Article Text

Abstract
Objective Ankylosing spondylitis (AS) is a chronic inflammatory disease of unknown origin in which interleukin (IL) 17 has been genetically and therapeutically recognised as a key player. Identification of the cellular sources and inducers of IL-17 is crucial in our understanding of the drivers of inflammation in AS. Recently, mucosal-associated invariant T (MAIT) cells have been implicated in autoimmune diseases. Their gut origin, effector phenotype and expression of multiple AS-associated genes, such as IL7R and IL23R, makes them potential contributors to the pathogenesis of AS.
Methods Mononuclear cells from patients with AS, healthy controls (HCs) and patients with rheumatoid arthritis were isolated from blood and synovial fluid (SF). Flow cytometry was used to identify MAIT cells. Phenotype was assessed by intracellular staining for cytokines and granzyme. Function was assessed by antigen-specific stimulation using Salmonella, or antigen non-specific activation via priming with IL-7 or IL-23.
Results MAIT cells were reduced in frequency in the blood of patients with AS compared with HCs, yet patients with AS had an elevated frequency IL-17A+ MAIT cells. There was an enrichment of MAIT cells in SF, which had an exaggerated IL-17 phenotype. IL-17 elevation in AS MAIT cells was dependent on priming with IL-7 but not IL-23 or antigen stimulation. The AS-associated IL7R single nucleotide polymorphism (SNP), rs11742270, had no effect on IL-7R expression or function in the experiments performed.
Conclusions This study reveals a potential role for MAIT cells in patients with AS and is the first linking IL-7 to the elevated IL-17 profile in patients through the AS-associated risk gene IL7R.
- Ankylosing Spondylitis
- T Cells
- Cytokines
- Gene Polymorphism
Statistics from Altmetric.com
Introduction
Ankylosing spondylitis (AS) is an inflammatory arthritis of the axial skeleton with a prevalence of up to 0.2% in Caucasian populations.1 The Th17-axis has been implicated in the pathogenesis of AS by elevations in type-17 cells including ILC3, γδ T cells and T helper 17 cells, and by increases in serum interleukin (IL)-23 and IL-17.2 The majority of patients with AS have subclinical gut inflammation,3 with genetic studies confirming the link between inflammatory bowel disease (IBD) and AS.4 Yet, a cellular basis of the gut-joint axis in AS is unresolved.
Mucosal-associated invariant T (MAIT) cells are innate-like lymphocytes found preferentially in the gut lamina propria.5 They have been implicated in a variety of autoimmune diseases, such as multiple sclerosis, IBD, systemic lupus erythematosus and rheumatoid arthritis (RA).6–8 In a murine model of RA, MAIT cells were found to exacerbate arthritis,9 suggesting they may be arthritogenic.
The T cell receptor (TCR) repertoire of human MAIT cells is comprised chiefly of Vα7.2-Jα33, which restricts their specificity to the major histocompatibility compex (MHC) class 1-like molecule, MR1. MR1 is unique in that it presents bacterially derived vitamin B metabolites.10 Indeed, bacteria are essential for the development of MAIT cells as germ-free mice lack MAIT cells.5 Functionally, MAIT cells produce multiple inflammatory effectors such as IL-17, interferon gamma (IFNγ), tumour necrosis factor alpha (TNFα) and granzyme. These cells are predominantly CD8α+ and express Th17-associated markers such as IL-23R, CCR6, CD161 and RORγt.11 MAIT cells also express high levels of IL-7R.12
IL-7 is a pleiotropic cytokine. In primary lymphoid organs, IL-7 is a modulator of B cell and T cell development, while in the periphery it plays a crucial role in regulating homoeostatic proliferation of T cells.13 ,14 IL-7 has recently been shown to prime γδ T, Th17 and MAIT cells to produce proinflammatory cytokines, including IL-17.12 ,15 ,16 The IL-7R forms a heterodimer with the common γ chain (γc) to mediate signalling via JAK-STAT pathways.17 IL-7R can also associate with thymic stromal lymphopoietin (TSLP) receptor (TSLPR) to mediate TSLP signalling, a key component of Th2 polarisation. The IL7R gene was recently identified in immunochip studies to be associated with AS.18 Whether IL-7R expression or function is altered in patients with AS is unknown.
Given the inflammatory nature of MAIT cells, their role in gut inflammation and the involvement of the gut in AS, we sought to characterise MAIT cells in AS. We further aimed to characterise the function of MAIT cells in patients with AS in response to antigen-specific and non-specific (IL-7) stimulation.
Methods
Patient cohorts
All patients with AS had radiographic sacroiliitis and satisfied the modified New York (NY) criteria for AS. Healthy controls (HCs) were age-matched and sex-matched to patients with AS (see online supplementary table S1). Volunteer HCs were recruited concurrently with patients. Patients with RA served as an inflammatory disease control group. A subset of male patients and HCs were examined in MAIT cell functional studies (see online supplementary table S2). All patients and controls completed an research ethics board (REB)-approved consent form to participate.
Supplemental material
Flow cytometry
Peripheral blood mononuclear cells (PBMCs) were isolated by Ficoll-Paque plus (General Electric (GE)) from venous blood obtained during clinic hours. Synovial fluid (SF) from joint aspiration was diluted 1:5 in phosphate buffered saline (PBS), centrifuged, and SF mononuclear cells isolated by Ficoll-Paque.
Where applicable, cells were stimulated with Phorbol 12-myristate 13-acetate (PMA)/ionomycin (Sigma) and brefeldin A (Becton Dickinson (BD)) in complete media (Roswell Park Memorial Institute (RPMI) with 10% heat inactivated fetal calf serum (HI-FCS) and penicillin/streptomycin), for 4 h prior to staining with fixable live/dead near infra-red (NIR) (Invitrogen). Cells were blocked with FcX (BioLegend). The following fluorochrome-conjugated antibodies were used for surface staining: CD8α-pacific blue (PB), CD3ε-BV605, CD4-fluorescein isothiocyanate (FITC), CD161-PE, CCR6-PerCP.Cy5.5, CD69-PE.Cy7 and Vα7.2-Allophycocyanin (APC) (BioLegend). For the intracellular panel CD161-PerCP.Cy5.5 and Vα7.2-APC (BioLegend) were used for surface staining. For intracellular staining, cells were further fixed and permeabilised (BD) prior to staining with IFNγ-PB, TNFα-BV605, granzyme B-FITC, IL4-PE and IL-17A-PE.Cy7 (BioLegend). Data were acquired on LSRII or CantoII (BD) and analysed in FlowJo (TreeStar).
For in vitro culture assays, the above protocol was used with the following antibodies: CD69-PB, CD161-PE, Vα7.2-APC, TNFα-BV605, granzyme B-FITC and IL-17A-PE.Cy7 (BioLegend). For assessment of MAIT cell proliferation and TSLPR expression with cytokine priming, the additional reagents used were eFlour450 Cell Proliferation Dye (eBioscience) and TSLPR-PE (BioLegend).
For IL-7R characterisation, frozen PBMCs were rested overnight in media. The above protocol was used for surface staining with the following antibodies: IL-7R-BV421, CD3ε-BV605, CD4-FITC, CD161-PE, CD14-PerCP.Cy5.5, CD8α-PE.Cy7 and Vα7.2-APC (BioLegend).
MR1-specific antigen stimulation assay
For the antigen-specific stimulation assay, Salmonella enterica serovar typhimurum (a gift from Dr D Philpott), was grown to confluence in Luria broth (Sigma) by shaking overnight at 37°C. The number of live bacteria was estimated by OD600 (1 OD=108 bacteria). Salmonella was fixed with 2% paraformaldehyde (PFA) in PBS for 20 min on ice. Aliquots of fixed Salmonella were subsequently frozen in PBS at −80°C. THP-1 cells (a gift from Dr J Jongstra-Bilen) were cultured in complete media.
THP-1 cells were cultured overnight at 5×104 cells in a 96-well u-bottom plate with/without Salmonella. The dose of Salmonella was titrated in pilot experiments (see online supplementary figure S1) and 50 multiplicity of infection (MOI) was selected for experiments based on previous reports.19 Fresh PBMCs were rested overnight in complete media prior to use. Free bacteria were washed from the THP-1 cells and 0.75×106 PBMCs added to each well. Low endotoxin anti-MR1 antibodies, or isotype control (BioLegend) were added to respective wells at 10 µg/mL. Cells were cocultured for 24 h, with brefeldin A added for final 4 h at 10 µg/mL. PBMCs were assayed for activation by flow cytometry as discussed above.
Cytokine response assay
Fresh PBMCs were plated at 0.75×106 cells/well in complete media. IL-7 or IL-23 (BioLegend) were added to respective wells for 48 h. In pilot experiments, IL-7 and IL-23 were titrated (see online supplementary figure S2) based on previous reports,12 and 20 ng/mL selected for experiments. Brefeldin A 2 µg/mL was added to the cultures overnight with or without T cell activation beads (Miltenyi) as recommended by the manufacturer. PBMCs were assayed for activation by flow cytometry.
qPCR
Frozen PBMCs were rested overnight in complete media. RNA was extracted using RNeasy columns (Qiagen) and purity assessed by Nanovue (GE). All samples that had 260/280>1.8. RNA were subjected to DNAse treatment (Invitrogen), prior to reverse transcription with Superscript III (Invitrogen).
IL7R primers (F:5′-GCACTCACTGACCTGTGCTTTT-3′, R:5′- GCCCCACATATTTCAAATTCCA-3′) were designed using Primer Express 3 (Applied Biosystems). Specificity to IL7R was determined using PrimerBlast (National Center for Biotechnology Information (NCBI)), and was confirmed with a melt curve showing a single peak. Housekeepers were designed, validated and tested for stable expression as previously reported.20
Gene expression was analysed in triplicate using N',N'-dimethyl-N-[4-[(E)-(3-methyl-1,3-benzothiazol-2-ylidene)methyl]-1-phenylquinolin-1-ium-2-yl]-N-propylpropane-1,3-diamine (SYBR) green on the 7900HT system (Applied Biosystems). As primers displayed efficiencies >95%, the 2−ΔΔCt method was used for analysis.
Single nucleotide polymorphism (SNP) assay
Primers for rs11742270 (C_31636680_10) and Genotyping MasterMix were ordered from Applied Biosystems. Analysis was performed on 7900HT following the manufacturer's instructions.
Statistics
All data were graphed and analysed in GraphPad Prism. Data were subjected to D'Agostino and Pearson normality test, before selecting appropriate test as indicated in the respective figure legends. For all graphs, non-significant results (p>0.05) were indicated or left blank; *p=0.05–0.01; **p=0.01–0.001 and ***p<0.001.
Results
MAIT cells are enriched in the inflamed joint where they display an activated phenotype
Initially we characterised MAIT cells in AS by assessing their frequency and resting phenotype in PBMC. As no MR1-tetramer is commercially available, we followed the convention of defining MAIT cells as CD3+Vα7.2+CD161hi (figure 1A). The majority of these cells were CD8α+ and all were CCR6+ as demonstrated by comparison to CD3+Vα7.2+CD161− cells, which are considered conventional T cells.21 MAIT cells, as a frequency of total T cells, were reduced in AS PBMC by almost 50% compared with HCs (figure 1B; median 2.1% vs 1.2%). Despite this, levels of the activation marker CD69, the homing marker CCR6 and the CD4/CD8 subtype were equivalent to that seen in HC (figure 1B). MAIT cell frequency and CD8α positivity was reduced in patients with RA as previously reported.6 Blood MAIT cell frequency did not correlate with c reactive protein (CRP), erythrocyte sedimentation rate (ESR) or Bath Ankylosing Spondylitis Disease Activity Index (BASDAI), nor did it stratify with sex or treatment type.
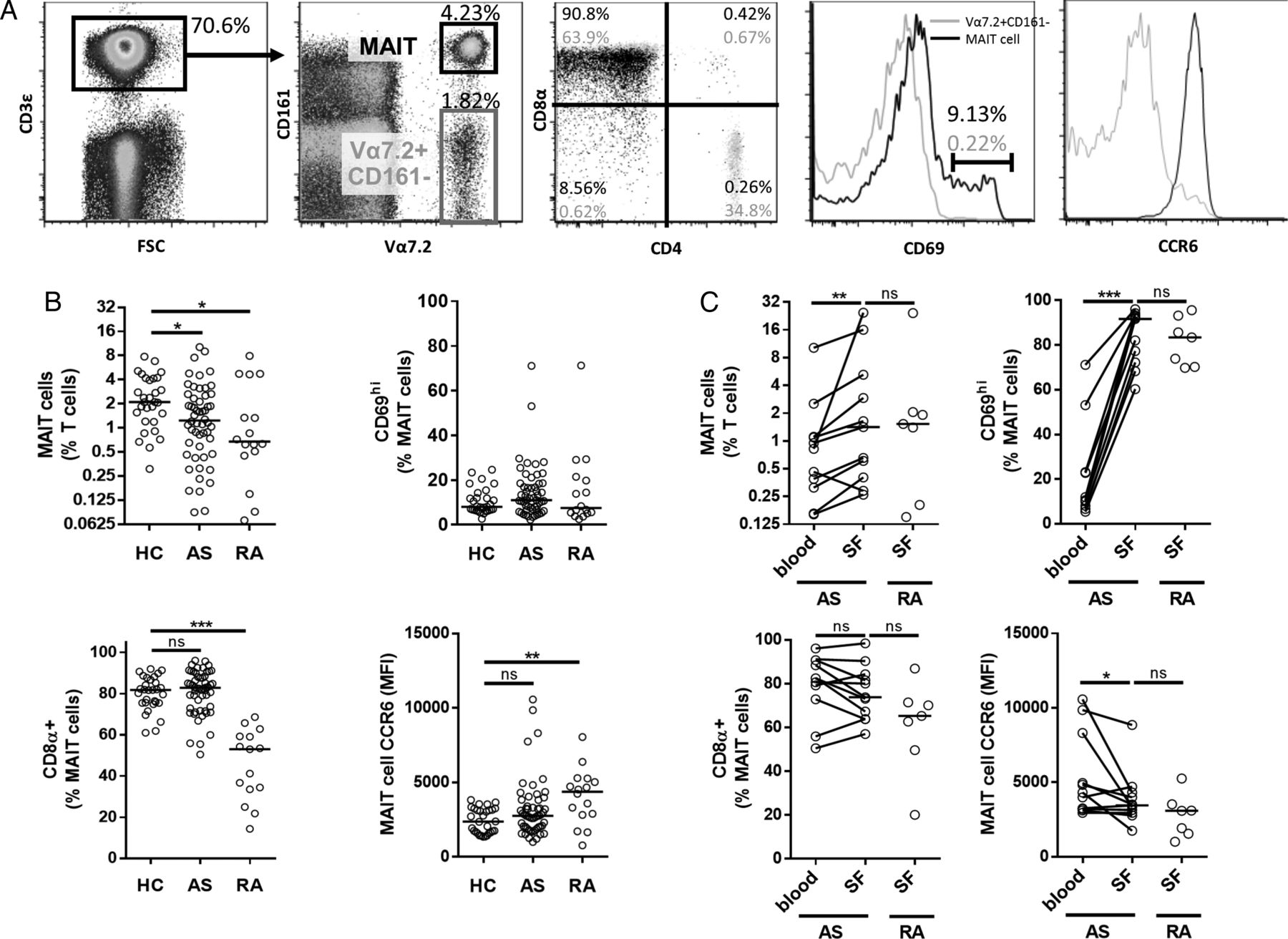
Identification and composition of blood and synovial fluid (SF) mucosal-associated invariant T (MAIT) cells by flow cytometry. (A) Representative plots demonstrating gating strategy on fresh, unstimulated peripheral blood mononuclear cells (PBMCs)/SF mononuclear cells (SFMCs). Live, singlet lymphocytes (not shown) subjected to indicated gating strategy to identify MAIT cells (CD3+Vα7.2+CD161hi) in black. CD3+Vα7.2+CD161− cells (grey) used for comparison to demonstrate CD4, CD8α, CCR6 and CD69 in representative plots. Percentages given in plots represent MAIT cells (black) and CD3+Vα7.2+CD161− cells (grey). (B) Frequency of MAIT cells and their expression of CD69, CD8α and CCR6 in patients with ankylosing spondylitis (AS), patients with rheumatoid arthritis (RA) and healthy controls (HCs). (C) MAIT cell frequency and composition in coincident blood/SF samples from patients with AS and RA SF disease controls. For all graphs, horizontal lines represent the median. Kruskal–Wallis test with Dunn's post-test analysis used to compare patients with HCs in (B) and Wilcoxon matched pairs test and Mann–Whitney test used where appropriate in (C).
We next sought to characterise MAIT cells in inflamed joints. As axial joint synovial samples are not accessible, we used SF from peripheral joints of patients with AS. The SF obtained was highly cellular (17.6±10×106 cells/mL) and ∼90% polymorphonuclear based on Wright–Giemsa staining (not shown). MAIT cells were enriched in SF as compared with the matched PBMCs; however a similar frequency of SF MAIT cells was seen in RA SF (figure 1C). It has previously been reported that MAIT cells are enriched in RA SF.6 MAIT cells in SF remained CCR6-positive, however CCR6 median fluorescent intensity (MFI) was diminished compared with paired MAIT cells in blood. This reduced CCR6 coincided with an activated phenotype as judged by elevated CD69 expression.
Frequency of IL-17+ MAIT cells is elevated in male patients with AS
As MAIT cells are capable of producing TNFα and IL-17, cytokines central to the pathogenesis of AS, we examined the cytokine profile of these cells in patients with AS. Since all Vα7.2+CD161hi cells were CD3+ (not shown), we did not use CD3 for the identification of MAIT cells in our intracellular panel. In unstimulated samples treated with brefeldin, only granzyme B was detectable at equivalent levels to PMA/ionomycin stimulated cells (not shown). As cytokines were otherwise undetectable in the absence of stimulation, we used PMA/ionomycin for all experiments. Figure 2A demonstrates the expression pattern of granzyme B, IL-4, IL-17, IFNγ and TNFα in MAIT and Vα7.2+CD161− cells. Expression of IL-17 was mutually exclusive with respect to IL-4 and granzyme B (figure 2B). On the other hand, IL-17 was coexpressed with IFNγ and TNFα, as were IFNγ and TNFα.

Intracellular cytokine and granzyme B composition of blood and synovial fluid (SF) mucosal-associated invariant T (MAIT) cells. (A) Representative plots demonstrating gating strategy used on stimulated peripheral blood mononuclear cells (PBMCs)/SF mononuclear cells (SFMCs). Live, singlet lymphocytes (not shown) subjected to indicated gating strategy to identify MAIT cells (Vα7.2+CD161hi). Histograms demonstrate gating strategy for respective factors using Vα7.2+CD161− cells (grey) as a comparison for MAIT cells (black). (B) Biexponential plots display co-staining of indicated factors in MAIT cells. (C) Frequency of granzyme B and cytokine-positive MAIT cells in the peripheral blood of patients with ankylosing spondylitis (AS), patients with rheumatoid arthritis (RA) and healthy controls (HCs). (D) IL-17A+ and IFNγ+ MAIT cell stratified for sex in the peripheral blood of patients with AS and HCs. (E) Frequency of granzyme B and cytokine-positive MAIT cells in coincident blood/SF samples and RA SF controls. For all graphs, horizontal line represents the median. Kruskal–Wallis test with Dunn's post-test analysis used to compare patients with HCs in (C), Mann–Whitney test used in (D) and Wilcoxon matched pairs test and Mann–Whitney test used where appropriate in (E).
In blood of patients with AS, there was a significantly higher frequency of IL-17+ MAIT cells, while their IFNγ production was significantly lower compared with HC (figure 2C). The elevated frequency of IL-17A+ MAIT cells was not seen in the peripheral blood of RA controls. Granzyme B, TNFα and IL-4 were unchanged in patients with AS versus HC. MAIT cell cytokine/granzyme levels did not correlate with BASDAI, CRP or ESR. We further observed that while the downregulation of IFNγ in MAIT cells was independent of sex, the upregulation of IL-17 was restricted to male patients with AS (figure 2D), as we have previously reported in Th17 cells.20 Male and female patients in this cohort did not significantly differ in any clinical variables presented in online supplementary table S1.
Synovial MAIT cells in patients with AS display unique inflammatory phenotype
Using our paired blood/SF samples, we observed significant alterations in the phenotype of SF MAIT cells in patients with AS. While IL-4 levels remained unchanged in the SF (not shown), IFNγ↑-positive and TNFα-positive MAIT cells were significantly decreased in AS SF (figure 2E), while the granzyme and IL-17 frequency of AS SF MAIT cells was increased. Surprisingly, MAIT cells in RA SF showed a distinct phenotype: the frequencies of IFNγ-positive and TNFα-positive cells were comparable to that seen in AS SF, however IL-17 and granzyme B producing MAIT cells were significantly lower compared with AS SF.
AS patient MAIT cells do not have altered response to Salmonella antigen
MAIT cells can be activated directly through TCR recognition of MR1-presented microbial metabolites.10 ,12 As MR1-presented antigens are not commercially available, we tested MAIT cell antigen response using an in vitro assay in which THP-1 cells present PFA-killed Salmonella to PBMC.10 Because the PMA/ionomycin-induced MAIT cell IL-17 upregulation in patients with AS was specific to men, we opted to examine a defined cohort of male patients. We further focused on young subjects (<35 years) to reduce the variation of MAIT cell frequency/phenotype, which is age-dependent.22
Our assay demonstrated upregulation of MAIT cell CD69, TNFα, granzyme B and IL-17A in the presence of Salmonella (figure 3). This activation was MAIT cell-specific as illustrated by Vα7.2+CD161− control cells showing no such activation with Salmonella infection. Further, anti-MR1 treatment effectively abrogated MAIT cell activation, demonstrating this activity was TCR-dependent. Strikingly, AS MAIT cells did not have significantly elevated inflammatory profiles when compared with HC. This provides evidence that the aberrant MAIT cell phenotype observed in the blood and SF of patients with AS is not due to altered response to MR1-presented antigen.
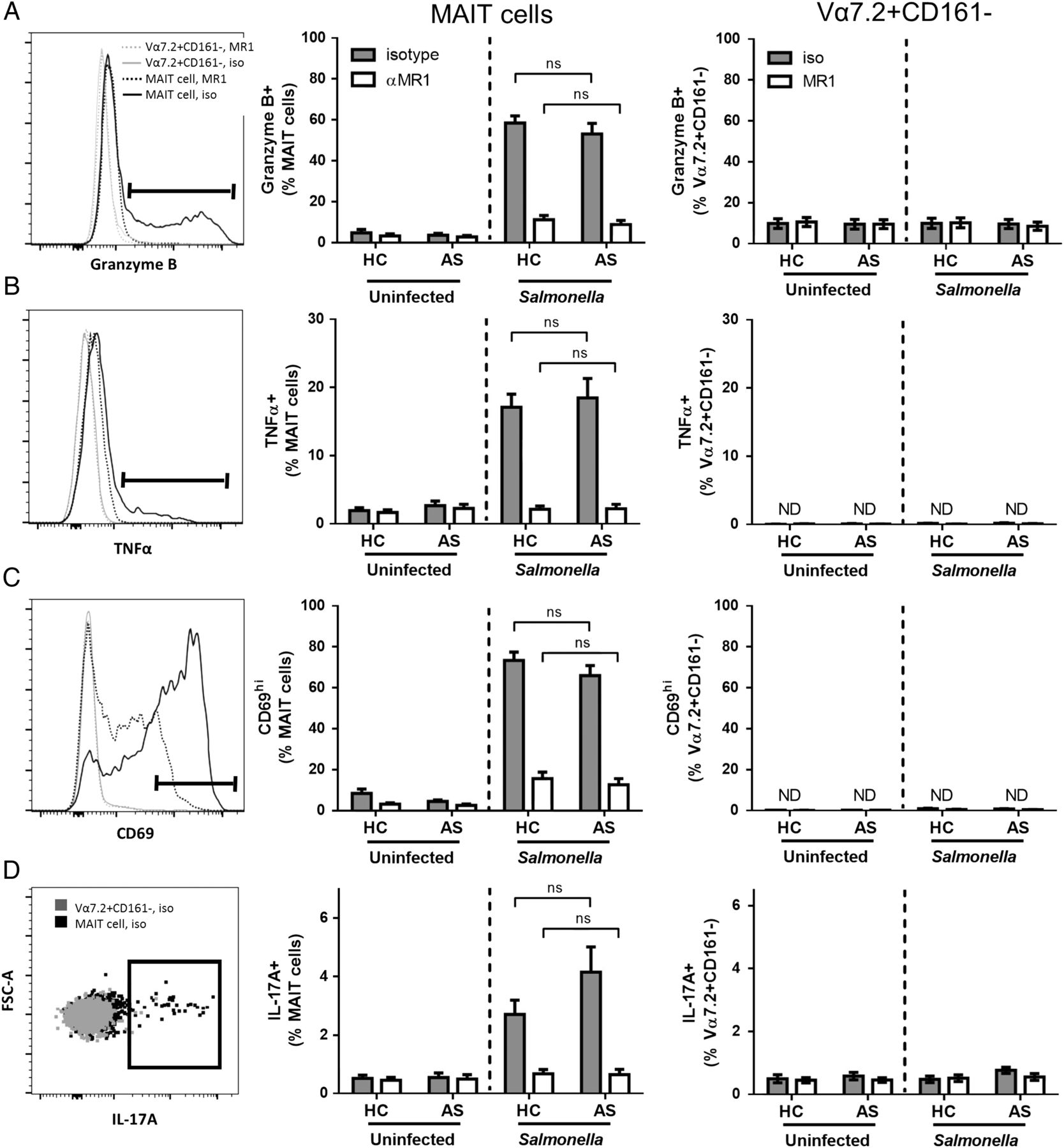
Mucosal-associated invariant T (MAIT) cell antigen-specific responses in patients with ankylosing spondylitis (AS) and healthy controls (HCs). THP-1 cells were infected overnight with PFA-killed Salmonella prior to 24 h co-culture with peripheral blood mononuclear cells (PBMCs). Brefeldin A was added for final 4 h of co-culture. Live, single lymphocytes (FSCloSSClo) selected to identify MAIT cells (Vα7.2+CD161hi) and Vα7.2+CD161− control cells for (A) granzyme B, (B) TNFα, (C) CD69 and (D) IL-17A. Representative histograms show expression of selected markers in Salmonella exposed co-cultures with MR1 blocking antibody (hashed lines) or isotype control (full lines) in MAIT cells (black) or Vα7.2+CD161− cells (grey). Graphs display pooled results from three independent experiments with or without Salmonella and with anti-MR1 or isotype control. ND=not detected. Data displayed as mean±SEM (AS n=10, HC n=11). Salmonella stimulated co-cultures analysed by repeated measures two-way analysis of variance (ANOVA) with Bonferroni post-test analysis to compare AS and HC.
MAIT cells express IL-7R and are primed following IL-7 stimulation
As we saw no differential activation of AS MAIT cells through TCR stimulation, we explored MAIT cell activation through cytokine stimulation. IL-7 was of particular interest to us as polymorphisms in its receptor, IL-7R, are associated with AS18 and IL-7 can prime MAIT cells to produce IL-17.12 IL-23 was selected because of its established role in the induction of IL-17, and its role in the pathogenesis of AS.2
We performed flow cytometry on PBMC for IL-7R to confirm expression on MAIT cells and to compare expression across cell types, (figure 4A). Of note, IL-7R was undetectable on CD14−CD3− cells, which includes natural killer (NK) and B cells. Further, monocytes expressed marginal levels of IL-7R relative to isotype control staining. Vα7.2−CD4+, Vα7.2−CD8+ and Vα7.2+CD161− T cells expressed negative to intermediate levels of IL-7R. Strikingly, IL-7R expression was highest on MAIT cells and CD161hiVα7.2− T cells, which include Th17 cells (see online supplementary figure S3A). In addition, we examined PBMC for TSLPR expression, which can compete with the γc for IL-7R, thus favouring TSLP over IL-7 as a ligand.17 TSLPR was expressed on <0.1% of lymphocytes and <0.3% of monocytes (not shown) and was not upregulated after IL-7R stimulation (see online supplementary figure S4), so we did not focus on it.

Characterisation of IL-7R expression in peripheral blood mononuclear cells (PBMCs) by flow cytometry. (A) Live, singlet lymphocytes/monocytes (not shown) subjected to illustrated gating strategy on representative subject. Shaded histograms demonstrate IL-7R expression and empty histograms isotype control staining for indicated cells. (B) IL-7R MFI on mucosal-associated invariant T (MAIT) cells and (C) Vα7.2+CD161− cells determined by flow cytometry. (D) IL7R expression normalised to GAPDH in PBMCs determined by qPCR. Data in (B–D) displayed as a scatter plot with median and analysed by Mann–Whitney test.
Using this gating strategy, we compared IL-7R levels across cell subsets of interest between patients with AS and controls. We found that there was significantly higher expression of IL-7R on MAIT cells from patients with AS compared with controls (figure 4B). No such difference was seen in cell subsets expressing intermediate levels of IL-7R, such as Vα7.2+CD161− cells (figure 4C). This dichotomy demonstrates a cell-specific alteration in IL-7R expression in patients with AS, in which the majority of cells have IL-7R levels equivalent to that seen in HCs. This is reflected by a lack of differential expression of the IL7R gene between patients with AS and HCs in whole PBMCs (figure 4D).
To test MAIT cell responsiveness to these cytokines, we treated PBMCs for 60 h with IL-7 or IL-23. Treatment with IL-7, but not IL-23, was able to increase TCR (Vα7.2) and CD69 on MAIT cells (figure 5). In contrast, the Vα7.2+CD161− controls cells did not demonstrate a response to IL-7. When comparing TCR and CD69 expression between AS and HC, we observed no difference at baseline or with cytokine stimulation. This suggests that IL-7R signalling leading to TCR/CD69 upregulation is not altered between patients with AS and controls. Further, IL-7 stimulation does not induce MAIT cell proliferation (see online supplementary figure S4), favouring cell activation over expansion as an explanation for enhanced frequency of activated cells.

A role for IL-7 in priming mucosal-associated invariant T (MAIT) cells. Peripheral blood mononuclear cell (PBMC) treated for 60 h with 20 ng/mL IL-7 or IL-23 prior to analysis by flow cytometry. Live, single lymphocytes (FSCloSSClo) selected to examine MAIT cells (Vα7.2+CD161hi) and Vα7.2+CD161− cells for (A) Vα7.2 and (B) CD69. Representative histograms show MAIT cells either untreated (hashed), IL-23 treated (grey) or IL-7 treated (black). Graphs display pooled results from five independent experiments with data displayed as mean±SEM (ankylosing spondylitis (AS) n=21, healthy control (HC) n=13). TCR, T cell receptor.
AS patient MAIT cells have elevated IL-17 production upon priming with IL-7
In order to examine MAIT cell priming by IL-7, we followed published protocols treating PBMCs with cytokine for 48 h prior to overnight stimulation with anti-CD3/CD2/CD28.12 The priming of MAIT cells by IL-7 was dramatic as illustrated by the large increases in TNFα, granzyme B and IL-17 in all subjects (figure 6). Patients with AS and HCs showed no significant difference in TNFα and granzyme production following IL-7 priming, in line with our observations of CD69 and TCR upregulation. On the contrary, MAIT cell IL-17 frequency was found to be higher in patients with AS than HCs with IL-7 priming, and not with IL-23 or in the absence of cytokine treatment (figure 6C). As previously reported,23 IL-23 alone has a minimal effect on MAIT cell IL-17 production despite MAIT cells expressing high levels of IL-23R.12 Further, the effect of IL-7 was dramatically reduced in Vα7.2+CD161− which are IL-7R-negative/intermediate (figure 4A). Thus we observed a distinct effect of IL-7 priming on IL-17 production in MAIT cells in patients with AS.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cytokine and granzyme production in IL-7 primed, T cell receptor (TCR) stimulated peripheral blood mononuclear cells (PBMCs) assessed by flow cytometry. PBMCs treated for 60 h with 20 ng/mL IL-7 or IL-23. Brefeldin A added with or without anti-CD3/CD2/CD28 for final 12 h. Representative histograms show cytokine/granzyme frequency of mucosal-associated invariant T (MAIT) cells either untreated (hashed), IL-23 treated (grey) or IL-7 treated (black) for (A) granzyme B, (B) TNFα and (C) IL-17A. Graphs display pooled results from five independent experiments with data displayed as mean±SEM (ankylosing spondylitis (AS) n=21, healthy control (HC) n=13). Data from TCR-stimulated samples analysed by repeated measures two-way analysis of variance (ANOVA) with Bonferroni post-test analysis to compare AS and HC.
To further support the IL-17 inducing role of IL-7 in IL-7Rhi cells, we examined the Vα7.2-CD161hi cells, which contain many Th17 cells.24 In a TCR-stimulated, unprimed state, this population contained a higher frequency of IL-17+ cells in patients with AS compared with HCs, but not granzyme B+, TNFα+ or CD69hi (see online supplementary figure S3B–E). This supports observations of an elevated Th17 frequency in patients with AS.2 IL-7 priming induced significantly higher frequencies of IL-17+Vα7.2-CD161hi cells in patients with AS versus controls (see online supplementary figure S3E), suggesting that the effect of IL-7 on augmentation of IL-17 in patients with AS extends beyond MAIT cells.
No significant impact of the AS-associated IL7R SNP (rs11742270) on IL-7R expression or IL-7 priming
Recent immunochip studies have identified rs11742270 as a risk factor for AS. This SNP is ∼2 kb downstream from the 3′ untranslated region (UTR) of IL7R. We performed SNP analysis to determine if the data from our experiments stratified with rs11742270. None of our patients or controls was homozygous for the protective allele (A), however we did have a reasonable number of patients heterozygous or homozygous for the risk allele (G). We saw no significant differences in IL-7R expression at the mRNA level in whole PBMCs, or at the protein level on MAIT cells between subjects with AG or GG genotypes (see online supplementary figure S5A,B). Further there was no significant effect on IL-17 production in IL-7 primed MAIT cells by the rs11742270 genotype (see online supplementary figure S5C). We further examined IL-17 production in IL-7 primed MAIT cells to see if it was related to the expression level of IL-7R; however we found direct correlation in patients with AS or controls (see online supplementary figure S5D). These results suggest that the rs11742270 SNP is not a causative genetic variant, but is likely linked to one as is the case for most genome-wide association study (GWAS)-identified SNP.4
Discussion
Although MAIT cells have only recently been identified, their importance in inflammatory disease is becoming evident. Our observations of reduced MAIT cell frequency in the peripheral blood of patients with AS mirrors that seen in IBD.8 In AS the reduced MAIT cell frequency in blood likely reflects recruitment to sites of inflammation such as the gut, or as we report, the inflamed joint. It is important to point out that MAIT cell enrichment in the inflamed joint is not specific to AS as RA SF has equivalent levels. Nevertheless, the observation of gut-associated MAIT cells in the arthritic joint suggests a sharing of tissue homing molecules. MAIT cells express markers implicated in homing to the gut and joint, such as CCR2, CCR6, CCR9 and α4β7.11 The factors that mediate such gut-joint trafficking need to be clearly defined and may be important therapeutic targets as has been shown with α4β7 in Crohn's disease.25
Recent genomic and immunophenotyping studies have implicated Th17 cells in the pathogenesis of AS. Despite MAIT cells being distinct in their development and MHC restriction compared with Th17 cells, they show a similar increase in IL-17 frequency in AS. In addition to this, there was sexual dimorphism in IL-17 frequency in AS MAIT cells, a phenomenon we have recently observed in Th17 cells of patients with AS.20 This suggests a global alteration of the type-17 axis in patients with AS rather than a specific alteration in Th17 cells, supported by observations of elevated frequencies of IL-17+ γδ T cells and ILC3.26 ,27
Our findings provide a mechanistic insight into the IL-17 phenotype of AS MAIT cells. Although we confirm previous reports of an MR1-dependant MAIT cell activation, we observed no difference between such responses in patients with AS and HCs. We caution that the exact nature of the Salmonella antigen in our assay is not defined. It is possible that different vitamin B metabolites may alter MAIT cell polarisation as is seen in natural killer T cell (NKT) cells.28 In contrast we found a differential response to IL-7 priming between patients with AS and controls, specifically in IL-17. Pertinent to this, IL-7 is elevated in gut and joint tissues of patients with AS.27 ,29
The linking of IL-7 to IL-17 production in patients with AS is of particular importance given the association of IL7R in immunochip studies.18 The associated SNP rs11742270 is located downstream of IL7R in a gene encoding-free and promotor-free region. It is thus unlikely that rs11742270 is itself a causative SNP. Although rs11742270 is flanked by two promotor regions, which can bind STAT3 and Runx3 (UCSC Genome Browser), the causative variant is unlikely in a transcription factor binding region as we observed no alteration in the IL-7R gene or protein expression with rs11742270. This hypothesis must be tested in a cohort larger than ours to insure adequate statistical powering. Allelic IL7R variants in leukaemia, multiple sclerosis (MS), RA and type 1 diabetes (T1D) occur in or near exon 6,30 ,31 which may be where the IL-17 promoting allelic variant lies in patients with AS. These exon 6 allelic variants both enhance IL-7R signalling and promote alternative splicing. Sequencing and in vitro functional studies will be required to address whether this occurs in AS-associated IL7R variants.
There are a number of caveats to this study: First, MAIT cells occur at too low a frequency for detailed in vitro studies on sorted/purified cells. Thus we can't exclude bystander activation in our in vitro assays. Second, AS primarily affects joints of the axial skeleton, and thus our observations in peripheral joints must acknowledge that it is not known whether the cell enrichment in sites of peripheral arthritis mirrors that seen in sites of axial inflammation. Third, our RA controls were not age-matched and sex-matched or treatment-matched to patients with AS. We recognise that this is not an ideal comparison but, given the demographics of the respective diseases, it is insurmountable. Thus, it is unclear whether differences between AS and RA is due to disease status, gender, age or treatment. Fourth, owing to the rarity of subjects homozygous for the protective IL7R SNP (AA), we were not able to examine the effect of such a genotype in our experiments. It is possible that the risk allele (G) is dominant over the protective allele (A). Detailed in vitro studies would be required to address such an issue.
In conclusion, these results suggests that MAIT cells contribute to the elevated type-17 axis in AS and may play a role in the pathogenesis of AS. The fact that patients with AS often have gut inflammation and that MAIT cells are critically dependent on the gut for their development, strengthens the concept of a gut-joint axis of inflammation in AS, and provides a cellular target for intervention. We further show a crucial role for IL-7 in promoting IL-17 production. This observation provides a rationale for targeting IL-7, or IL-7R, in the treatment of AS.
Acknowledgments
The authors thank Dr Juandy Jo and Dr Antonio Bertoletti from the Singapore Institute for Clinical Sciences for their advice in optimising the MAIT cell cytokine priming assay. The authors thank the following clinical research staff at the Toronto Western Hospital Spondylitis clinic for assistance in gathering clinical information and patient samples: Sister Esther Paeste, Kevin Seetaram, Renise Ayearst, Ammepa Anton and Julie Gagnier. The authors also thank Dr Edward Keystone (Mt Sinai Hospital, Toronto), for providing them with RA SF.
References
Footnotes
Handling editor Tore K Kvien
Contributors EG and RDI designed experiments. EG, ZQ, IA, SK, NA and YY performed experiments. EG, ZQ, DL, ZZ, VR, YB and RDI analysed and interpreted data. DL facilitated patient recruitment and procurement of patient data. EG and RDI wrote the manuscript. All authors agreed to publish the data and reviewed the manuscript.
Funding Canadian Institutes of Health Research (CIHR), and the Arthritis Research Center.
Competing interests None declared.
Ethics approval University Health Network.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement EG and RDI have full access to the data and take responsibility for the integrity of analysis. Data available upon request.