Article Text

Abstract
Objective: To investigate the presence and regulation of lymphatic vessels in inflamed joints of mice with experimental arthritis as well as patients with rheumatoid arthritis (RA) and spondyloarthritis (SpA).
Methods: Lymphatic vessels and blood vessels were assessed in synovial tissue of human tumour necrosis factor transgenic (TNFtg) mice and synovial biopsies from patients with RA and SpA by immunohistochemistry for podoplanin and CD31, respectively. Assessments were performed before and after TNF blockade in all biopsies.
Results: Lymphatic vessels were abundantly present in the synovial tissue of hTNFtg mice as well as patients with RA and SpA. The number of lymphatic vessels was positively related to the severity of synovial inflammation. Treatment with infliximab led to an increase in the formation of lymphatic vessels in murine and human inflammatory tissue.
Conclusions: This study shows that TNF blockade promotes the proliferation of lymphatic vessels in the inflamed synovium of RA and SpA. This finding leads to the assumption that promotion of lymphangiogenesis may play an important part in efflux of cells and fluid out of the inflamed tissue.
Statistics from Altmetric.com
Chronic destructive arthritides, such as rheumatoid arthritis (RA), psoriatic arthritis and spondyloarthritis (SpA), are common rheumatic diseases. The morphological substrate of the swollen joint in these diseases is an inflammatory synovial infiltrate, which causes the clinical picture of swelling, pain and dysfunction, and, if untreated, also results in irreversible structural damage.1 2 Histologically, synovial inflammation is based on activated resident cells (fibroblasts) and infiltrating immune cells.3–5
The pathophysiological steps leading to an influx of immune cells into the joint are considered as important therapeutic targets and require the spreading of blood vessels into the newly formed inflammatory synovial tissue. Recent reports have revealed some of these mechanisms required for the formation and activation of blood vessels in the inflamed synovium, thereby attracting inflammatory cells to the joint.6 In animal models of chronic arthritis and in human RA, the formation of a thick synovial lining layer and the accumulation of inflammatory cells in the sublining goes along with the appearance of intense neoangiogenesis.7 Although most studies have focused on cell influx into the joint, it is likely that immune cells do not just remain in the joint but that there is rather an extensive trafficking and recirculation of immune cells into draining lymph nodes. Furthermore, in the case of resolution of synovial inflammation upon efficient treatment, immune cells may leave the joint,8 which is likely to be accomplished by the lymphatic vascular system.9
Indeed, lymphatic vessels can be detected in RA joints and synovial cells produce lymphangiogenic factors upon stimulation with proinflammatory cytokines. However, little is known about the time course of lymphangiogenesis, its response to anti-inflammatory treatment or its pathophysiological relevance in chronic arthritis. Recently, the identification of podoplanin, a specific marker expressed on lymphatic endothelial cells (LECs), but not blood endothelial cells, enabled detailed studies on the distribution of LECs by immunohistochemical localisation; this provides new insights into pathological lymphangiogenesis.10
In this study, we identified the dynamics of lymphatic neovascularisation in experimental arthritis as well as human inflammatory arthritis. In addition, we investigated whether anti-inflammatory treatment with tumour necrosis factor (TNF) blocking agents alters the lymphatic vasculature upon resolution of synovial inflammation.
PATIENTS AND METHODS
Animals and treatment
The human TNF transgenic mice (hTNFtg, tg197 strain) have been described previously.11 These mice develop a destructive polyarthritis before 5–6 weeks of age. Two separate experiments were carried out in this study and a total of 33 mice were included. In the first experiment, hTNFtg mice were sacrificed at different time points (n = 3–5 per group). In the second experiment, hTNFtg mice were either treated with a chimeric anti-TNF antibody (infliximab, Centocor, The Netherlands) at a dose of 10 mg/kg body weight, intraperitonally, three times weekly or received placebo (saline) (n = 5/group). Treatment was started early (6-week-old mice) or late (10-week-old mice). After 6 weeks of treatment, mice were sacrificed and hind paws were obtained for histology as described previously.12 Serial sections (2 μm) were stained with haematoxylin and eosin to analyse quantitatively synovial inflammation. All animal procedures were approved by the local ethical committee.
Patients and biopsies
Sixteen patients with RA and 16 patients with SpA were studied (ankylosing spondylitis n = 7, psoriatic arthritis n = 7, undifferentiated SpA n = 2); they fulfilled either the American College of Rheumatology criteria for RA or the European Spondyloarthropathy Study group criteria. All patients presented with an actively inflamed knee joint and underwent needle arthroscopy at baseline of the study as described previously.13 Subsequently, paired synovial samples were obtained from all patients after treatment with a chimeric anti-TNF antibody (infliximab) was commenced. Patients received either 3 mg/kg body weight (RA) or 5 mg/kg body weight (SpA) infliximab, intravenously, at weeks 0, 2 and 6, after which they underwent arthroscopy of the same knee joint with repeat biopsy at week 4 (RA) or week 12 (SpA). Synovial biopsies were either fixed in 4% p-formaldehyde and then paraffin-embedded (SpA) or snap-frozen (RA). Clinical characteristics and activity measurements were obtained from all patients at week 0 and 16 and are depicted in tables 1 and 2. All procedures were approved by the local ethics committee and all patients gave written informed consent.
Immunohistochemistry
Immunohistochemistry was performed on formalin-fixed, paraffin-embedded sections of synovial biopsies of patients with SpA and on acetone-fixed frozen sections of patients with RA. For immunohistochemical detection, the following antibodies were used: anti- vascular endothelial growth factor (VEGF)-C (Zymed, San Francisco, CA, USA), anti-CD31 (Dako, Glostrup, Denmark) and anti-podoplanin (Pathology, Medical University of Vienna, Austria) or isotype and concentration-matched irrelevant antibody (negative control). CD31 was chosen as a pan-endothelial marker, as other endothelial markers such as CD14614 have not been tested for their reactivity on lymphatic vessels. Antigen retrieval was performed with citrate buffer for paraffin-embedded sections. Non-specific binding was blocked by the addition of 10% rabbit serum for 10 min at room temperature followed by incubation with the primary antibody for 1 h at room temperature. Sections were then incubated with species-specific biotinylated immunoglobulins (Vector) for 30 min and afterwards rinsed with phosphate-buffered saline. Visualisation of staining was performed with streptavidin-biotin (Dako).
Histomorphometry
All analyses were performed using a microscope (Nikon, Duesseldorf, Germany) equipped with a digital camera and an image analysis system (Osteomeasure; OsteoMetrics, Decatur, GA, USA), as described previously.12 The area of synovial inflammation in arthritic hTNFtg mice was quantified in haematoxylin and eosin-stained paw sections. For the determination of microvessel density and lymphatic microvessel density (LMVD), three areas of inflammation with the greatest number of vessels were selected (hot spot) and capillaries were counted in this area. Positive counts were determined at a magnification of 200×. The percentage of VEGF-C positive pannus cells was counted in the area of highest density at a magnification of 400×. All analyses were carried out by a single investigator in blinded fashion.
Statistical analysis
Data are presented as the mean ± SEM. Group mean values were mean±SEM compared by Wilcoxon’s matched pairs test.
RESULTS
Lymphatic vessel formation in the synovium of arthritic mice
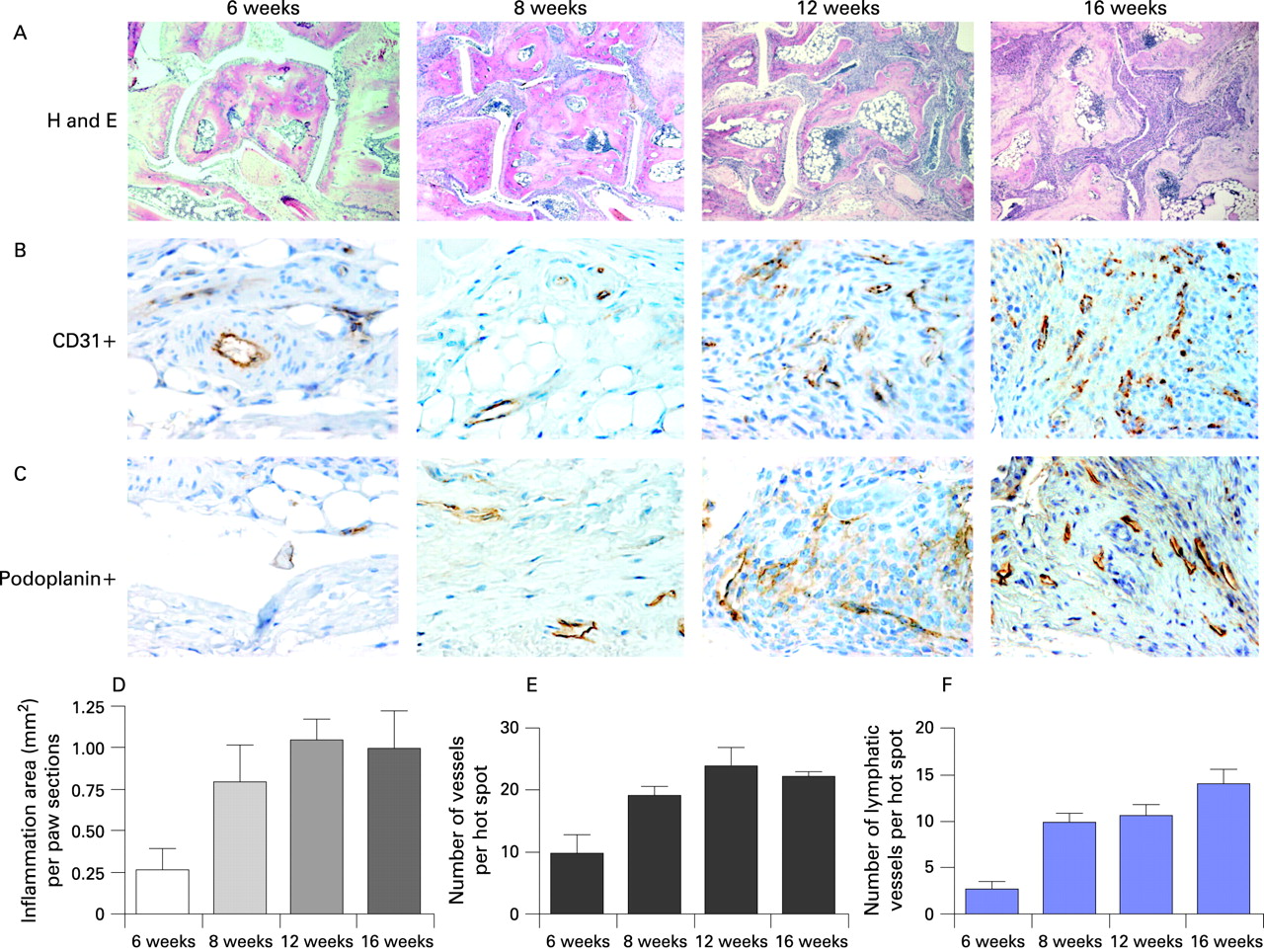
First, we asked whether chronic arthritis triggers lymphatic neovascularisation in the synovium. To determine the evolution of synovial neovascularisation, we analysed arthritic mice at four different time points to assess all stages of arthritis in this animal model of RA. We quantitatively assessed the total area of synovial inflammation (fig 1A) and total density of synovial microvessels by immunohistochemical staining with an anti-CD31 antibody (fig 1B), a marker for all endothelial cells regardless of its origin. The expression of lymphatic vessels was performed by the analysis of podoplanin, which is a specific marker for lymphatic vessels that does not appear on other blood vessels.

Untreated hTNFtg mice progressively developed a chronic proliferative arthritis. Six-week-old mice had only minimal signs of arthritis (0.13±0.07 mm2), whereas inflammation progressively developed from 8 weeks (1.42±0.07 mm2) to 12 weeks of age (2.53±0.14 mm2, p<0.05 versus 8 weeks) and then did not further increase up to 16 weeks of age (2.51±0.08 mm2, p = NS versus 12 weeks). Representative sections and quantification of results are shown in fig 1A. When assessing total microvascular density, significant numbers of CD31-positive vessels already arose in early arthritis (6-week-old hTNFtg mice: mean vessel density 9.8±3.0 per hot spot). Consistent with increasing inflammation, the synovial vessel density steadily increased from 8 weeks (mean 19.0±1.5 LMVD; p<0.05 versus 6 weeks) to 12 weeks of age (mean 24.1±2.7 vessels) but did not further evolve in the late stage of arthritis (16-week-old hTNFtg mice: mean 22.1±0.8 LMVD; p = NS versus 12 weeks). Representative sections and quantification results are shown in fig 1B.
When we assessed the LMVD, we observed only low numbers of podoplanin-positive LECs in 6-week-old hTNFtg mice (mean 2.7±0.9 vessels). However, lymphatic neovascularisation then rapidly emerged resulting in a significant increase in podoplanin-expressing vessels at 8 weeks of age (mean 9.9±1.0 vessels; p<0.05 versus 6 weeks). Further progression of arthritis had no significant impact on lymphatic vessel density (12-week-old hTNFtg mice: 10.6±1.3 vessels; 16-week-old hTNFtg mice: 14.1±1.6 vessels; p = NS versus 8 weeks). Representative sections and quantification results are shown in fig 1C. Interestingly, lymphatic vessels showed no specific predilection site in the inflamed synovium, but were spread throughout the synovium. However, when lymphatic vessels were observed, they were mostly present as groups of vessels. Thus, synovial inflammation effectively induces lymphatic neovascularisation, constituting about 50% of all vessels found within the synovial tissue of hTNFtg mice.
Increased lymphatic vessel density in arthritic mice treated with infliximab
Next, we tested the consequence of anti-TNF therapy on the reorganisation of LMVD in murine inflamed synovial tissue. We thus compared untreated and treated hTNFtg mice. Treatment with a TNF-blocking antibody (infliximab) was carried out either early (weeks 6–12) or late (weeks 10–16; fig 2). As expected, early anti-TNF treatment led to a significant decrease of synovial inflammation (mean reduction versus control 63%, p<0.05); late treatment had a milder effect on arthritis (mean reduction versus control 47%, p<0.05). Surprisingly, increased formation of lymphatic vessels upon treatment with infliximab was observed, regardless of whether treatment was commenced at early or late stages of arthritis. All mice treated with infliximab showed a significant increase of lymphatic vessels in the inflamed areas when analysed at week 12 (LMVD untreated: 10.6±1.3 versus treated 14.6±2.1); p<0.05) or week 16 (untreated: 14.1±1.6 versus treated 23.3±3.9; p<0.05). Representative sections and quantitative results are shown in fig 2. Thus, blockade of synovial inflammation with anti-TNF does not induce regression of LMVD in arthritic lesions but, in contrast, stimulates new lymphatic vessel formation.

Clinical effects of infliximab treatment in patients with active rheumatoid arthritis and spondyloarthritis
In order to determine lymphatic neovascularisation in human inflammatory arthritis, sequential synovial biopsy specimens from patients with active RA (n = 16) and SpA (n = 16) were assessed. All patients underwent needle arthroscopy of swollen joints before and at week 4 in RA and week 12 in SpA after initiation of TNF-blocking therapy with a chimeric anti-TNF antibody (infliximab, Centocor) at the usual doses in the respective diseases. As depicted in tables 1 and 2, all patients were active at study entry and, in concordance with clinical trials of infliximab treatment in human RA and SpA, improved significantly with regard to both clinical and laboratory parameters.
Increased lymphatic vessel formation in human inflammatory arthritis after treatment with infliximab
Immunohistochemical quantification of blood and lymphatic vessels in synovial specimens from patients with RA revealed abundant CD31-positive vascular structures, a large proportion of which was identified as lymphatic due to the expression of podoplanin in the inflamed tissue of pre-treatment samples. In addition to lymphatic endothelium, podoplanin was also found to be expressed in the intimal lining layer. Lymphatic vessels were mostly found in the vicinity of fibroblast-like synoviocytes and macrophages.
Consistent with our data in hTNFtg mice, treatment with infliximab led to the proliferation of synovial vessels in patients with RA (fig 3). This increase in CD31-positive microvessels (MVD per hot spot before treatment: 38.8±6.2, after treatment: 58.8±6.4; p<0.05) was mediated by an emergence of new lymphatic vessels in the rheumatoid synovium. Statistical analysis of LMVD expression revealed a significant difference in podoplanin expression in patients with RA before (11.0±1.6) versus after treatment with infliximab (LMVD: 23.5±2.0; p<0.05). In addition, we evaluated the expression and change upon treatment of VEGF-C (the major lymphangiogenic growth factor) by immunohistochemical analysis. We could observe VEGF-C expression mostly in the intimal lining layer. Interestingly, though not statistically significant, VEGF-C expression was increased after administration of infliximab.
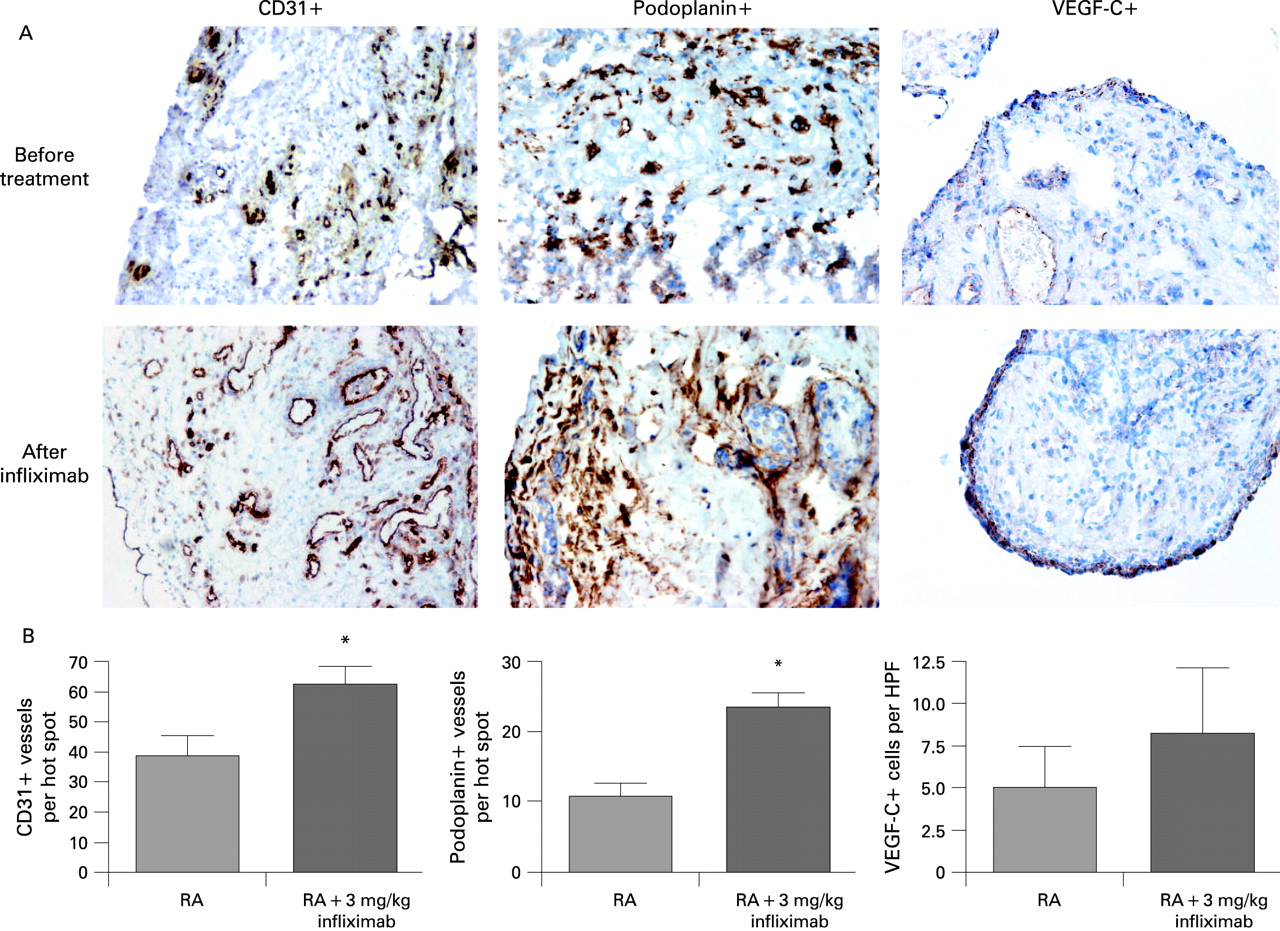
To get an idea of synovial neovascularisation in other inflammatory autoimmune diseases, inflamed synovial tissue from patients with SpA before and after treatment with infliximab was analysed for lymphatic and blood vessel density (fig 4A). Comparable with patients with RA, the synovial tissue of SpA revealed abundant microvessels (approximately 50% were of lymphatic origin). As seen in RA, increased numbers of lymphatic vessels could be observed after treatment with infliximab (before treatment: 9.5±3.5, after treatment: 17.0±2.0, p<0.05; fig 4B). These data suggest that treatment with an anti-TNF antibody increases LMVD in human inflammatory arthritis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
Neovascularisation of the synovium is regarded as a crucial pathogenic step in RA. However, synovial microvessels might not only harbour blood cells constituting an afferent arm to the joint, but may also form the basis of an efferent arm being lymphatic vessels. The role of lymphatic vessels in the arthritic synovium is as yet unclear. In this study we examined the expression of lymphatic vessels in the arthritic synovium and the effects of TNF blockade on synovial vascularity both in animal and human arthritis. We show that (a) intense de novo lymphangiogenesis occurs in chronic arthritis, and (b) the effective treatment of arthritis with TNF blockade decreasing synovial inflammation further increases LMVD in the synovium.
Recent evidence suggests an important role for lymphatic vessels in acute and chronic inflammation. Elegant studies in patients suffering from allograft rejection have revealed the role of lymphatic neoangiogenesis at sites of tissue rejection such as the cornea and the kidney.15–17 In these conditions, acute and chronic inflammation provokes the production of growth factors for lymphatic vessels such as VEGF-C and lead to intense de novo lymphangiogenesis. The origin of the LECs is not yet clear, but might relate at least partially to macrophages undergoing transdifferentiation.18 Interestingly, macrophages do not only produce but also respond to VEGF-C with transdifferentiation and phenotypic change towards a LEC type expressing specific LEC markers such as podoplanin and Prox-1.
RA is a prototype of a chronic immune-mediated inflammatory disease associated with the formation of a heavily vascularised synovial tissue. Little is known about the presence, role and origin of lymphatic vessels in RA as specific markers for lymphatic vessels such as podoplanin, LYVE-1 and Prox-1 have been identified only recently. Previous studies investigating the presence of lymphatic vessels in the rheumatoid synovium reported conflicting results: some authors could not find lymphatic vessels in patients with RA, other studies found lymphatic vessels both in osteoarthritis as well as in inflammatory arthritis.19–21 To investigate whether synovial lymphatic vascularisation occurs in animal and human arthritis, we used a specific marker for LECs, podoplanin, which is not found on vascular endothelial cells. By investigating synovial biopsy specimens of patients with RA we could detect lymphatic neovascularisation in the affected joints. Lymphatic neovascularisation is also relevant in other forms of chronic arthritis as we were able to detect very similar changes in peripheral arthritis in conjunction with SpA.
The in vivo findings in patients with chronic arthritis were confirmed by our studies in hTNFtg mice allowing us to determine the kinetics of lymphatic vessel formation in the joint. We found that lymphatic neovascularisation occurs significantly later than the appearance of blood vessels in arthritic joints, whereas both vessel types then steadily increase in later stages of arthritis. Interestingly, the lymphatic vessels do not seem to be evenly distributed in the arthritic synovium but are rather grouped together. The factors triggering lymphatic vessel growth in chronic arthritis are as yet unclear. However, proinflammatory cytokines such as TNF-α and interleukin-1 are potent inducers of VEGF-C, the major lymphangiogenic growth factor, in synovial fibroblasts in vitro.7 Consequently, we could find distinct VEGF-C expression in the rheumatoid synovium, especially in the intimal lining layer.
The functionality of the synovial lymphatic system is currently not understood. In general, the lymphatic system collects extracellular fluid and returns it to the venous circulation.22–24 However, under certain pathological conditions, lymphatic neovascularisation may occur adjacent to tumours allowing the metastatic spread of tumour cells and progression of the disease. This demonstrates not only a role of lymphatic vessels for fluid collection but also active transportation of cells. Indeed, chronic inflammation such as found in renal allograft rejection is also a potent trigger for lymphatic neovascularisation. Interestingly, these vessels arise next to inflammatory infiltrates mainly consisting of T and B cells as well as dendritic cells.15 Moreover, the LECs produce chemokines attracting immune cells into the lumen.9 Thus, lymphatic vessels may actively recruit immune cells resulting in either drainage to the next lymph node or clearance of potentially dangerous cells. The latter is being supported by the fact that increased LMVD in kidney allograft is independently associated with a better clinical outcome.25
Whether these findings also apply to chronic arthritis remains to be elucidated but it is intriguing that we did not observe a decrease in LMVD upon effective anti-inflammatory treatment. In contrast, we found an increase in lymphatic neovascularisation both in experimental and human arthritis after blockade of TNF. Although lymphatic vessels form in conjunction with chronic arthritis, they rather follow blood vessels and may be seen as a regulatory mechanism to allow efflux of fluid and cells out of the joint. This mechanism appears to be facilitated by the effective treatment of RA, such as with TNF-blocking agents and might be seen as the basis for reduced swelling and stiffness upon the resolution of arthritis. Supporting these findings, it was recently shown that an increased capacity of local lymph nodes to drain inflammatory cells is associated with decreased synovial inflammation in TNF-driven experimental arthritis.26 Also, macrophages are known to have the capacity to transdifferentiate into LECs under inflammatory conditions. Thus, an explanation for these findings could be that evading macrophages undergo transdifferentiation, form lymphatic endothelia and thereby contribute to the new formation of lymphatic vessels. Our findings might thus explain how effective anti-inflammatory treatment reduces synovial inflammation. Given the fact that apoptosis does not seem to be a major contributor to the rapid and sustained reduction of synovial inflammatory cells during effective treatment of RA,27 increased efflux of fluid and cells due to lymphatic neovascularisation might be a valid alternative explanation.
In conclusion, we could show significant lymphatic neovascularisation in chronic arthritis, which follows the spreading of blood vessels. TNF-blocking therapy increases lymphatic vessel formation in the synovium, opening the possibility for an increased clearance of inflammatory cells and fluid out of the joint. This mechanism may be the basis for the reduction of joint swelling during the effective treatment of inflammatory arthritis.
Acknowledgments
We thank Birgit Tuerk for excellent technical assistance. This study was supported by the START prize of the Austrian Science Fund (GS). DMG was supported by the Dutch Arthritis Association. PPT and DB received support from the European Community’s FP6 funding (Autocure). This publication reflects only the author’s views. The European Community is not liable for any use that may be made of the information herein.
REFERENCES
Footnotes
Competing interests: None.