Article Text

Abstract
Objective: To determine whether oncostatin M (OSM) + tumour necrosis factor α (TNFα) induces aggrecanase activity in chondrocyte membranes, to determine the effects of transforming growth factor β1 (TGFβ1), interleukin 4 (IL4), and tissue inhibitor of metalloproteinases (TIMPs) on this activity, and to determine whether this activity is due to a known ADAMTS aggrecanase.
Methods: Aggrecanase activity and ability of agents to prevent membrane associated aggrecanase activity were assessed by Western blotting. Expression of known aggrecanases was measured by real time polymerase chain reaction in bovine nasal and human articular chondrocytes.
Results: Chondrocyte membrane associated aggrecanase activity and increased mRNA expression of ADAMTS-1, -4, -5, and -9, but not ADAMTS-4 or -15, were enhanced after stimulation by OSM+TNFα in bovine chondrocytes. This activity was inhibited by TIMP-3. In human chondrocytes, OSM+TNFα also enhanced ADAMTS-1 and -4 expression, but not that of other ADAMTSs. TNFα alone induced ADAMTS-9 expression, whereas OSM addition caused suppression. Both TGFβ1 and IL4 blocked membrane associated aggrecanase activity and decreased OSM+TNFα-induced expression of ADAMTS-9 in bovine and human chondrocytes. IL4 down regulated ADAMTS-4 mRNA, whereas TGFβ1 increased this expression in both bovine and human chondrocytes.
Conclusions: OSM+TNFα up regulates membrane associated aggrecanase activity and several ADAMTS aggrecanase mRNAs in chondrocytes. The chondroprotective effects of IL4 and TIMP-3 suggest that they may have therapeutic benefit for aggrecanolysis, whereas the differential inhibitory effects of TGFβ1 may limit its therapeutic potential. Induced membrane associated aggrecanase activity is distinct from known soluble ADAMTS aggrecanases and merits further investigation.
- ADAMTS, A Disintegrin And Metalloproteinase ThromboSpondin motif
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- IL, interleukin
- MMPs, matrix metalloproteinases
- OA, osteoarthritis
- OSM, oncostatin M
- PBS, phosphate buffered saline
- PCR, polymerase chain reaction
- PMSF, phenylmethylsulphonyl fluoride
- RA, rheumatoid arthritis
- TFGβ, transforming growth factor β
- TIMPs, tissue inhibitors of metalloproteinases
- TNFα, tumour necrosis factor α
- aggrecanase
- ADAMTS
- TIMP
- chondrocytes
- membrane
Statistics from Altmetric.com
- ADAMTS, A Disintegrin And Metalloproteinase ThromboSpondin motif
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- IL, interleukin
- MMPs, matrix metalloproteinases
- OA, osteoarthritis
- OSM, oncostatin M
- PBS, phosphate buffered saline
- PCR, polymerase chain reaction
- PMSF, phenylmethylsulphonyl fluoride
- RA, rheumatoid arthritis
- TFGβ, transforming growth factor β
- TIMPs, tissue inhibitors of metalloproteinases
- TNFα, tumour necrosis factor α
Cartilage degradation leading to a loss of joint function in the arthritides involves proteolytic cleavage of both aggrecan and collagen. Loss of aggrecan from the cartilage extracellular matrix is an early and sustained feature of joint degeneration as is seen in both osteoarthritis (OA) and rheumatoid arthritis (RA). Matrix metalloproteinases (MMPs) cleave the aggrecan core protein at the Asn341–Phe342 bond,1 but the proteolytic fragments generated are retained within the tissue, suggesting that MMPs may not be responsible for pathological aggrecan degradation.2 Proteolysis at the Glu373–Ala374 bond within the interglobular domain of aggrecan is mediated by proteinases called aggrecanases,1 which release the glycosaminoglycan-rich C-terminus of aggrecan that is important for the mechanical properties of cartilage. Aggrecanases are metalloproteinases that belong to a subfamily of the A Disintegrin and Metalloproteinase (ADAM) proteinases that contain thrombospondin motifs, the ADAMTSs. Although aggrecanase-1 and -2 (ADAMTS-4 and -5) were first reported and implicated in cartilage degradation,3,4 ADAMTS-1, -4, -5, -8, -9, and -15 are now referred to as aggrecanases.3–,8 Aggrecan fragments generated by aggrecanase activity have been identified in cartilage explant cultures in vitro, as well as in human synovial fluid and cartilage tissues.2,9,10
Data implicating ADAMTS-4 and -5 in aggrecanolysis are, however, conflicting and contradictory. Some studies on aggrecanase expression have found raised mRNA levels, whereas others have failed to see regulation even after cytokine stimulation.11–,15 It has been suggested that both these aggrecanases are important in OA, based on in vitro observations with a hydroxamate compound that inhibits both MMPs and aggrecanases,16 while mRNA profiling data for OA have suggested that ADAMTS-5 and not ADAMTS-4 is the pathophysiologically relevant aggrecanase,13 and decreased levels of ADAMTS-1, -5, and -9 have also been reported.17 ADAMTS-4 mRNA is up regulated by cytokines such as interleukin 1 (IL) 1 and tumour necrosis factor (TNFα), while ADAMTS-1 is down regulated by IL1.18,19 ADAMTS-5 mRNA has been shown to be expressed in synovium, although it is not regulated by either IL1 or retinoic acid.12 ADAMTS-1 and -8 are anti-angiogenic,20 suggesting that they are likely to have anabolic effects on cartilage. ADAMTS-9 is expressed in embryonic and adult tissues, and to date no physiological roles have been ascribed to ADAMTS-9 or -15.7,21
TNFα is an important mediator in RA pathogenesis, and anti-TNFα treatment reduces inflammation, joint destruction, and serum MMP levels.22 This proinflammatory cytokine stimulates cartilage aggrecan and collagen release and inhibits their synthesis by chondrocytes.23,24 Furthermore, synovial fluid TNFα levels correlate with cartilage degradation.25 Oncostatin M (OSM) is known to induce joint inflammation and cartilage damage,26–,28 and has been localised to macrophages in rheumatoid synovium.28 Moreover, raised levels of OSM are detected in RA synovial fluids,28 and these levels correlate with joint inflammation and the markers of aggrecan degradation in RA.25 Blockade of OSM ameliorates joint inflammation and cartilage damage in collagen induced arthritis.30 Furthermore, OSM is an important co-factor in both IL1 and TNFα mediated cartilage and bone damage and inflammation in arthritis.26,27
IL4 and transforming growth factor β1 (TGFβ1) are both major chondroprotective factors which can protect cartilage from degradation.30–,34 IL4 suppresses synovial IL1 levels and prevents the activation of pro-MMPs.30,31 TGFβ is an important anabolic regulator of cartilage34; it stimulates the synthesis of matrix components, and blocks cytokine induced degradation by down regulating MMPs and up regulating tissue inhibitors of metalloproteinases (TIMPs).33–,35 Recently, TGFβ has been found to stimulate ADAMTS-4 expression and aggrecanolysis in cartilage.15
TIMPs are important metalloproteinase regulators in extracellular matrix biology, and four have been identified in human synovium and cartilage that are regulated by growth factors and cytokines.35 TIMP-3 potently inhibits ADAMTS-4 and -5,36,37 and blocks aggrecanolysis from bovine and porcine cartilages.38 Conversely, the other TIMPs which efficiently inhibit many MMPs, are poor inhibitors of ADAMTS-4 and -536,37 and fail to prevent aggrecanolysis, although all TIMPs block cartilage collagenolysis.39
We have shown that OSM in combination with IL1 or TNFα induces synergistic cartilage proteoglycan degradation in vitro and in vivo,26–29,40 and that IL1+OSM up regulates a chondrocyte membrane associated aggrecanase activity.41 In this study we investigated whether this membrane associated aggrecanase activity can be induced by OSM+TNFα, if TGFβ, IL4 or TIMPs can modulate this activity, and whether this activity can be attributed to one of the known ADAMTS aggrecanases.
MATERIALS AND METHODS
Chemicals were obtained from the following suppliers: human recombinant TNFα, IL4, and TGFβ1 were purchased from R&D Systems (Abingdon, UK). OSM was a gift from Professor JK Heath (Birmingham, UK). The polyclonal antibody R663 was raised in a rabbit against the keyhole limpet haemocyanin conjugated peptide Ala-Arg-Gly-Ser-Val-Ile-Cys-Gly-COOH, corresponding to the amino terminus generated by cleavage of aggrecan at the Glu373–Ala374 peptide bond, and was generously supplied by Dr DJ Buttle (University of Sheffield, UK).42 Recombinant human TIMP-1 and TIMP-3, and native human TIMP-2 were produced in-house.43–,45 Primers and probes for human glyceraldehyde-3-phosphate dehydrogenase (GAPDH) and bovine ribosomal 18S were purchased from Applied Biosystems (Warrington, UK), and all other oligonucleotides were obtained from Sigma (Poole, UK). Chondroitinase ABC (catalogue No 320301) and keratanase (catalogue No 320321) were purchased from ICN Biomedicals Ltd (Thame, UK). All other chemical and biochemicals were commercially available analytical grade reagents obtained from Fisons (Loughborough, UK) or Sigma, or have been previously described.26–,28
Chondrocyte culture and membrane preparation
Chondrocytes were isolated from bovine nasal septum cartilage samples and human knee joints, as previously described.26–,28 Cells were incubated in fresh serum-free Dulbecco’s modified Eagle’s medium containing 25 mM HEPES (Gibco, Paisley, UK) supplemented with glutamine (2 mmol/l), streptomycin (100 μg/ml), penicillin (100 U/ml), amphotericin B (2.5 μg/ml), and 10% (vol/vol) fetal calf serum in a humidified atmosphere of 95% air/5% CO2 at 37°C. When cells reached 80–90% confluence, the medium was removed and replaced with serum-free medium overnight and then cultured in fresh medium containing test cytokines for the indicated time periods.
Chondrocyte membrane purification was adapted from that previously described.41 Briefly, media were removed and the stimulated bovine nasal chondrocyte monolayer was washed twice with ice cold phosphate buffered saline (PBS), harvested into PBS using a scraper, and the cell pellet resuspended in lysis buffer (10 mM Tris/HCl, pH 7.4, containing 8.5% (wt/vol) sucrose, 1 mM N-ethylmaleimide, 1 mM phenylmethylsulphonyl fluoride (PMSF), and 10 µg/ml soybean trypsin inhibitor). Cells were lysed by sonication, and unlysed cells and cell nuclei were removed by centrifugation at 1700 g for 10 minutes at 4°C. The supernatant was collected and then centrifuged at 33 000 g for 1.5 hours at 4°C to pellet the membrane fraction, which was resuspended in 1 ml of lysis buffer. After centrifugation at 150 000 g for 1.5 hours at 4°C and layering of 0.5 ml of the sample on top of 10 ml of 17% (wt/vol) sucrose, the resultant pellet was resuspended in 10 mM Tris/HCl, pH 7.4, assessed for 5’-nucleotidase activity as an indicator of plasma membrane enrichment,46 and stored at −20°C.41 Protein concentration was determined by the BCA protein assay (Pierce, Rockford, Illinois, USA).
Aggrecan preparation
Aggrecan was prepared from bovine nasal septum cartilage using a modification of an established protocol.47 Briefly, cartilage was cut into 2 mm3 pieces and added to 10 volumes of extraction buffer (4 M guanidinium chloride, 50 mM sodium acetate, pH 6.0, 2 mM PMSF, 5 mM benzamidine-HCl, 10 mM EDTA, 0.1 mM 6-aminohexanoic acid) at 4°C. The mixture was stirred for 48 hours at 4°C, and aggrecan isolated by 1.45 g/ml caesium density gradient centrifugation at 100 000 g for 48 hours at 10°C. Purified aggrecan was dialysed exhaustively into 10 mM Tris/acetate, pH 7.4, before storage at −20°C.41 Aliquots of the aggrecan were treated with chondroitinase ABC lyase at 2.5 mU/mg of proteoglycan, and keratinase at 50 mU/mg of proteoglycan, in the presence of proteinase inhibitors for 5 hours at 37°C to remove chondroitin and keratan sulphate chains. This deglycosylated aggrecan was dialysed against 10 mM Tris/HCl, pH 7.4, and stored at −20°C, and used in all experiments as the aggrecanase substrate.41
Chondrocyte membrane associated aggrecanase activity assay
The membrane preparation (50 µg of protein) was incubated in the presence of the bovine aggrecan substrate (100 µg protein) in Tris buffer (pH 7.4) containing 10 mM CaCl2 for 16 hours at 37°C.41 Samples were then boiled for 5 minutes and subjected to electrophoresis on 10% sodium dodecyl sulphate/polyacrylamide gel electrophoresis gels and the separated proteins transferred to nitrocellulose. After blocking, the membrane was incubated overnight with anti-(aggrecan neoepitope)antibody (R663) diluted 1:200 in Tris buffered saline containing 0.1% (vol/vol) Tween 20 with 1% (wt/vol) milk powder. The blots were then incubated for 1 hour with anti-(rabbit IgG)-horseradish peroxidase conjugate diluted 1:1000. The cleaved aggrecan was visualised by enhanced chemiluminescence (Amersham Biosciences, Little Chalfont, UK).
Real time polymerase chain reaction (PCR)
Total cellular RNA was isolated and purified using the RNeasy kit (Qiagen, Crawley, UK). RNA (1 μg) was reverse transcribed in a 20 μl reaction using random hexamers and Superscript II reverse transcriptase (Invitrogen, Paisley, UK), according to the manufacturer’s instructions.48 Oligonucleotide primers were designed using Primer Express 1.0 software (Applied Biosystems). To prevent amplification of any contaminating genomic DNA, the primers were located within different exons close to, or spanning, the intron/exon boundary. Table 1⇓ outlines the primer sequences.
Primers sequences
Relative measurement of human gene expression was performed using a Lightcycler (Roche Diagnostics Ltd, Lewes, UK) or an ABI 7900HT. PCRs were performed in triplicate in 5 mM MgCl2, 2 μl of Hotstart SYBR Green Master Mix (Roche), 0.5−1.0 μmol/l of each primer in a 20 μl reaction. Thermocycler conditions comprised an initial activation step at 95°C for 10 minutes. This was followed by a three step programme consisting of 95°C for 20 seconds, 60°C for 5 seconds, and 72°C for 10 seconds for 45 cycles. A one step melt curve analysis was also performed at the end of each run to ensure the crossover values obtained were due to the amplification of a single specific product.48 For TaqMan reactions, VIC or FAM labelled probes were used with an initial step of 50°C for 2 minutes, then 95°C for 10 minutes, followed by a two step programme of 95°C for 15 seconds and 60°C for 1 minute for 45 cycles, according to the manufacturer’s instructions. Bovine genes were assessed using SYBR Green (Cambrex, Wokingham, UK), essentially as above after optimisation.
RESULTS
OSM in combination with TNFα induces a chondrocyte membrane associated aggrecanase activity that is inhibited by TIMP-3
Stimulation of bovine nasal chondrocytes with OSM or TNFα alone induced very low levels of the membrane associated aggrecanase at 24 hours, although for TNFα this level of activity increased at 48 hours (not shown). When OSM was combined with TNFα, a marked dose dependent increase in membrane associated aggrecanase activity was found (fig 1A⇓). The pattern of aggrecan fragments generated containing the aggrecanase neoepitope was very similar to that previously reported for IL1+OSM.41 Inclusion of IL4 or TGFβ1 with OSM+TNFα resulted in a marked reduction of the membrane associated aggrecanase activity while both of these anti-inflammatory agents failed to induce the activity when used alone (fig 1B⇓).

Effect of cytokines and TIMPs on chondrocyte membrane associated aggrecanase activity. Bovine nasal chondrocytes were cultured until 80–90% confluence, serum starved overnight, and then stimulated with various cytokines individually or in combination for 24 hours. Harvested cells were lysed and the plasma membrane fraction enriched by density centrifugation. Membranes (50–100 µg) were incubated with deglycosylated aggrecan (100 µg) at 37°C for 16 hours. Aggrecanase activity was examined by Western blot using an antibody (R663) that recognises the neoepitope generated after cleavage of aggrecan at the Glu373–Ala374 bond. (A) Effect of cytokine dose on the membrane activity; (B) effect of TGFβ1 (15 ng/ml) or IL4 (25 ng/ml) inclusion in the 24 hour chondrocyte stimulation; (C) the effect of TIMP-1, -2, and -3 (0.1, 0.5, or 1.5 µg), added for 30 minutes at room temperature before the addition of substrate. The data shown are representative of three separate chondrocyte preparations.
Neither TIMP-1 nor TIMP-2 caused any significant inhibition of the membrane associated aggrecanase, as was previously reported for IL1+OSM.41 However, inclusion of TIMP-3 almost completely inhibited this activity even at the lowest concentration used (fig 1C⇑), suggesting that this activity is a metalloproteinase such as an ADAM or ADAMTS.
OSM in combination with TNFα induces the expression of ADAMTS aggrecanases in chondrocytes
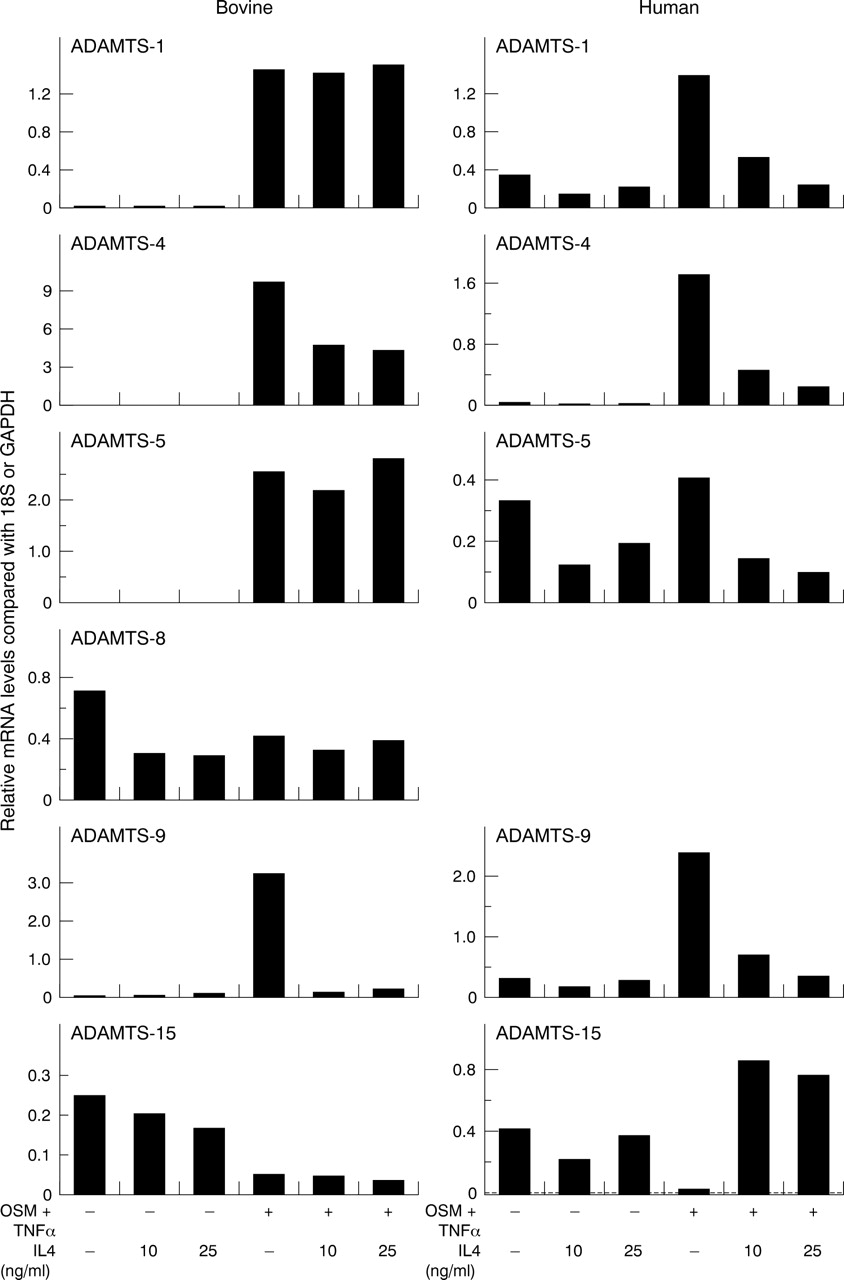
In bovine chondrocytes, OSM+TNFα enhanced ADAMTS-1, -4, -5, and -9 expression while ADAMTS-8 expression was relatively unaffected. All treatments appeared to reduce ADAMTS-15 expression compared with control (fig 2⇓, bovine). In human articular chondrocytes, ADAMTS-1 and -4 mRNA appeared to be dose dependently up regulated by the OSM+TNFα combinations at 24 hours. A minor, but consistent induction of ADAMTS-4 mRNA was seen after either OSM or TNFα stimulation. ADAMTS-5 was constitutively expressed, and this expression was reduced by both OSM and TNFα. The OSM+TNFα combination failed to increase this expression above basal levels. TNFα enhanced ADAMTS-9 expression, although inclusion of OSM markedly decreased this TNFα stimulated expression. Cytokines alone, and in combination, appeared to decrease ADAMTS-15 expression compared with control (fig 2⇓, human). We failed to detect ADAMTS-8 mRNA in human chondrocytes (not shown). Similar profiles were also found at 6 and 12 hour time points for both bovine and human chondrocytes (data not shown).

Effects of OSM and TNFα on the expression of ADAMTS aggrecanases in chondrocytes. Bovine nasal or human articular chondrocytes were cultured until 80–90% confluence and then serum starved overnight. Cells were stimulated with OSM, TNFα alone or in combination for 24 hours. Total RNA was isolated, reverse transcribed, and the resulting cDNA used in a separate real time PCR with specific primers for ADAMTS-1, -4, -5, -8, -9, or -15. No signal was detected for ADAMTS-8 in human chondrocytes with any treatment. Results were normalised to either 18S (for bovine) or GAPDH (for human) and are presented graphically as relative mRNA levels.48 The data shown are representative of two separate chondrocyte preparations.
TGFβ1 and IL4 have differential effects on OSM+TNFα-induced ADAMTS aggrecanase mRNA expression in chondrocytes
Addition of TGFβ1 to bovine chondrocytes increased both basal and OSM+TNFα stimulated levels of ADAMTS-1, -4, and -5. Basal levels of ADAMTS-8 and -9 were unaffected while the relatively low levels of ADAMTS-15 were suppressed. OSM+TNFα-induced levels of ADAMTS-8 and -15 were not notably affected by TGFβ1, but ADAMTS-9 levels were markedly lower (fig 3⇓, bovine). In human chondrocytes, the relatively low basal ADAMTS expression levels were moderately suppressed by TGFβ1 except for a more marked reduction for ADAMTS-15 (fig 3⇓, human). Again, similar profiles at 6 and 12 hour time points were found (data not shown).

Effects of TGFβ1 on OSM+TNFα-induced ADAMTS aggrecanases. Bovine nasal or human articular chondrocytes were stimulated with OSM+TNFα (both at 10 ng/ml)±TGFβ1 for 24 hours. Real time PCR was performed on isolated RNA, and the normalised results presented graphically as described in fig 2⇑. No signal was detected for ADAMTS-8 in human chondrocytes with any treatment. The data shown are representative of two separate chondrocyte preparations.
IL4 had little effect on basal ADAMTS expression levels in bovine chondrocytes except for a modest suppression for ADAMTS-8 and -15. For OSM+TNFα-induced ADAMTS expression, IL4 had no affect on ADAMTS-1, -5, -8, or -15, but suppressed both ADAMTS-4 and -9 expression (fig 4⇓, bovine). For human chondrocytes, IL4 again had little effect on basal ADAMTS expression except modest suppressions for ADAMTS-1, -5, and -15, although this was more pronounced for OSM+TNFα-induced ADAMTS-1, -4, -5, and -9 but not ADAMTS-15 (fig 4⇓, human). Similar results were observed at 6 and 12 hour time points (data not shown).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of IL4 on OSM+TNFα-induced ADAMTS aggrecanases. Bovine nasal or human articular chondrocytes were stimulated with OSM+TNFα (both at 10 ng/ml) ± IL4 for 24 hours. Real time PCR was performed on isolated RNA, and the normalised results presented graphically as described in fig 2⇑. No signal was detected for ADAMTS-8 in human chondrocytes with any treatment. The data shown are representative of two separate chondrocyte preparations.
DISCUSSION
The breakdown of aggrecan and collagen from cartilage leads to a significant loss of functionality of its extracellular matrix. Collagen loss has been attributed to the collagenolytic MMPs—most notably, MMP-1 and -13, although this is dependent on the degenerative disorder.49 These enzymes are up regulated by a plethora of proinflammatory cytokines, and several cytokine combinations have now been shown to synergistically induce these proteinases.26–28,50 Most of these cytokines also induce proteoglycan loss from cartilage, an event that typically precedes collagen loss.9,28 Several MMPs are known to cleave aggrecan,1 although cleavage at the Glu373–Ala374 bond is considered to be the mechanism of pathological aggrecan degradation in cartilage.2 The discovery of two aggrecanases—namely, ADAMTS-4 and -5, represented a watershed in matrix biology.3,5 However, since these discoveries conflicting data have been reported about the identity of the proteinases that are responsible for aggrecan degradation. Early studies suggested that proteoglycan release from cartilage occurred at the cell surface,51,52 and our own work has demonstrated an aggrecanase activity associated with the chondrocyte membrane.41 Our data also indicate that this activity is not readily solubilised from membrane preparations (unpublished data), suggesting that it may be an integral protein which would exclude the possibility of it being a soluble proteinase such as one of the ADAMTS aggrecanases.3–,8
This study confirms that the combination of OSM+TNFα also induces this membrane associated aggrecanase which has a similar activity against aggrecan to that of other aggrecanases, in that it cleaves aggrecan monomer at multiple sites.1,8 Moreover, this activity is inhibited if either IL4 or TGFβ1, both chondroprotective agents, are present. OSM alone induced weak activity, and although it does not induce any of the ADAMTS aggrecanases it does promote cartilage aggrecanolysis.28 TNFα also mediates aggrecan release,23,24 and induced the membrane associated aggrecanase in this study. Interestingly, in vitro assays using aggrecanase neoepitope recognition failed to detect soluble TNFα-induced aggrecanase activity, although exogenous addition of deglycosylated aggrecan blocked aggrecan release from bovine cartilage.24 Possibly, therefore, TNFα-induced aggrecan release is partly mediated by ADAMTS-4 because this aggrecanase is modestly up regulated by this growth factor, but the synergistic release of proteoglycan from cartilage28 might be explained by the presence of a new aggrecanase.41
TIMP-3 has been shown to inhibit IL1 and retinoic acid mediated proteoglycan release,38 and our data also confirm the ability of TIMP-3 to inhibit the proteolytic action of the chondrocyte membrane associated aggrecanase. However, differences were seen between the IL1 and retinoic acid treatments, which may be explained by the action of hyaluronidases.53,54 Although TIMP-3 has been shown to potently inhibit ADAMTS-4 and -5,36,37 the identity of the specific aggrecanase(s) susceptible to TIMP-3 inhibition in these ex vivo studies remains unknown.38 Our data also indicated that neither TIMP-1 nor TIMP-2 inhibited the membrane associated aggrecanase even at >10-fold molar excess compared with TIMP-3.
Comparison of this membrane associated aggrecanase activity and ADAMTS aggrecanase gene expression in bovine chondrocytes (stimulated under the identical conditions that generate the membrane aggrecanase activity) excludes ADAMTS-1, -4, -5, -8, and -15 because the expression profiles of these enzymes do not coincide with that of the membrane activity. This therefore leaves ADAMTS-9 as a possible candidate as it as been shown that this soluble enzyme can localise to the cell surface.6 However, in cell based assays, ADAMTS-9 cleaves versican at the same Glu–Ala bond as ADAMTS-4, but was barely active against aggrecan compared with ADAMTS-4, although aggrecanase activity at the Glu373–Ala374 bond was not assessed.6 Although this does not definitively preclude ADAMTS-9 as the membrane associated aggrecanase described in the present study, data from human articular chondrocytes also support this exclusion because ADAMTS-9 is induced by TNFα but reduced by OSM. Another important point here is that this membrane associated aggrecanase hydrolyses deglycosylated aggrecan monomer, a property not attributed to aggrecanases such as ADAMTS-4.55
Activation of a constitutively produced aggrecanase such as ADAMTS-9 as a mechanism to explain OSM+TNFα-induced membrane associated aggrecanase activity cannot be discounted. We have previously proposed that OSM contributes to cartilage collagenolysis by inducing potential activators of procollagenases.28 Indeed, although activation of constitutive ADAMTS-4 has been reported for IL1,56 this mechanism is not universally accepted. Glycosylphosphatidyl inositol-anchored MMP-17 has been proposed to contribute to this activation,57 and a furin-like protease has also been suggested; indeed, our own data support the involvement of a furin-like activity in the pathways leading to proteoglycan degradation.58 A furin inhibitor blocks aggrecanase enzyme processing59 and furin levels are raised in diseased cartilage compared with normal.60 It is therefore apparent that aggrecanolysis is a complex process that most probably involves several distinct aggrecanase proteinases, depending on the type of stimulus, species, and age of the cartilage.53
Although TGFβ1 inhibited the membrane associated aggrecanase, we confirm that it induces ADAMTS-4.15 These differential effects may therefore limit the potential therapeutic benefits of this growth factor,34 while IL4 appears to be able to block the action of collagenases30,31 and aggrecanases. Our own data support the notion that a new membrane associated aggrecanase may contribute significantly to proinflammatory cytokine induced aggrecanolysis, and a better understanding of the complexity involved is critical in the identification of new therapeutic targets that prevent cartilage degradation.
Acknowledgments
This work was funded by the Wellcome Trust, the Arthritis Research Campaign, the Dunhill Medical Trust, the Anne Coleman Fund, and the JGW Patterson Trust. We thank John Heath and Dave Buttle for their reagents. There are no competing interests.
REFERENCES
Footnotes
Published Online First 5 May 2005