibre density was highest in OA and Co, followed by RA. The correlation between density of sympathetic nerve fibres and SP+ sensory nerve fibres in OA (R = 0.425, p = 0.001) was strongly positive, had a positive trend in Co (R = 0.243, NS), but was negative in RA (R = −0.292, p = 0.040). In RA and OA tissue the density of BDNF+ cells was high in sublining areas but markedly lower in Co (p = 0.001). BDNF+ cell density correlated positively with the ratio of sympathetic/SP+ sensory nerve fibre density in Co (R = 0.433, p = 0.045) and in OA (R = 0.613, p = 0.015), but not in RA (R = 0.101, NS). Immunohistochemical double staining demonstrated that some macrophages and fibroblasts were positive for BDNF.
Conclusions: The correlation of density of SP+ sensory with sympathetic nerve fibres was positive in Co and OA but negative in RA. BDNF may have a stimulatory role on growth of sympathetic in relation to SP+ sensory nerve fibres in Co and OA, but not in RA.
- APAAP, alkaline phosphatase–anti-alkaline phosphatase
- BDNF, brain derived neurotrophic factor
- NGF, nerve growth factor
- NPY, neuropeptide Y
- OA, osteoarthritis
- RA, rheumatoid arthritis
- SP, substance P
- TH, tyrosine hydroxylase
- brain derived neurotrophic factor
- sensory nerve fibres
- substance P
- sympathetic nerve fibres
- tyrosine hydroxylase
Statistics from Altmetric.com
- APAAP, alkaline phosphatase–anti-alkaline phosphatase
- BDNF, brain derived neurotrophic factor
- NGF, nerve growth factor
- NPY, neuropeptide Y
- OA, osteoarthritis
- RA, rheumatoid arthritis
- SP, substance P
- TH, tyrosine hydroxylase
- brain derived neurotrophic factor
- sensory nerve fibres
- substance P
- sympathetic nerve fibres
- tyrosine hydroxylase
Peripheral sympathetic neurotransmitters and their cofactors such as norepinephrine, neuropeptide Y (NPY), and adenosine inhibit important inflammatory functions such as, for example, production of tumour necrosis factor by monocytes/macrophages,1–4 production of oxygen radicals by neutrophils,5–7 and activity of natural killer cells.8,9,10,11 High concentrations of these neurotransmitters (10−7 to 10−5 mol/l via β-adrenoceptors, Y1 NPY receptors, or A2 adenosine receptors) down regulate many aspects of the innate immune system and, thus the presence of sympathetic nerve fibres should be favourable in the inflamed synovium of patients with rheumatoid arthritis (RA) (reviewed by Straub and Cutolo12). However, the density of sympathetic nerve fibres is dramatically reduced in inflamed RA synovium,13,14 whereas the density of proinflammatory substance P positive (SP+) sensory nerve fibres is increased in the synovium of patients with RA as compared with osteoarthritis (OA).15
The reasons for the imbalance of the density of sympathetic in relation to SP+ sensory nerve fibres is not yet known, but the ratio of neurotrophic factors to nerve repellent factors may have an important role.16,17 We have recently shown that the sympathetic nerve repellent factor semaphorin 3C is markedly up regulated in RA synovium as compared with OA or control tissue; this was not observed for nerve repellent factors of sensory nerve fibres.18 This might lead to a decrease of sympathetic nerve fibres in RA as compared with OA and controls. However, there is no good explanation why the density of sympathetic nerve fibres is higher in OA than in RA or controls. It is unlikely that the repellent factors of the semaphorin group play a part because repellent factors of sensory neurons were not detected in the tissue of all subgroups. On the other hand, changes of expression of neurotrophic factors may be responsible for the observed imbalance of sympathetic and SP+ sensory nerve fibres.
Since the discovery of the original nerve growth factor (NGF) in the 1960s by Levi-Montalcini, other peripherally acting neurotrophins, such as brain derived neurotrophic factor (BDNF), neurotrophin-3, neurotrophin-4, neurotrophin-5, and ciliary neurotrophic factor, have been described. In addition to their canonical role in promoting neuronal survival and growth, these molecules appear to regulate multiple aspects of the development of the nervous system in vertebrates, including neuronal differentiation, axon elongation, and target innervation.19 The growth promoting effects of the above mentioned neurotrophins on sensory or sympathetic neurons are unspecific because their receptors appear on both types of nerve fibres (see for example, Barbacid20 and Chalazonitis21). Interestingly, NGF supports the inflammatory process and it is up regulated during inflammatory episodes in the skin,22 in rheumatoid synovium,23 and in the bronchial system.24 On the basis of its unspecific effects on both types of peripheral nerve fibres, NGF should induce nerve fibre sprouting of sympathetic and sensory nerve fibres. Because only SP+ sensory nerve fibres are increased in the inflamed synovium of patients with RA other mechanisms must be responsible for the observed imbalance.
In contrast with NGF, the role of BDNF in inflammation is not precisely known. BDNF is up regulated in rat dorsal root ganglia after acute peripheral inflammation.25 Activated cells of the immune system express BDNF upon stimulation,26,27 and BDNF has been found in inflammatory lesions in the brain and in the pancreas during pancreatitis.26,28 However, its expression has never been investigated in arthritic animals or in patients with RA or OA.
This study aimed at investigating the presence of BDNF+ cells in synovial tissue of patients with RA, OA, and traumatic controls. Furthermore, we wanted to investigate a possible relationship between the density of BDNF+ cells and the density of sympathetic/SP+ sensory nerve fibres.
PATIENTS AND METHODS
Patients and controls
Fifty two patients with RA and 59 patients with OA who underwent knee joint replacement surgery and 26 control subjects with acute trauma who underwent arthroscopic knee surgery were included without further selection. In the original pool of 30 control subjects, four patients had chronic traumatic joint affections. These latter control subjects had clear signs of inflammation so that they were removed from the analysis. The 26 remaining control subjects had acute disease and no visible signs of inflammation. Diagnosis of RA was based on the established American College of Rheumatology criteria.29 All patients were informed about the purpose of the study and gave written consent. The study was approved by the ethical committee of the University of Regensburg.
Table 1 shows details of the clinical and laboratory data for the entire group and for the subgroup included in the BDNF immunohistochemical study. Respective control subjects (11 women, 15 men) had a mean (SEM) age of 32.2 (2.4) (range 16–60) years. Patients for the subgroup analysis included in the BDNF immunohistochemistry study are shown in braces in table 1. Respective control subjects (2 women, 5 men) had a mean (SEM) age of 41.1 (4.9) (16–60) years. Variables such as erythrocyte sedimentation rate, C reactive protein, and blood leucocyte count were measured by standard techniques. Although our control subjects were markedly younger, we decided to include them in order to present a completely different group without any longstanding inflammation. Furthermore, preliminary correlation analyses indicated that nerve fibre density was not related to age (see also “Results”). This group comprised patients with acute knee joint problems such as tibial plateau fracture, anterior cruciate ligament rupture, and loose bodies in the knee joint.
Basic characteristics of all patients with RA and OA
Synovial tissue preparation and immunohistochemistry
Synovial tissue samples were obtained immediately after opening the knee joint capsule or during arthroscopic surgery (in controls). The tissue for histology was prepared as described previously.14 Synovial tissue pieces of at least 3×3 mm2 were used for histology. Samples intended for alkaline phosphatase–anti-alkaline phosphatase (APAAP) staining were immediately placed in protective freezing medium (Tissue Tek, Sakura Finetek, Zoeterwoude, The Netherlands) and then quick frozen by floating on liquid nitrogen. Tissue samples for the detection of nerve fibres were fixed for 12–24 hours in phosphate buffered saline containing 3.7% formaldehyde, and then incubated in phosphate buffered saline with 20% sucrose for 12–24 hours. Thereafter, tissue was embedded in Tissue Tek and quick frozen. All tissue samples were stored at −80°C.
Histological evaluation was carried out as described previously.14 Briefly, the frozen tissue samples were cut into 5–8 µm thick sections. To determine the number of BDNF+ cells (catalogue No MAB248; R&D Systems, Wiesbaden, Germany) in the synovial tissue of each patient, eight cryosections were investigated using APAAP staining and the number of identified structures was averaged from 17 randomly selected high power fields of view (×400) and expressed per square millimetre. Because we did not have authentic BDNF, we controlled the positive BDNF staining by incubating the tissue with mouse isotype IgG control antibodies which always yielded a negative result. The determination of synovial innervation has been described previously.14 Briefly, six to eight cryosections (5–9 µm thick) of the formaldehyde/sucrose fixed tissue samples were used for immunohistochemistry with a primary antibody against tyrosine hydroxylase (TH+, the key enzyme for norepinephrine production in sympathetic nerve endings, catalogue No MAB318; Chemicon, Temecula, CA, USA) and against SP+ (the key neurotransmitter of SP+ sensory nerve fibres, catalogue No AB1566; Chemicon). An Alexa 546 conjugated secondary antibody (catalogue No A-11030 against mouse IgG, catalogue No A-121010 against rabbit IgG; Molecular Probes, Leiden, The Netherlands) was used to achieve immunofluorescent staining of sympathetic and SP+ sensory nerve fibres (fig 1). The numbers of TH+ sympathetic and SP+ sensory nerve fibres per square millimetre were determined by averaging the number of stained nerve fibres (minimum length 50 µm, determined through a micrometre eyepiece) in 17 randomly selected high power fields of view (×400). We controlled the positive nerve fibre staining by incubating the tissue with polyclonal control antibodies which always yielded a negative result.
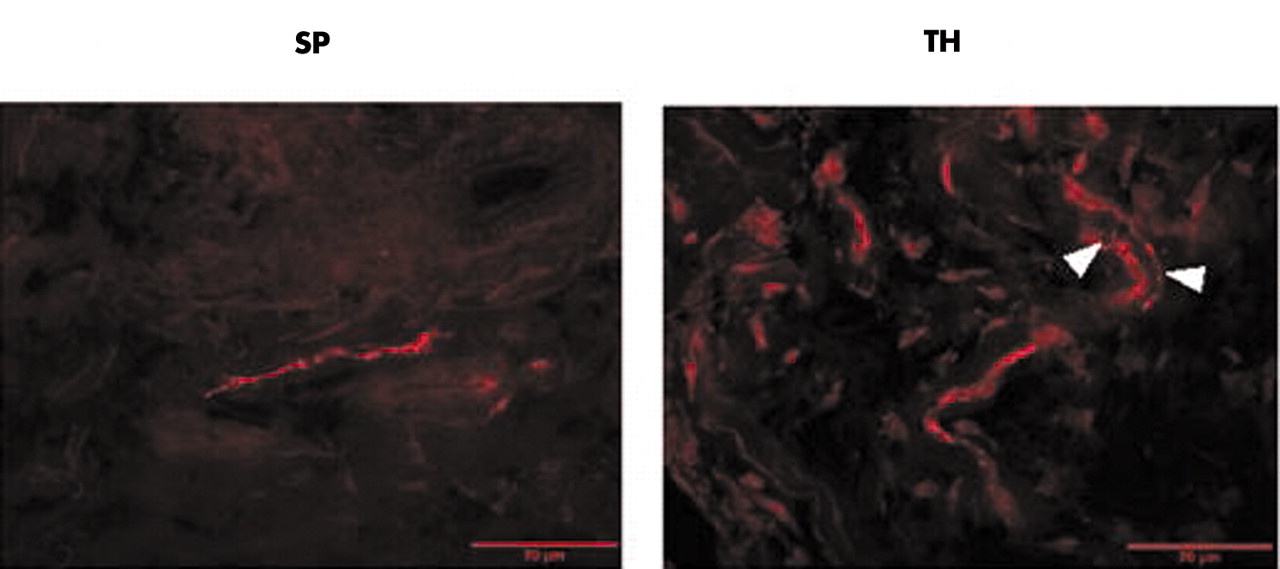
Representative immunohistochemistry of substance P (SP) positive sensory nerve fibres and sympathetic tyrosine hydroxylase (TH) positive nerve fibres. Note the typical juxtaposition of TH positive fibres and the structure of an artery (arrow heads). Micrographs were taken at ×400 magnification.
Immunofluorescence double staining of BDNF+ cells and macrophages/fibroblasts
Double staining of BDNF+ cells and macrophages/fibroblasts was carried out as follows: acetone fixed cryosections were incubated overnight with a polyclonal rabbit antihuman BDNF antibody (catalogue No AB6201-500; Abcam Ltd, Cambridge, UK). Thereafter, fibroblasts and macrophages were stained by mouse monoclonal antibodies against prolyl-4-hydroxylase or CD163 (catalogue No M0877 or M0794; Dako, Glostrup, Denmark). Immunofluorescence staining was achieved with F(ab)2 fragments of goat antirabbit or antimouse IgG (Alexa Fluor 555 or 488, catalogue No. A-21430 or A-11017; Molecular Probes, Leiden, The Netherlands). Nuclei were stained with DAPI (Vectashield mounting medium, catalogue No H-1200; Vector Laboratories, Burlingame, USA).
BDNF in situ hybridisation of synovial tissue
Synovial tissue was stored at −80°C before sectioning (20 µm) on a cryostat. Sections (4–6 per tissue) were mounted onto gelatin coated microscope slides that were stored at −80°C until required. In situ hybridisation histochemistry was carried out as described previously.30 Briefly, the sections were taken through a series of dehydrating alcohol washes, air dried, and a [35S]dATP 3′ end labelled oligonucleotide BDNF probe added in hybridisation buffer. The probe sequence was 5′-CCA-GTG-CCT-CTT-GTC-TAT-GCC-CCT-GCA-GCC-CTC-CTT-TGT-GTA-CCC-CAT-3′, representing nucleotides 746–795.31 The sections were incubated overnight at 37°C before washing to remove unbound and non-specifically bound probe. The sections were dried and exposed to autoradiography film. Further sections were dipped in photographic emulsion to visualise cellular distribution of the probe bound. Counter staining was carried out with haematoxylin and eosin.
Presentation of data and statistical analysis
Data are presented as mean (SEM). Groups were compared by the non-parametric Mann-Whitney test, correlations were calculated by Pearson’s correlation analysis (SPSS/PC, version 11.5, SPSS Inc, Chicago, USA). Frequencies in two different groups were compared by χ2 test using Yates’s continuity correction or Fisher’s exact test if possible. p<0.05 was the level of significance.
We used the ratio “density of sympathetic TH+ nerve fibres/density of SP+ sensory nerve fibres” in order to demonstrate the relation between both types of nerve fibre (values are without units).15 To investigate the interrelation between the density of BDNF+ cells and the density of sympathetic TH+ or SP+ sensory nerve fibres, we used correlation analysis (x value: density of BDNF+ cells; y value: above mentioned nerve fibre ratio) and a ternary plot (x value: density of BDNF+ cells; y value: density of sympathetic TH+ nerve fibres; z value: density of SP+ sensory nerve fibres). A ternary plot is an analytical and plotting application that normalises and plots three values on a triangular diagram and three bivariate cross plots. In the plot, means and standard deviations are calculated in the normalised space (see http://home.c2i.net/astandne/help_htm/english/terplot1.htm; accessed 27 September 2004). The ternary diagram was created using the standard ternary plot software of SigmaPlot 2002 (SPSS Inc, Chicago, USA).
RESULTS
Sympathetic and SP+ sensory innervation of synovial tissue
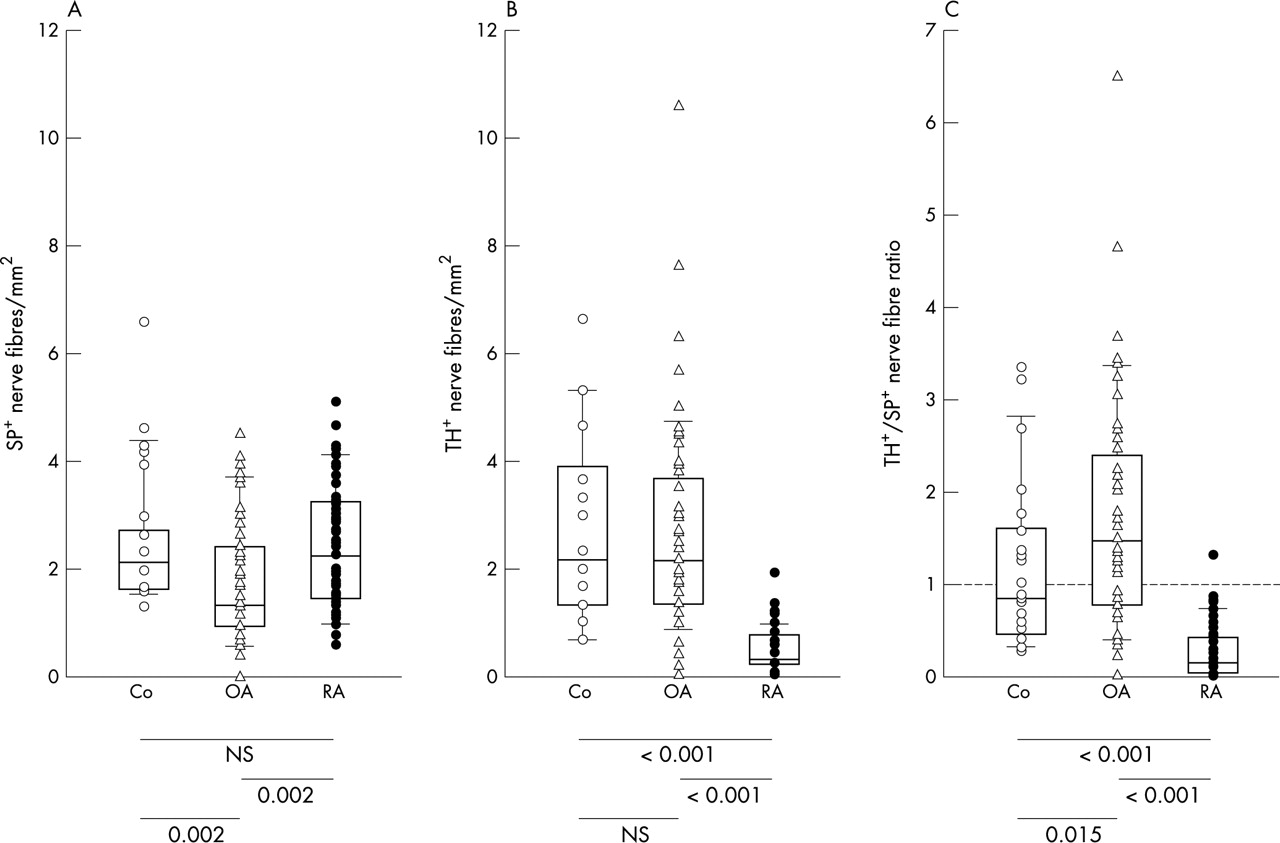
Figure 1 shows representative immunohistochemical stainings of SP+ sensory nerve fibres and sympathetic TH+ nerve fibres. Note the typical pattern of TH+ staining which follows larger arteries (fig 1). A significant reduction of SP+ sensory nerve fibres was found in patients with OA compared with patients with RA or control subjects with non-inflammatory conditions (fig 2A). This indicates a reduction of SP+ sensory innervation in patients with OA. For sympathetic synovial innervation, there was a clear reduction of sympathetic TH+ nerve fibres in patients with RA as compared with patients with OA and controls (fig 2B). No difference was seen between patients with OA and control subjects (fig 2B). The ratio of the density of sympathetic TH+ nerve fibres/SP+ sensory nerve fibres was markedly lower in patients with RA than in patients with OA and controls (fig 2C). Interestingly, patients with OA had a significantly higher ratio than controls (fig 2C), which indicates that patients with OA had the highest density of sympathetic in relation to SP+ sensory nerve fibres when compared with the other two groups.

Density of sympathetic and SP+ sensory nerve fibres in control subjects (Co), patients with OA, and RA. (A) Density of SP+ sensory nerve fibres; (B) density of sympathetic TH+ nerve fibres; and (C) ratio of density of sympathetic/SP+ sensory nerve fibres. Values of p for comparison of group medians are given below the panels. The box plots demonstrate the 10th and 90th centile, the 25th and 75th centile, and the median.
Correlation analysis showed a trend for a positive interrelation between sympathetic TH+ nerve fibres and SP+ sensory nerve fibres in control subjects (fig 3A) and in patients with OA (fig 3B). In patients with RA, this interrelation was negative, which indicates inverse coupling of sympathetic and SP+ sensory nervous inputs to RA synovium (fig 3C).
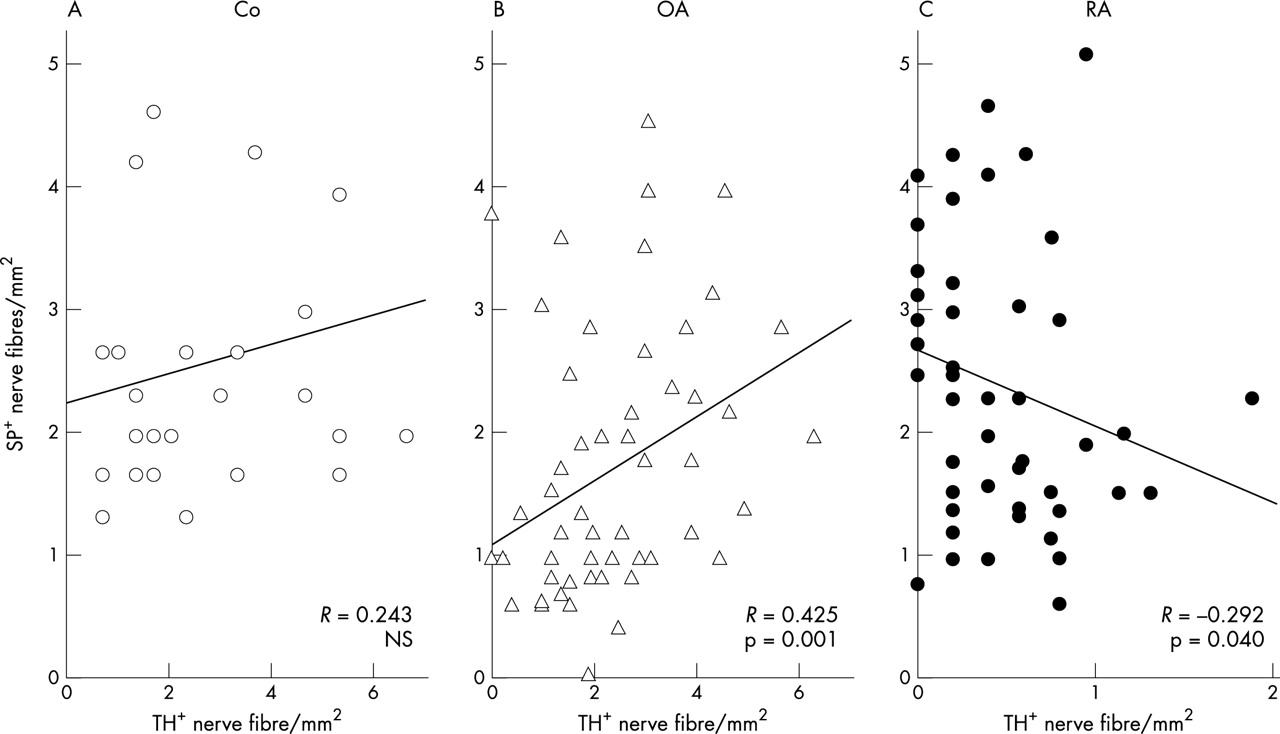
Interrelation between density of sympathetic TH+ nerve fibres and density of SP+ sensory nerve fibres in (A) control subjects (Co); (B) patients with OA; and (C) patients with RA. The linear regression line and its Pearson correlation coefficient are given.
In a receiver operator curve analysis including all investigated subjects, both the density of sympathetic nerve fibres and the ratio of the density of sympathetic nerve fibres divided by the density of SP+ nerve fibres turned out to be excellent variables to separate patients with RA from either patients with OA or control subjects. In this analysis, it was found that a cut off value of 0.94 for the density of sympathetic nerve fibres yielded the highest sensitivity of 92.0%, a specificity of 87.8%, and an accuracy of 89.4%. When the nerve fibre ratio was used as the variable, it was found that a cut off value of 0.50 yielded the highest sensitivity of 84.0%, a specificity of 85.4%, and an accuracy of 84.8%.
Because the control patients were markedly different in age, we investigated whether an interrelation existed between age and nerve fibre density in the various patient subgroups: no such correlation was found for control subjects (age v TH+: R = 0.192, p = 0.348; age v SP+: R = −0.056, p = 0.785), patients with OA (age v TH+: R = 0.243, p = 0.069; age v SP+: R = 0.127, p = 0.342), and patients with RA (age v TH+: R = −0.090, p = 0.525; age v SP+: R = 0.158, p = 0.273).
In another analysis we compared women and men in order to investigate a possible difference in nerve fibre density. We detected no difference for density of sympathetic TH+ nerve fibres and SP+ sensory nerve fibres between women and men in the different patient groups (p values at least >0.20, data not shown).
BDNF+ cells in synovial tissue
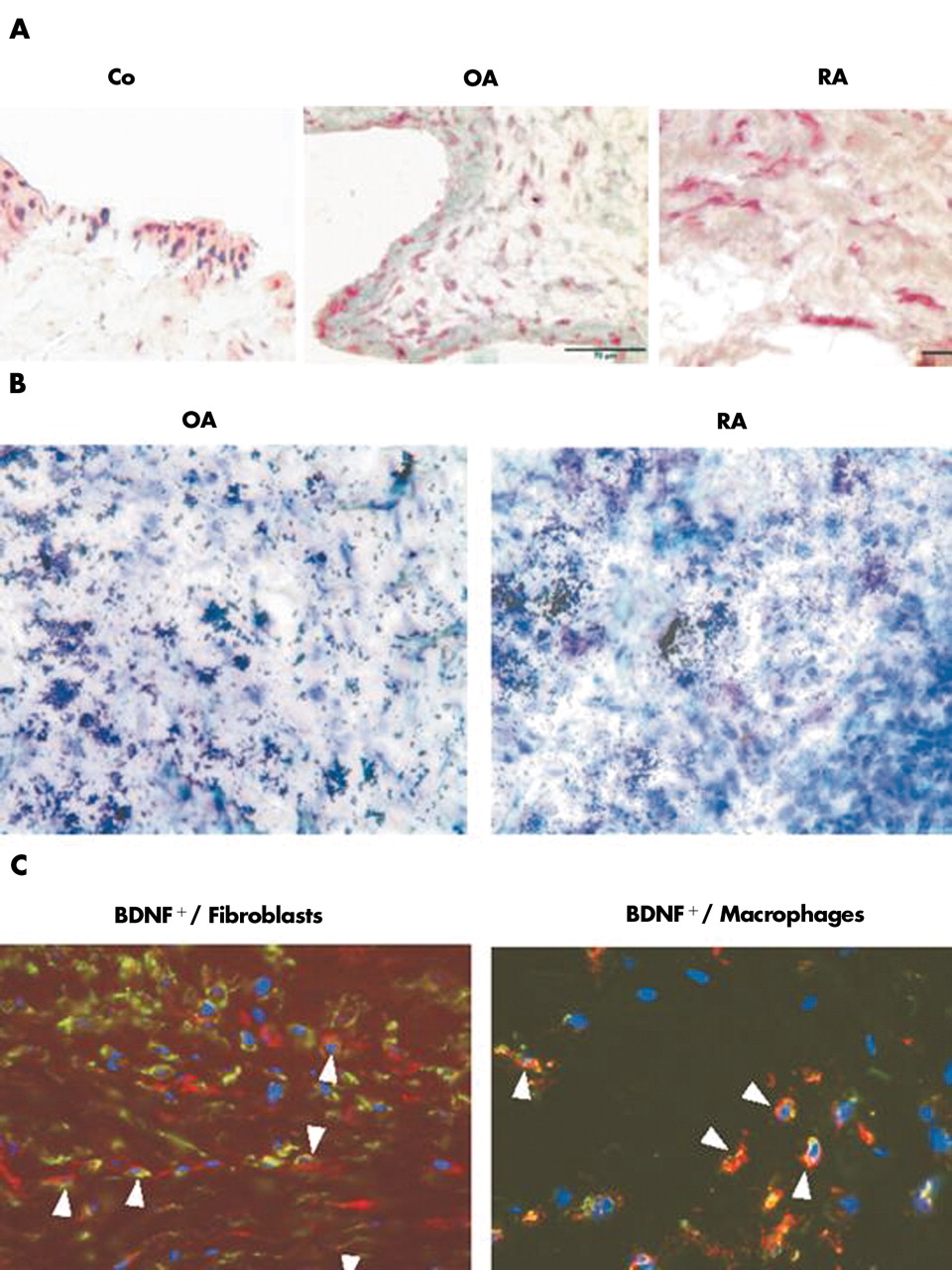
Neurotrophic factors such as BDNF may potentially influence the growth of sympathetic and SP+ sensory nerve fibres in a very differential way (supporting the growth of one, inhibiting the other). To link the expression of BDNF to sympathetic and SP+ sensory nerve fibres, BDNF was detected immunohistochemically: BDNF+ cells were found in all three patient groups (fig 4A). In all three patient groups, BDNF cells were located in the lining area (fig 4A). However, only in patients with RA and OA were the cells in the synovial sublining area stained positive for BDNF (fig 4A). A representative in situ hybridisation of OA and RA synovial sublining tissue demonstrated marked expression of BDNF mRNA (fig 4B). Immunohistochemical double staining confirmed that BDNF+ cells most often stained positive also for markers of fibroblasts (prolyl-4-hydroxylase) and macrophages (CD163), respectively (fig 4C). Furthermore, there was a highly significant increase in the density of BDNF+ cells in patients with RA and OA in comparison with control subjects (fig 5). This indicates that BDNF is particularly expressed under more inflammatory conditions. In the different groups, age and sex were not related to the density of BDNF+ cells (data not shown).

Immunohistochemistry of BDNF in synovial tissue. (A) Synovial tissue from a control subject without an inflammatory condition (Co), a patient with OA, and a patient with RA stains positive for BDNF. APAAP staining was used. (B) BDNF in situ hybridisation of synovial tissue from a patient with OA and RA. (C) Immunofluorescence double staining of BDNF+ cells (green fluorescence) versus fibroblasts (left panel: red fluorescence, prolyl-4-hydroxylase) or macrophages (right panel: red fluorescence, CD163). Nuclei were stained with DAPI. In all panels, respective control stainings were always negative. All micrographs were taken at ×400 magnification.

Density of BDNF+ cells in control subjects (Co), patients with OA, and patients with RA. Values of p for comparison of group medians are given.
Interrelation between BDNF+ cells and sympathetic TH+ or SP+ sensory nerve fibres
A correlation analysis for the density of BDNF+ cells and the nerve fibre ratio demonstrated a positive interrelation in control subjects and patients with OA (figs 6A and B), but no such relationship existed in patients with RA (fig 6C). In controls and patients with OA this indicates that increased density of BDNF+ cells is linked to a higher ratio of sympathetic nerve fibres to SP+ sensory nerve fibres.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Interrelation of the density of BDNF+ cells and the ratio of the density of sympathetic TH+ nerve fibres/the density of SP+ sensory nerve fibres in (A) control subjects (Co); (B) patients with OA; and (C) patients with RA. The linear regression line and its Pearson correlation coefficient are given. (D) Ternary plot of the density of BDNF+ cells, the density of sympathetic TH+ nerve fibres, and the density of SP+ sensory nerve fibres. A ternary plot is an analytical tool that normalises and plots three values on a triangular diagram. In the plot, means and standard deviations are calculated in the normalised space (SigmaPlot 2002).
In the ternary plot, control subjects clearly differed from patients with RA and OA. This is demonstrated by the lower density of BDNF+ cells and higher density of sympathetic TH+ nerve fibres (fig 6D, open circles). Patients with RA and some patients with OA had a high density of BDNF+ cells and very low density of sympathetic TH+ nerve fibres (fig 6D, red circles and black triangles in lower left corner). A subgroup of patients with OA exists who had an intermediate pattern (fig 6D, black triangles, between lower left corner and control subjects).
DISCUSSION
This is the first study which has demonstrated similar synovial density of sympathetic nerve fibres in a control group without inflammation in comparison with patients with OA. This analysis was important in order to give an unbiased absolute measure of the density of sympathetic nerve fibres in non-inflamed synovial tissue. In addition, we confirmed the striking loss of sympathetic nerve fibres in patients with RA14,15 as compared with patients with OA and control subjects.
In animal studies, reduction of nerve fibre density was dependent on the stage of the disease.32 However, we were unable to link reduction of sympathetic nerve fibres to markers of inflammation or disease duration. Our patients had long term disease, and it seems that in this stage of the disease sympathetic nerve fibres are almost absent. In control subjects and patients with OA, the densities of sympathetic and SP+ sensory nerve fibres were positively correlated, whereas in patients with RA this correlation was negative.
Because high levels of sympathetic neurotransmitters inhibit many immunological functions, particularly, those of the innate immune system,1,2,3,4,5,6,7,8,9,10,11 a normal density of sympathetic nerve fibres of about 2.6 nerve fibres/mm2 may be necessary for a balanced immune reaction ( = the value of the control subjects). This is particularly important because the density of proinflammatory SP+ sensory nerve fibres remains unaltered.14,15 It is important to mention that low concentrations of norepinephrine (<10−7 mol/l, via α-adrenoceptors) and adenosine (<10−7 mol/l, via A1 adenosine receptors) can exert a completely opposite, now proinflammatory influence on the immune system (summarised by Straub and Cutolo12). The equilibrium, as estimated by the ratio of the density of sympathetic to the density of SP+ sensory nerve fibres in control subjects and patients with OA, is typically above 0.50–1.00. We may speculate that under normal conditions this equilibrium is maintained in order to establish a balanced tissue supply. This balance is largely shifted to proinflammatory SP+ sensory nerve fibres in patients with RA. At this point the question arises why, in this inflammatory process, the density of sympathetic nerve fibres decreases and the density of SP+ sensory nerve fibres remains constant.
The presence of nerve fibres in peripheral tissue is controlled by nerve growth factors and nerve repellent factors. It has been repeatedly demonstrated that NGF is present in inflamed synovial tissue and fluid in RA,23 in circulating blood,33 and in the synovial fluid of patients with spondyloarthropathies.34 NGF is produced by different leucocytes upon stimulation (see, for example, Pezzati et al35 and Morgan et al36), and was found to be an important stimulator of wound healing.37–39 In contrast with NGF, the presence of BDNF has never been described in synovial tissue in animals or men. BDNF is up regulated in rat dorsal root ganglia after acute peripheral inflammation.25 Activated cells of the immune system express BDNF,26,27 and BDNF was found in inflammatory lesions.26,28 Both BDNF and NGF are up regulated in inflammatory lesions. Our study demonstrated increased density of BDNF+ cells in inflamed synovium of patients with RA and OA but not in control subjects. The difference was most pronounced in the sublining area, and both fibroblasts and macrophages stained positive for BDNF.
Moreover, we tried to link cellular expression of BDNF to the described dissociation of sympathetic versus SP+ sensory nerve fibres. The analysis showed that the density of BDNF+ cells is positively related to the increased density of sympathetic in relation to SP+ sensory nerve fibres in control subjects and patients with OA, but not in patients with RA. One may speculate that BDNF supports the growth of sympathetic nerve fibres more than the growth of SP+ sensory nerve fibres, which may be disrupted in patients with RA. As patients with RA in comparison with OA and controls demonstrate increased density of the sympathetic nerve repellent factor semaphorin 3C in synovial tissue,18 the presence of BDNF may not influence the growth of sympathetic nerve fibres in the same way as in controls and patients with OA.
Although the deductive nature of our approach does not allow us to find a causative relationship between BDNF and nerve fibre dissociation, it stimulates new ideas for following up this intriguing subject in experimental models of arthritis. In addition, the decrease of visible nerve fibres may also depend on exaggerated release of neuropeptides or alterations of expression of key enzymes such as TH during the inflammatory process.16,17 Nevertheless, it seems obvious that the detected loss of visible nerve fibres is probably linked to functional alterations of these neuronal pathways.
A second interesting finding was the decreased density of SP+ sensory nerve fibres in patients with OA as compared with control subjects and patients with RA. At present, we do not know how the loss of SP+ sensory nerve fibres influences the osteoarthritic process but one may speculate that loss of joint sensation would lead to continuous mechanical microtraumas.40,41 Furthermore, loss of SP may lead to a lack of growth promoting factors because SP can stimulate extracellular matrix production etc.42–44 Both conditions may lead to a loss of joint protection in patients with OA.
In conclusion, the interrelation of the density of SP+ sensory and sympathetic nerve fibres was positive in control subjects and OA but negative in RA, which demonstrates inverse coupling of these two nervous systems in RA synovium. BDNF, a neurotrophic factor, may have a stimulatory role on the growth of sympathetic in relation to SP+ sensory nerve fibres in control subjects and OA, which may not be the case in RA. Further studies will be needed to prove whether BDNF is a relevant cofactor for the observed dissociation of sympathetic versus SP+ sensory nerve fibres in animal models of arthritis.
Acknowledgments
Part of this study were funded by the Deutsche Forschungsgemeinschaft (Str 511/10-1) and by the respective institutions.
REFERENCES
Footnotes
-
↵* C Weidler and C Holzer contributed equally to this study.