Article Text

Abstract
Objectives Methotrexate (MTX) functions as an antiproliferative agent in cancer and an anti-inflammatory drug in rheumatoid arthritis (RA). Although macrophages critically contribute to RA pathology, their response to MTX remains unknown. As a means to identify MTX response markers, we have explored its transcriptional effect on macrophages polarised by GM-CSF (GM-MØ) or M-CSF (M-MØ), which resemble proinflammatory and anti-inflammatory macrophages found in RA and normal joints, respectively.
Methods The transcriptomic profile of both human macrophage subtypes exposed to 50 nM of MTX under long-term and short-term schedules were determined using gene expression microarrays, and validated through quantitative real time PCR and ELISA. The molecular pathway involved in macrophage MTX-responsiveness was determined through pharmacological, siRNA-mediated knockdown approaches, metabolomics for polyglutamylated-MTX detection, western blot, and immunofluorescence on RA and normal joints.
Results MTX exclusively modulated gene expression in proinflammatory GM-MØ, where it influenced the expression of 757 genes and induced CCL20 and LIF at the mRNA and protein levels. Pharmacological and siRNA-mediated approaches indicated that macrophage subset-specific MTX responsiveness correlates with thymidylate synthase (TS) expression, as proinflammatory TS+ GM-MØ are susceptible to MTX, whereas anti-inflammatory TSlow/− M-MØ and monocytes are refractory to MTX. Furthermore, p53 activity was found to mediate the TS-dependent MTX-responsiveness of proinflammatory TS+ GM-MØ. Importantly, TS and p53 were found to be expressed by CD163+/TNFα+ GM-CSF-polarised macrophages from RA joints but not from normal synovium.
Conclusions Macrophage response to MTX is polarisation-dependent and determined by the TS-p53 axis. CCL20 and LIF constitute novel macrophage markers for MTX responsiveness in vitro.
- Methotrexate
- Rheumatoid Arthritis
- DMARDs (synthetic)
Statistics from Altmetric.com
Introduction
Methotrexate (MTX) is currently used for treatment of a number of malignant diseases including acute lymphoblastic leukaemia and osteosarcoma.1 ,2 As a competitive inhibitor of dihydrofolate reductase (DHFR), MTX suppresses tetrahydrofolate (THF)-dependent reactions primarily by depletion of cellular folate cofactors.3 MTX is polyglutamylated by the enzyme folylpolyglutamate synthetase (FPGS) increasing their retention within the cells. Importantly, polyglutamates of MTX (MTX-Pgl) are long-lived active compounds and potent inhibitors of thymidylate synthase (TS) and aminoimidazole-carboxamide-ribonucleoside transformylase (AICARTF or ATIC), enzymes involved in de novo synthetic pathway of pyrimidines and purines required for DNA replication and repair.3 This mechanism of action is responsible for the success of MTX as an antimalignancy drug. Beyond its use as an antiproliferative agent, MTX has long been used as an anti-inflammatory agent for the treatment of autoimmune diseases such as rheumatoid arthritis (RA) and psoriasis.4 Although the precise mechanism for the therapeutic action of MTX on these pathologies remains unclear, release of adenosine, inhibition of polyamines and generation of reactive oxygen species though JNK activation have been reported to mediate the anti-inflammatory action of MTX in RA.5–7 Recent studies indicate that MTX treatment restores the expression of altered folate metabolism genes during RA.8 Importantly, different doses of MTX are administered for the treatment of different pathologies: high-dose MTX therapy (5000 mg/week) is used for malignant diseases whereas low doses of MTX are used for autoimmune diseases (5–25 mg/week).5
Macrophages play an important role in the sequence of events between chronic inflammation/tumorigenesis and tumour progression.9 ,10 The contribution of macrophages to tumour development could be inferred from the impaired tumour progression seen in animals with altered macrophage differentiation.11 ,12 Macrophages are also critical cellular components in RA pathogenesis since increased numbers of macrophages in the synovial tissue correlate with the degree of joint erosion.13–15 Macrophages are key cells of the innate immune response and fulfil a variety of effector functions depending on the activation of specific signalling pathways, transcription factors and their metabolic adaptation, all of which underlie macrophage plasticity and polarisation.16–18 In vitro, a well established method for generating proinflammatory and anti-inflammatory macrophages is the differentiation of monocytes in the presence of GM-CSF or M-CSF.19 In response to TLR ligands, GM-CSF-driven macrophages produce proinflammatory tumour necrosis factor alpha (TNFα) and IL-23 and promote type 1 immunity, whereas M-CSF-derived macrophages produce IL-10, inhibit Th1 responses and exhibit regulatory properties.12 ,20 ,21 Therefore, GM-CSF-and M-CSF-polarised macrophages are considered as proinflammatory (GM-MØ) and anti-inflammatory (M-MØ) macrophages, respectively.
Whereas MTX has a clear antiproliferative action on tumour cells, its effect on proinflammatory and anti-inflammatory macrophages remains undefined. Therefore, we decided to determine the MTX-dependent gene expression profile in macrophages. We explored the role of MTX in long-term and short-term schedules in peripheral blood monocytes, GM-MØ and M-MØ, and found evidences for the context-dependent effect of MTX on human myeloid cells. Moreover, we found a series of novel markers for MTX responsiveness in proinflammatory GM-CSF polarised-macrophages.
Methods
Detailed methods are supplied in the online supplementary file.
Supplemental material
Human peripheral blood mononuclear cells (PBMCs) were isolated from buffy coats from normal donors. Monocytes were purified from PBMCs by magnetic cell sorting using CD14 microbeads (Miltenyi) and were cultured at 0.5×106 cells/mL for 7 days in RPMI supplemented with 10% fetal calf serum, containing GM-CSF (1000U/mL) or M-CSF (10 ng/mL, ImmunoTools) to generate GM-CSF-polarised macrophages (GM-MØ) or M-CSF-polarised macrophages (M-MØ). For long-term MTX treatment, independent preparations of monocytes unexposed or exposed to MTX (50 nM) and differentiated to GM-MØ or M-MØ for 7 days were analysed by gene expression profiling, quantitative real time PCR (qRT-PCR), immunoblotting or ELISA. For short-term schedule, independent samples of GM-MØ or M-MØ unexposed or exposed to MTX for 48 h were analysed by gene expression profiling, qRT-PCR, immunoblotting or immunofluorescence.
Results
Identification of the gene profile of monocytes exposed to long-term MTX
Weekly administered MTX is the most important DMARD for the treatment of RA.4 ,22 To determine the influence of MTX on macrophage differentiation, monocytes were exposed to a single dose of 50 nM MTX immediately before the initiation of the GM-CSF or M-CSF differentiation process and gene expression changes were analysed at the end of the 7 day culture (figure 1A). No effect of MTX on cell viability was observed (not shown). This long-term MTX treatment resulted in the generation of a transcriptional profile with 547 upregulated and 210 downregulated genes in GM-CSF-exposed monocytes. By contrast, MTX did not modify the transcriptional profile of M-CSF-polarised macrophages (figure 1B). This result indicates that the effect of MTX on macrophages is critically conditioned by the cytokine context (GM-CSF vs M-CSF). Interestingly, Canonical Pathways analysis in Ingenuity Pathways Analysis (IPA) software indicated that MTX significantly regulates the expression of genes involved in inflammatory responses (‘Granulocyte Adhesion and Diapedesis’), immune responses (‘Communication between Innate and Adaptive Immune Cells’) and inflammation resolution (‘PPAR signalling’) (figure 1C, see online supplementary table S1). Moreover, DiseaseBioFunction analysis in IPA revealed that MTX significantly modulates the expression of genes under the terms ‘Inflammatory Response’ (p=1.28×10−16) and ‘Connective Tissue Disorders, Skeletal and Muscular Disorders’ (p=2.99×10−15), further supporting the ability of MTX to modulate the expression of genes associated with inflammation and RA (see online supplementary table S1).
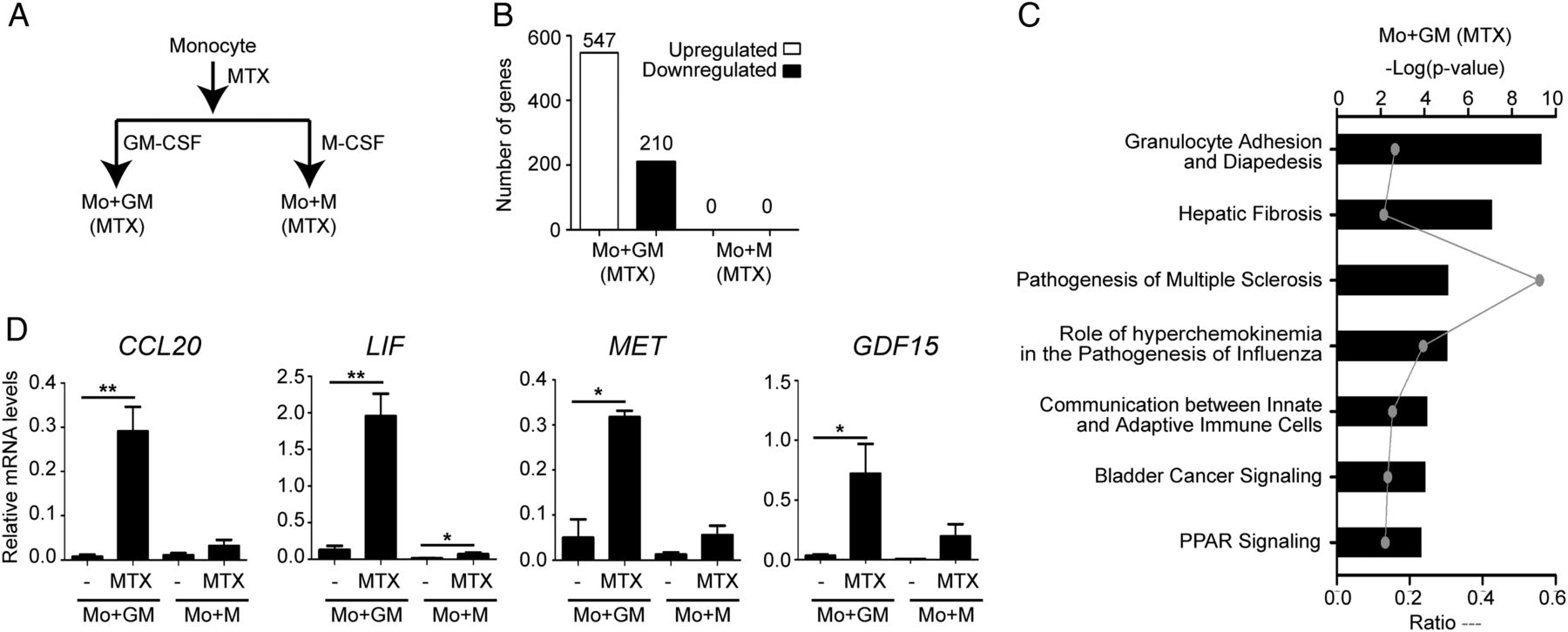
Determination of the long-term methotrexate (MTX)-dependent transcriptomic profile of monocytes and validation of the transcriptomic data. (A) Schematic representation of the experiments. MTX (50 nM) was added once on monocytes and mRNA levels were determined at day 7 of the GM-CSF (Mo+GM) or M-CSF (Mo+M)-induced differentiation. (B) Quantification of the number of genes whose expression is significantly altered (adjusted p<0.05) on MTX-treated monocytes after 7 days of the GM-CSF (Mo+GM) or M-CSF (Mo+M)-induced differentiation. (C) Analysis for canonical pathways significantly enriched within the sets of genes whose expression is altered by MTX exposure in Mo+GM. (D) CCL20, LIF, MET and GDF15 mRNA expression levels determined by qRT-PCR on monocytes differentiated with GM-CSF (Mo+GM) or M-CSF (Mo+M) in the absence or in the presence of MTX. Mean and SEM of six independent donors are shown (*p<0.05, **p<0.01).
Validation of the transcriptomic data and kinetics of MTX response during macrophage differentiation
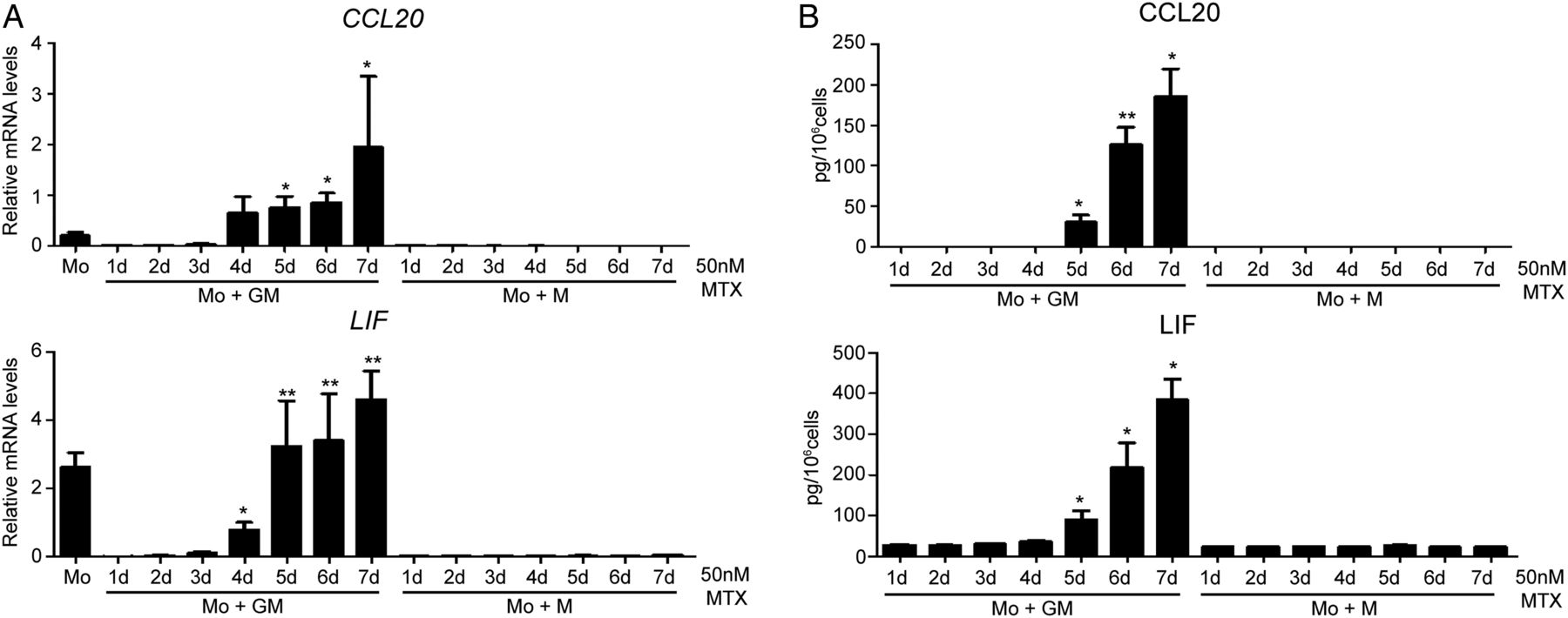
MTX-treated monocytes led to the generation of macrophages with a significantly increased CCL20, LIF, MET and GDF15 mRNA expression and decreased CCR2 mRNA (genes associated with processes like ‘Inflammatory Response’ and ‘Rheumatic Disease’, see online supplementary tables S1 and S2). To verify the transcriptomic data, monocytes from different donors were exposed to MTX and differentiated into GM-MØ and M-MØ. Long-term MTX treatment resulted in significantly enhanced expression of CCL20 (158-fold), LIF (25-fold), MET (45-fold) and GDF15 (24-fold) mRNA in GM-CSF cells, thus validating the transcriptomic data (figure 1D). In the next experiments we focused on CCL20 and LIF as representative MTX-response genes in macrophages. Kinetic analysis revealed that CCL20 and LIF mRNA and protein expression significantly increased 4–5 days after MTX treatment and the protein reached maximum levels after 7 days during GM-CSF macrophage differentiation (figure 2A, B). Altogether, these results validate the data from the global analysis of gene expression and indicate that CCL20 and LIF constitute novel markers for MTX responsiveness in GM-MØ in vitro.

Methotrexate (MTX) induces CCL20 and LIF expression on day 5 of the GM-CSF differentiation. CCL20 (upper panel) and LIF (lower panel) expression levels determined by quantitative real-time PCR (A) or ELISA (B) on monocytes differentiated with GM-CSF or M-CSF in the absence or in the presence of 50 nM of MTX. MTX was added once on monocytes and mRNA (A) or protein (B) levels were measured at the indicated time points along the GM-CSF (Mo+GM-CSF) or M-CSF (Mo+M-CSF)-induced differentiation. For (A), results are expressed as relative mRNA levels, which indicates the expression of each gene in MTX-treated cells relative to its expression in untreated cells at every time point (d, days). Mean and SEM of four independent donors are shown (*p<0.05, **p<0.01 compared with the level detected in non-stimulated macrophages at every time point). For B, results indicate the expression of each cytokine in MTX-treated cells relative to its expression in untreated cells at every time point. Mean and SEM of four independent donors are shown (*p<0.05, **p<0.01 compared with the level detected in non-stimulated macrophages at every time point).
Short-term exposure of monocytes and macrophages to MTX
In order to explore the molecular basis for MTX response, monocytes and 7-day differentiated GM-MØ and M-MØ were exposed to MTX for a short period (24–48 h) and the expression of MTX-response genes was analysed by qRT-PCR. MTX significantly increased CCL20, LIF, MET and GDF15 mRNA in GM-MØ after 24 h, an effect further enhanced after 48 h (figure 3A, see online supplementary figure S1A). By contrast, short-term exposure to MTX did not significantly modify gene expression either in monocytes or in M-MØ. Interestingly, no induction of these genes was observed in MTX-exposed fibroblast or HEK 293T cells (see online supplementary figure S1B). These results indicate a different MTX responsiveness in monocytes and macrophages and support the context-dependent and differentiation-dependent effect of MTX in myeloid cells.

Short-term exposure of monocytes and macrophages to methotrexate (MTX). (A) CCL20 and LIF mRNA expression levels determined by quantitative real-time PCR (qRT-PCR) on monocytes (Mo), GM-MØ or M-MØ in the absence or the presence of 50 nM of MTX for the indicated time points. Mean and SEM of four independent donors are shown (*p<0.05). (B) Key components of the folate/MTX metabolism with emphasis in the thymidylate biosynthesis pathway. (C) MTX (MTX-Glu1) and MTX polyglutamates (MTX-Glu(2–5)) were quantified by UHPLC in GM-MØ and M-MØ exposed to 50 nM of MTX during 48 h. Mean and SEM of six independent donors are shown (*p<0.05). (D) The expression of the folate/MTX metabolising enzymes FPGS, GGH, DHFR, TYMS, GART, ATIC and MTHFR was analysed by qRT-PCR on monocytes (Mo), GM-MØ (GM) and M-MØ (M). Mean and SEM of six independent donors are shown (*p<0.05, **p<0.01, ***p<0.001). (E) Immunoblot analysis of thymidylate synthase (TS) in lysates of HEK 293T cells transfected with pcDNA3 or TS-pcDNA3. Vinculin protein levels were determined as a loading control. (F) Immunoblot analysis of TS in lysates of monocytes (Mo), GM-MØ (GM) or M-MØ (M). Vinculin protein levels were determined as a loading control. (G) Immunofluorescence analysis of monocytes (Mo), GM-MØ and M-MØ, as determined by confocal microscopy after using a TS-specific antibody. Nuclei were counterstained with DAPI. The experiment was done in five independent donors, and a representative experiment is shown. Scale bars: 50 µm.
The uptake of MTX was similar in GM-MØ and M-MØ, indicating that the differential responsiveness to MTX is not caused by a different expression of folate transporters (see online supplementary figure S2).23 Measurement of the intracellular concentration of MTX-Pgl showed that accumulation of MTX-Glu2 and MTX-Glu3 was higher in GM-MØ than in M-MØ, whereas similar levels of MTX-Glu1, MTX-Glu4 and MTX-Glu5 were found in both macrophage subtypes (figure 3C). Since MTX and MTX-Glu(2–5) inhibit various enzymes involved in one-carbon metabolism (DHFR, TS, ATIC) (figure 3B),3 we next analysed the expression of MTX-metabolising enzymes (figure 3D). Similar levels of DHFR mRNA were found in monocytes, GM-MØ and M-MØ, whereas FPGS and MTHFR mRNA expression was significantly higher in monocytes than in macrophages and ATIC mRNA levels differed between GM-MØ and M-MØ (figure 3D). By contrast, TS (TYMS) expression significantly differed between the three cell populations and its expression was found to be higher in GM-MØ than in M-MØ or monocytes both at the mRNA and protein levels (figure 3D–G). This result suggested that the different MTX responsiveness of GM-MØ and M-MØ could be derived from their distinct TS content, a hypothesis that was tested subsequently.
TS expression conditions MTX-responsiveness in macrophages
The differential expression of TS in monocytes and macrophages led us to explore the role of this enzyme in MTX responsiveness by altering its function or expression. The antifolates pemetrexed (PMX) and raltitrexed (ZD1694), potent TS inhibitors,24 significantly increased CCL20, LIF, MET and GDF15 mRNA in GM-MØ (figure 4A, see online supplementary figure S3A), indicating that the inhibition of TS activity contributes to antifolate (MTX, PMX, ZD1694) responsiveness in GM-MØ. Next, we determined the changes in gene expression induced by MTX or PMX in the presence of thymidine in GM-MØ. Supplementation with thymidine significantly diminished the upregulation of CCL20 and LIF mRNA in MTX or PMX-exposed GM-MØ (figure 4A). To fully confirm that MTX-triggered gene expression changes were dependent on TS inhibition, we determined the effect of silencing TS expression in GM-MØ. Knockdown of TS using two different small interfering RNA significantly increased CCL20 and LIF mRNA levels (figure 4B) whereas SLC19A1 mRNA levels were kept constant and TYMS mRNA expression diminished, thus suggesting that the effect of MTX on CCL20 and LIF gene expression depends on its capacity to inhibit TS. Moreover, MTX-triggered and PMX-triggered gene induction significantly diminished (70–80%) when silencing TS in GM-MØ (figure 4C, D), thus confirming the above hypothesis.

Thymidylate synthase (TS) expression conditions methotrexate (MTX) responsiveness in macrophages. (A) CCL20 and LIF mRNA expression levels determined by quantitative real-time PCR (qRT-PCR) on GM-MØ treated with MTX or pemetrexed (PMX) in the absence or the presence of thymidine (dT) for 48 h. Mean and SEM of eight independent donors are shown (*p<0.05, **p<0.01). (B) Gene expression determined by qRT-PCR on GM-MØ transfected with control siRNA (c) or two different siRNA for TYMS (si8, si9). The values in the presence of control siRNA were given an arbitrary value of 1 (*p<0.05, ***p<0.001). (C) CCL20 and LIF mRNA expression levels determined by qRT-PCR on GM-MØ transfected with control siRNA (c) or siRNA for TYMS (si8, si9) and exposed to MTX (left) or PMX (right) for 48 h. Results are expressed as % induction, and the values in the presence of control siRNA were given an arbitrary value of 100. Mean and SEM of eight (MTX) or five (PMX) independent donors are shown (*p<0.05, **p<0.01, ***p<0.001). (D) Immunoblot analysis of TS and GAPDH in GM-MØ transfected with siRNA (c) or siRNA for TYMS (si8) in the absence or the presence of MTX for 48 h in two donors. (E) TYMS mRNA expression levels determined by qRT-PCR on monocytes differentiated with GM-CSF or M-CSF in the presence of MTX. Mean and SEM of four independent donors are shown (*p<0.05, compared with the level detected in GM-CSF-differentiated monocytes in day 1). Immunoblot analysis of TS and GAPDH in monocytes differentiated with GM-CSF in the presence of MTX at the indicated time points. (F) MTX (MTX-Glu1) and MTX polyglutamates (MTX-Glu(2–5)) were quantified by UHPLC in MTX-treated monocytes after 7 days of the GM-CSF (Mo+GM) or M-CSF (Mo+M)-induced differentiation. Mean and SEM of six independent donors are shown (*p<0.05, **p<0.01 compared with the level of MTX-Glu1 in Mo+GM or in Mo+M and $p<0.05, compared MTX-Glu(2–3) between Mo+GM and Mo+M).
Lastly, we found a correlation between TS expression during macrophage differentiation and MTX responsiveness. Kinetic studies revealed that TYMS mRNA and TS protein are initially detected 3 days after GM-CSF addition (figure 4E), whereas MTX-response (CCL20 and LIF upregulation) begins on day 4 (figure 2A). Conversely, the expression of other folate/MTX metabolising enzymes is essentially kept constant along GM-CSF-driven and M-CSF-driven macrophage differentiation (see online supplementary figure S4). Besides, accumulation of MTX-Glu2 and MTX-Glu3, greater TS inhibitors than MTX-Glu1,25 was higher in proinflammatory than in anti-inflammatory macrophages (figure 4F). Altogether, these results confirm that MTX responsiveness on macrophages depends on TS expression levels.
TS expression on macrophages from RA joints
Previously, we have shown that macrophages from RA joints exhibit a GM-CSF-dependent transcriptomic and phenotypical proinflammatory polarisation state.26 As macrophages represent an important MTX target in RA, the expression of TS was determined in normal and active RA synovium. TS expression was found in CD163+/TNFα+ macrophages within the sublining of RA synovial membranes (figure 5A, B). By contrast, CD163+ macrophages in the normal synovium lacked TS and TNFα expression. Moreover, TNFα treated macrophages maintained TYMS expression and MTX responsiveness, an effect not observed with other proinflammatory (IFN-γ) or anti-inflammatory (IL-4) stimuli (figure 5C). These results indicate that macrophages from RA, and not from normal joints, represent a cellular target for MTX.

Expression of thymidylate synthase (TS) by macrophages from rheumatoid arthritis (RA) joints and methotrexate (MTX)-induced gene expression in the presence of TNFα or folic acid. (A) Immunofluorescence analysis of normal and active RA synovial tissues, as determined by confocal microscopy using CD163-specific, TS-specific and TNFα-specific antibodies. Nuclei were counterstained with DAPI. The experiment was done on six independent samples and a representative experiment is shown. Scale bars: 100 µm. Magnification (20×) of the areas indicated by a dotted square in active RA synovial tissue. (B) Quantification of TS and TNFα staining in CD163 macrophages from normal (NS) and active RA synovial tissue (RA) (***p<0.001). (C) CCL20, LIF and TYMS mRNA expression levels determined by quantitative real-time PCR (qRT-PCR) on GM-MØ either untreated (−) or treated for 48 h with IFN-γ, IL-4 or TNFα in the absence or in the presence of 50 nM of MTX. Mean and SEM of four independent donors are shown (*p<0.05). (D) CCL20 and LIF mRNA expression levels determined by qRT-PCR on GM-MØ and M-MØ, generated in RPMI 1640 medium without folic acid supplemented with 10% fetal calf serum during 7 days, in the absence or in the presence of 50 nM of MTX for 48 h. After 24 h of incubation with MTX, cells were treated with folic acid (FA, 50 nM). Mean and SEM of four independent donors are shown (*p<0.05).
Folic or folinic acids are prescribed with MTX therapy to reduce toxicity without reducing efficacy.22 Importantly, supplementation with folic acid (figure 5D) or folinic acid (not shown) did not modify the upregulation of CCL20 and LIF mRNA in MTX-exposed GM-MØ, thus indicating that MTX-induced macrophage gene expression is potentially relevant in the context of RA.
TS inhibition modulates p53-regulated genes in GM-MØ
The molecular pathways potentially responsible for the MTX-responsive transcriptome in macrophages were analysed by IPA. TP53 appeared as the top upstream regulator of the genes modulated by MTX during long-term and short-term schedules and with a predicted ‘activated’ state (figure 6A). Moreover, gene set enrichment analysis in 48 h MTX-treated GM-MØ showed a significant enrichment of the ‘p53 gene signature’ (figure 6B). These bioinformatics analysis, together with the detection of p53 expression in macrophages from RA and not from normal joints (see online supplementary figure S5), and the fact that TS inhibition by MTX increased p53 target genes like LIF, CCL20 or GDF15 (figure 4A–C),27 ,28 led us to explore the link between TS inhibition (MTX) and p53 activation. As shown in figure 6C, p53 blockade with pifithrin-α (PFT), a p53 inhibitor, dose-dependently diminished MTX-responsiveness (CCL20, LIF mRNA) in GM-MØ, thus indicating that the MTX-triggered gene expression changes in GM-MØ are mediated by both TS (figure 4C) and p53. To investigate the relationship between TS and p53, we determined the effects of PFT on the upregulation of CCL20 and LIF expression in GM-MØ caused by siRNA-mediated TS knockdown. The upregulation of both genes in response to TS silencing was significantly diminished in the presence of the p53 inhibitor (figure 6D), indicating that lowering TS levels results in enhanced p53 activation. In line with this result, activation of p53 with nutlin-328 led to increased expression of CCL20 and LIF genes in TS+ GM-MØ and TS− M-MØ (see online supplementary figure S6). Altogether, these results indicate that MTX inhibits TS, and that TS inhibition results in a decreased deoxythymidine pool, which leads to activation of p53 and increased expression of p53-responsive genes (LIF, CCL20) (figure 6E). As MTX-Pgl only affects the transcriptome of TS+ GM-MØ, these results demonstrate that the macrophage transcriptional response to MTX is polarisation-dependent and determined by the TS-p53 axis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Molecular basis of methotrexate (MTX) response in GM-MØ: TP53. (A) Top upstream regulators in long-term schedule, MTX-exposed monocytes and differentiated with GM-CSF (Mo+GM-CSF) and, short-term schedule, 48 h MTX-exposed GM-MØ, determined by Ingenuity Pathways Analysis (IPA). (B) Gene set enrichment analysis (GSEA) results for MTX-treated versus untreated GM-MØ for 48 h indicating the normalised enrichment score (NES) and the false discovery rate (FDR). (C) Gene expression determined by qRT-PCR on GM-MØ stimulated with MTX in the absence or the presence of pifitrin-α (PFT, 25–50 µM) during 48 h. (D) Gene expression determined by quantitative real-time PCR (qRT-PCR) on GM-MØ transfected with control siRNA and siRNA for TYMS and exposed to PFT (50 µM) for 48 h. Results are expressed as fold induction, which indicates the expression of each gene in siRNA TYMS-transfected relative to siRNA control cells, and untreated or treated with PFT. For (C and D), mean and SEM of six independent donors are shown (*p<0.05, **p<0.01, ***p<0.001). (E) Proposed model of MTX-induced response in GM-MØ. MTX enters macrophages through reduced folate carrier (RFC), becomes polyglutamated (MTX-Pgl) and inhibits the enzyme thymidylate synthase (TS). TS inhibition results in decreased pool of deoxythymidine leading to activation of p53 and p53 response genes (LIF, CCL20). The putative role of LIF in the induction of Treg and M-MØ polarisation is shown.
Discussion
In the present report we show that responsiveness to MTX of human myeloid cells is cell type-specific and correlates with the level of expression of TS. Specifically, proinflammatory TS+ GM-MØ accumulate higher intracellular levels of MTX-Glu2 and MTX-Glu3 and are susceptible to MTX, whereas anti-inflammatory TSlow/− M-MØ and monocytes are refractory to MTX. In fact, MTX responsiveness of human GM-MØ can be modified by altering TS expression or function by (1) thymidine, that bypasses the metabolic blockade; (2) PMX or raltitrexed, that selectively target TS; (3) siRNA-mediated knockdown of TS. Furthermore, we show that p53 activity contributes to the TS-dependent MTX-responsiveness of human proinflammatory macrophages, thus providing evidence for the involvement of the TS-p53 axis in the macrophage response to MTX and in the differential responsiveness to MTX of monocytes and macrophages. Gene ontology analysis predicted CEBPβ, STAT3 and Smad3 as potential upstream regulators for the set of MTX-regulated genes in GM-MØ. Whereas these results support an anti-inflammatory modulation of the GM-MØ transcriptional profile by MTX, members of the NFκB family are also predicted as potential regulators of the MTX-dependent transcriptome, a prediction that is in line with the ability of MTX to trigger proinflammatory cytokine production in U-937 cells.29 This apparent discrepancy might be explained by the cell-type specific effect of MTX, which might even explain the anti-inflammatory activity of MTX through the promotion of a tolerant state in susceptible macrophages: by analogy to the LPS-tolerance mechanism,30 GM-MØ exposed to (‘primed by’) MTX might become less responsive to a subsequent stimulation by proinflammatory stimuli (like TNFα).
The role of antiproliferative drugs on myelomonocytic cells has been previously explored.31–34 Trabectedin, an anticancer drug used as second-line treatment of soft tissue sarcoma,32 blocks cancer cell proliferation by interacting with DNA, but specifically induces cell death in monocytes and tumour-associated macrophages.32 Like trabectedin, MTX has selectivity for tumour-proliferating cells3 as well as TS-expressing proinflammatory macrophages with a GM-CSF-dependent transcriptional signature. Since GM-CSF critically contributes to the gene expression profile of macrophages in RA,26 our results suggest that the profound clinical benefit of MTX in RA4 might derive, at least partly, from an anti-inflammatory skewing ability exerted on the GM-CSF-conditioned pathology-driving macrophages within joints of patients with RA.
The involvement of the TS-p53 axis in the macrophage response to MTX is not completely unexpected. TS catalyses the reductive methylation of dUMP by 5,10-methylentetrahydrofolate to generate thymidylate and dihydrofolate (figure 3B), providing for the sole intracellular de novo source of thymidylate, an essential precursor for DNA biosynthesis and repair. As a result, TS remains a critical target enzyme in cancer chemotherapy. Inhibition of TS by drugs such as 5-fluorouracil results in decreased production of TMP and imbalances in cellular deoxynucleotide pools, which slow tumour growth by impairing DNA synthesis.35 In the case of RA, genetic polymorphisms affecting TYMS gene expression have been evaluated for their association with MTX clinical response, but contradictory results have been reported so far.36–38 Interestingly, however, TS represses the expression of tumour suppressor genes such as myc and p53.39 These previous evidences are compatible with the data in this report, as we demonstrate that MTX responsiveness of macrophages is partly mediated by p53, because p53 inhibitors limit the MTX-induced upregulation of genes like LIF and GDF15, well known p53 targets.27 Based on these results, and given that p53 somatic mutations have been found in RA synovium,40 ,41 we speculate that mutations in p53 might lead to MTX resistance in patients with RA, a phenomenon already established in tumours.42 Thus, determining the expression/activity of either p53 or MTX-response genes in MTX responders and non-responders patients with RA, might allow the identification of clinically useful predictors of response to MTX, a tool currently non-existing in spite of the fact that MTX is the first choice for RA management.4
In the present report, we also demonstrate that LIF and CCL20 are novel markers for MTX responsiveness in proinflammatory macrophages in vitro. LIF is constitutively expressed in tissues where macrophages are immunosuppressive and protolerogenic43–45 and, in fact, tumour-associated LIF and IL-6 drive monocytes into M2-polarised macrophages in an M-CSF-dependent manner.46 Accordingly, conditioned-medium from MTX-exposed GM-MØ augments M2-polarising markers in monocytes (not shown). MTX is a drug with an exceptional potency to treat inflammatory joint disease; however, the precise mechanism by which MTX exerts its anti-inflammatory effects remains unclear. Consistent with the immunoregulatory role of LIF, it can be hypothesised that MTX-induced LIF might promote M2 polarisation in synovial monocytes, thus restoring normal macrophage phenotype within the synovium.26
MTX is a main anchor drug, alone or in combination regimens, for RA. Although MTX exhibits a good benefit/toxicity ratio, hepatotoxicity, pneumonitis or neutropenia are MTX-adverse effects which justify MTX treatment withdrawal in patients with RA.5 ,47 ,48 Since the transcriptional signature of MTX-treated macrophages indicates a significant increase in the expression of genes involved in hepatic fibrosis, and MTX selectively targets GM-CSF polarised macrophages as those found in the lung,49 the hypothesis that MTX-mediated hepatotoxicity and/or pneumonitis is macrophage-dependent deserves further consideration.
Acknowledgments
The authors thank Miguel A Vega for technical help and Paloma Sánchez-Mateos and Angel L Corbí for helpful discussions.
References
Footnotes
Handling editor Tore K Kvien
CM and BSP contributed equally.
Contributors CM, BSP and LE-C: designed research, performed research and analysed data. AB and AD: analysed microarray data. SF-A: performed research and analysed data. JJ, MEM-C, EG-L and IG-A contributed vital reagents and materials and analysed data. AP-K conceived the study, designed research, analysed data and wrote the paper. All authors approved the final version.
Funding This work was supported by grant PI14/00075 from Instituto de Salud Carlos III/FEDER to AP-K, and grants S2010/BMD2350 from Comunidad de Madrid/FEDER (RAPHYME Program) (MEM-C and AP-K) and RIER RD12/0009/FEDER (MEM-C, IG-A and AP-K) (FEDER, Fondo Europeo de Desarrollo Regional: una manera de hacer Europa). AP-K is supported by FIBHGM and LEC is supported by CONACYT.
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.