Article Text

Abstract
Objectives: To investigate potential differences in phenotype and behaviour of immature (iDC) and mature dendritic cells (mDC) from patients with RA and healthy subjects.
Methods: iDC and mDC were derived from blood monocytes of patients with RA and healthy controls following standardised protocols. FACS was used to analyse expression of FcγRI, II, and III and molecules to characterise DC. Discrimination between FcγRIIa and FcγRIIb was achieved by RT-PCR. Immunohistochemistry was performed on synovial biopsy specimens of three patients with RA and three healthy controls. TNFα production by iDC and mDC upon FcγR dependent stimulation was compared between patients with RA and controls by ELISA.
Results: iDC from patients with active RA but not from patients with inactive RA or healthy controls markedly up regulated FcγRII. mDC from patients with active RA also lacked the physiological down regulation of FcγRII that occurs upon maturation in both control groups. RT-PCR analysis confirmed the increased expression of FcγRII in RA—especially marked for FcγRIIb. FcγR dependent stimulation of DC using antigen-IgG immune complexes (IC) significantly increased TNFα production by DC from healthy subjects, but significantly decreased TNFα by DC from patients with RA. Overlapping expression patterns between FcγRII and DC-LAMP in the synovial tissue of patients with RA imply that in vivo, also, mature DC express increased levels of FcγRIIb.
Conclusion: The presence and altered characteristics of DC during active RA suggest that DC help to modulate autoimmunity in RA. Further studies should elucidate the role of local factors in altering the function of DC in RA and in increasing expression of FcγRII.
- DAS28, 28 joint disease activity score
- DC, dendritic cells
- ELISA, enzyme linked immunosorbent assay
- FACS, fluorescence activated cell sorter
- FcγR, Fc gamma receptor
- GAPDH, glyceraldehyde 3-phosphate dehydrogenase
- HAGGs, heat aggregated gamma immunoglobulins
- IC, immune complexes
- IL, interleukin
- LPS, lipopolysaccharide
- MFI, mean fluorescence intensity
- MHC, major histocompatibility complex
- PBMC, peripheral blood mononuclear cell
- RA, rheumatoid arthritis
- RT-PCR, reverse transcriptase-polymerase chain reaction
- TNFα, tumour necrosis factor α
- dendritic cells
- autoimmunity
- Fc gamma receptor
- rheumatoid arthritis
- tumour necrosis factor α
Statistics from Altmetric.com
- DAS28, 28 joint disease activity score
- DC, dendritic cells
- ELISA, enzyme linked immunosorbent assay
- FACS, fluorescence activated cell sorter
- FcγR, Fc gamma receptor
- GAPDH, glyceraldehyde 3-phosphate dehydrogenase
- HAGGs, heat aggregated gamma immunoglobulins
- IC, immune complexes
- IL, interleukin
- LPS, lipopolysaccharide
- MFI, mean fluorescence intensity
- MHC, major histocompatibility complex
- PBMC, peripheral blood mononuclear cell
- RA, rheumatoid arthritis
- RT-PCR, reverse transcriptase-polymerase chain reaction
- TNFα, tumour necrosis factor α
Dendritic cells (DC) are professional antigen presenting cells with the unique ability to activate not only memory but also naive T cells, resulting in the initiation and modulation of both primary and secondary immune responses.1–3 More recently, iDC have been implicated in the induction of immune tolerance.4,5 DC are scattered throughout the body and execute two key functions, which are segregated in time and place. Immature DC reside in the periphery and are specialised in uptake and processing of antigens. Upon encounter of antigens or as a result of inflammation, DC mature and migrate to secondary lymphoid organs.3 Maturation of DC coincides with major changes such as up regulation of CD80, CD86, major histocompatibility complex (MHC)-I and MHC-II, and “de novo” expression of maturation markers such as DC-LAMP and CD83.4,5 During maturation DC normally lose the ability to capture antigens, which is reflected in the down regulation of scavenger receptors.3,6,7 The expression of these markers together with a panel of recently identified antigens which are unique for DC (DC-SIGN, DC-CK1, and DC-LAMP) can be used to characterise the differentiation and maturation of DC.8,9,10,11
Accumulating evidence points to the involvement of DC in the pathogenesis of autoimmune diseases. Rheumatoid arthritis (RA) is a systemic autoimmune disease characterised by chronic synovitis, which ultimately leads to cartilage destruction and bone erosions. The strong association of RA with certain HLA class II antigens suggests that HLA restricted antigen presentation may be implicated in the disease pathogenesis.12–14 Moreover, the presence of numerous DC in the rheumatoid synovial fluid and membrane suggests a potential role of these cells in RA.15–17 DC which are present at the target organ probably derive from progenitors in the bone marrow and migrate from the peripheral blood to the inflamed joints. Once in the target organ, DC are implicated in the presentation of (auto)antigens to memory T cells or in the activation of naive T cells, or both.
For antigen capture, DC use a wide variety of receptors such as C-type lectins and Fc gamma receptors (FcγR).3,18 The latter are crucial in the recognition and uptake of immune complexes (IC) containing IgG, which are abundantly present in the serum and synovial fluid of patients with RA.19,20 In humans these FcγR can be divided into three classes: FcγRI, FcγRII, and FcγRIII.21–23 The FcγRII are further divided into three subtypes: FcγRIIa, FcγRIIb, and FcγRIIc. FcγRIIa, together with FcγRI and FcγRIII, activate cellular responses upon triggering. In contrast, FcγRIIb are the only FcγR known with the opposite effect.24,25 Both activation and inhibitory FcγR are expressed on DC and the concerted action of these opposing signalling systems determines the cellular response.23–25 Several elegant studies have shown that FcγR are implicated in a wide variety of cellular and disease mechanisms, including cross priming, a unique feature of iDC,26,27 tumour necrosis factor α (TNFα) production by monocytes,28 experimental arthritis,22,29,30 and other autoimmune diseases.22 Therefore, it has been suggested that FcγR constitute the link between humoral and cellular immunity, in which internalisation of IC results in the initiation of T cell immunity and modulation of autoimmune disease such as RA.
In this study we analysed the phenotype of monocyte derived DC from patients with active RA and compared them with DC from patients with RA in remission and healthy volunteers. Among a large panel of membrane markers, the unique difference was a consistently increased expression of FcγRII on iDC and mDC of patients with active RA. Moreover, a clear expression of this receptor was seen in mDC present only in rheumatoid synovial tissue. This provides strong evidence for an important role of DC in the modulation of autoimmunity in RA.
PATIENTS AND METHODS
Patients and samples
A total of 37 patients attending the outpatient clinics of the University Medical Centre, Nijmegen and 32 healthy controls were enrolled in the present study. All patients fulfilled the American College of Rheumatology criteria for RA30 and gave informed consent for the study. Disease activity was assessed with the 28 joint disease activity score (DAS28)31 and, for this study, RA in remission was defined as a DAS28 <3.1, whereas patients with a DAS ⩾3.2 were classified as active RA. Therapeutic regimens of all patients were recorded before blood sampling. Patients receiving prednisone within 6 weeks or biological treatments. including anti-TNFα or interleukin 1 receptor antagonist (IL1Ra), before the study were not included. Immunohistological studies were performed in synovial tissue of three patients with RA and three healthy volunteers. The study was approved by the ethics committee of the University Medical Centre, Nijmegen.
Isolation of monocytes and generation of monocyte derived DC
Venous blood was collected in 10 ml lithium heparin Vacutainer tubes (Becton and Dickinson) and the peripheral blood mononuclear cell (PBMC) fraction was obtained by density gradient centrifugation over Ficoll-Hypaque (Pharmacia Biotech AB). To obtain fresh monocytes, PBMCs were incubated with MACS CD14 labelled microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany). Positively labelled cells were then isolated using separation columns. For the generation of monocyte derived DC, PBMCs were allowed to adhere for 1 hour at 37°C in RPMI-1640, Dutch modification (Life Technologies), enriched with 2% heat inactivated pooled human serum from healthy subjects, in six well culture plates (Costar) after several stringent washing procedures. Adherent monocytes were cultured for 6 days in RPMI-1640, Dutch modification, supplemented with 10% fetal calf serum and antibiotic-antimycotic (all Life Technologies) in the presence of interleukin (IL)4 (500 U/ml; Schering-Plough) and granulocyte monocyte-colony stimulating factor (800 U/ml; Schering-Plough). Fresh complete culture medium was added after 3 days.
At day 6 the iDC were harvested. To generate mDC, iDC were transferred to new six well culture plates and cultured for two more days in the presence of complete culture medium supplemented with 2 μg/ml lipopolysaccharide (LPS; Ecoli; Sigma).
Fluorescence activated sell sorter (FACS) analysis
Expression of cell surface markers on both iDC and mDC was measured by indirect immunofluorescence staining. The first layer was attached after incubation of 1×105 monocyte derived DC for 30 minutes at 4°C with the monoclonal antihuman antibodies against CD14, CD16, CD32, CD64 (all DAKO), DC-SIGN,10 CD80 (Becton Dickinson), CD83 (Coulter), CD86 (Pharmingen), MHC-I (clone W6/32), MHC-II DQ (clone SPV-L3), and MHC-II DP/DR (clone Q1513). After a washing step, the cells were incubated with fluorescein isothiocyanate conjugated goat antimouse IgG for 30 minutes at 4°C in complete darkness. Subsequently, the cells were washed and analysed with a FACS flow cytometer (Becton and Dickinson). Cells were gated according to their forward and side scattering patterns and the expression of DC-SIGN and CD83 for iDC and mDC, respectively. For each marker 104 cells were counted in the gate.
Double immunofluorescence staining techniques were also used for certain studies. Therefore, mDC or monocytes were labelled with CD83-FITC and CD14-FITC, respectively. The second antibody consisted of anti-CD64-PC5 (FcγRI; DAKO), anti-CD32-PE (FcγRII; DAKO), or anti- CD16-PC5 (FcγRIII; Coulter). The cells were incubated for 30 minutes on ice. For these experiments the gate was set over the CD83 (mDC) or CD14 positive (monocytes) cell population.
During all FACS procedures, mouse IgG2b was used as an isotype control and the same instrumental settings were used. In all FACS analyses the amount and position of viable cells was controlled using propidium iodide staining.
FcγR mRNA expression using reverse transcriptase-polymerase chain reaction (RT-PCR)
Expression of FcγRIIa and FcγRIIb mRNA levels was detected by semiquantitative RT-PCR. Total RNA was extracted from PBMCs, iDC, and mDC using 1 ml TRIzol reagent (Life Technologies). RT-PCR was performed using 1 μg RNA. Oligo-dT primers were used for reversed transcription of mRNA and 5% of the resulting cDNA was used in one PCR amplification. The latter was performed at a final concentration of 200 μM dNTPs, 0.1 μM of each primer, and 1 unit Taq polymerase (Life Technologies) in standard PCR buffer. The FcγR specific primers comprised FcγRIIA (forward 5′-ATG TCT CAG AAT GTA TGT CCC AGA, reverse 5′-GTG AAG GCT GCC CAA TTT GAG) and FcγRIIB (forward 5′-GGA ATC CTG TCA TTC TTA CCT GTC and reverse 5′-GAT GAG GCT GAC AAA GTT GGG), respectively.32 All experiments were repeated twice and the PCR reaction was paused after 15, 18, 21, 24, 27, 30, 33, and 35 cycles at the end of the extension phase (72°C) to obtain 5 μl samples. The latter underwent electrophoresis on 1.6% agarose gels and were stained with ethidium bromide. The results are expressed as the number of cycles needed to detect FcγRIIa or FcγRIIb compared with the number needed to detect the housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
Immunohistochemical co-expression of DC-LAMP and FcγRII in synovial tissue
Percutaneous synovial tissue biopsy specimens of RA knee joints and synovial tissue from controls obtained during arthroscopic procedures were embedded in Tissue Tek OCT (Miles Inc) and snap frozen in liquid nitrogen. Cryostat sections (7 μM) were mounted on superfrost slides and stored at −70°C until processing. Immunohistochemical localisation of CD32 (FcγRII; DAKO) and DC-specific lysosome associated membrane glycoprotein (DC-LAMP), a specific marker for mDC, was performed using serial sections. Briefly, cryostat sections were dried, fixed in 100% cold acetone (10 minutes), and dried again. Thereafter, endogenous peroxidase was blocked using H2O2 in phosphate buffered saline and incubated with the first antibody (5 μg/ml, 60 minutes at room temperature). Subsequently, sections were incubated with normal horse serum and with biotinylated horse antimurine IgG (Vectorlabs) and ABC complexes (Vectastain). Slides were developed with diaminobenzidine and counterstained with haematoxylin for 3 minutes. Controls consisted of irrelevant primary isotype-specific IgG1 and IgG2a (DAKO) and omission of the secondary antibodies.
TNFα production by DC after stimulation with IC
In certain experiments DC were cultured in the presence of heat aggregated gamma immunoglobulins (HAGGs) which bind and crosslink FcγR. HAGGs were obtained by heating human IgG (Sigma Chemicals) at 63°C for 30 minutes. Thereafter the solution was centrifuged at 12 000 g for 10 minutes and the concentration of HAGGs was determined from the absorbance at 280 nm. HAGGs were then added to the culture medium (final concentration 100 μg/ml) of iDC or, during the maturation, to mDC. TNFα production from iDC or mDC before or after stimulation with HAGGs was measured in the supernatants with enzyme linked immunosorbent assay (ELISA) techniques (detection limit 2 pg/ml) using an experimental set up previously described for components of the plasminogen activator system.33
Statistics
A Mann-Whitney U test and Student’s t test were used as appropriate. Values of p were two sided and the level of significance was set at p<0.05.
RESULTS
Increased FcγRII expression on iDC and mDC from patients with RA
Table 1 shows the results of the FACS analysis of a panel of membrane markers on iDC and mDC. For the majority of markers, no differences were seen between the two groups for either the number of expressing cells (%) or the mean fluorescence intensity (MFI). In both healthy controls and patients with RA, DC maturation was reflected by the down regulation of the monocyte differentiation marker CD14, whereas the expression of costimulatory molecules (CD80 and CD86), CD83 (mDC marker), and MHC molecules was clearly increased. Interestingly, a clear difference in expression of FcγR was seen (fig 1). Comparison of the two groups showed a significantly increased FcγRII (CD32) expression (MFI) on iDC from patients with RA (RA DC) (n = 30) when compared with that of controls (C DC) (n = 31) (454 (94) v 162 (36) (mean (SD)), p = 0.001) (fig 1B). Expression of the FcγRI (CD64; 31 (12) v 33 (11), p = 0.46) and FcγRIII (CD16; 32 (35) v 43 (58), p = 0.38) was similar.
FACS analysis of a panel of membrane markers on iDC and mDC

FcγRI, II, and III expression on iDC and mDC from patients with RA and healthy subjects. (A) FACS analysis of the indicated markers (solid line) or isotype controls (dotted line) of DC within the life gate. Numbers within the histograms represent the mean fluorescence of the marker corrected for isotype values. Each graph displays data from one representative donor. (B) Averaged mean expression of the indicated markers from the whole group of healthy donors (n = 32) and patients with RA (n = 31) of both iDC and mDC.
Subsequent analysis of FcγR expression on mDC resulted in an even more intriguing observation. Expression of FcγRI (p = 0.03), FcγRII (p = 0.008), and FcγRIII (p = 0.003) was clearly down regulated upon maturation in DC from healthy subjects. Mature DC of patients with RA, however, showed a distinct decrease in FcγRI (p = 0.002) and FcγRIII (p = 0.004) expression but much less for FcγRII (p = 0.07). This resulted in an almost eightfold higher expression of FcγRII on RA mDC than control mDC (256 (40) v 34 (10), p = 0.0002) (fig 1B, right panel). To confirm this observation, we performed double staining procedures with mDC from a smaller group of patients (n = 13) and healthy controls (n = 9) using CD83 as a marker for mDC together with the three FcγR. This confirmed that mDC from patients with RA expressed FcγRII at higher levels than those from healthy subjects (500 (150) v 188 (97), p = 0.005), whereas this was not the case for FcγR classes I and III (fig 2B, left panel).

Expression of FcγRI, II, and III on mDC and monocytes from patients with RA and healthy subjects using double staining techniques. (A) MFI of the FcγR subtypes (solid line) and isotype control (dotted line) on mDC (CD83, FcγR double positive cells) and monocytes (CD14, FcγR double positive cells) within the life gate by using double staining FACS analysis. The mean fluorescence is indicated within the histogram. (B) Averaged mean expression of the indicated markers on mDC and monocytes from the whole group of healthy donors (n = 9, n = 10) and patients with RA (n = 13, n = 10), respectively. Of note, only CD83, FcγR and CD14, FcγR double positive cells are shown.
Because the DC were generated from monocytes, we investigated whether the increased FcγRII expression was specific for DC or merely a result of a pre-existing up regulation of this receptor on precursors of DC. Therefore freshly isolated PBMCs from 10 patients and 10 healthy donors were studied using double staining for CD14 as the monocyte marker and FcγRI, II, and III. A slightly higher frequency of CD16 positive monocytes was detected in patients with RA (11.1 (4) v 4.4 (2), p = 0.046) as found previously,34 but the mean expression level of FcγRI (133 (47) v 92 (21), p = 0.09), FcγRII (582 (150) v 461 (140), p = 0.08), and FcγRIII (178 (128) v 165 (63), p = 0.3) on cells double positive for CD14 and the corresponding FcγR was not different than that of controls (figs 2A and B). This suggests an aberrant regulation of FcγRII expression during the transition from monocytes to DC.
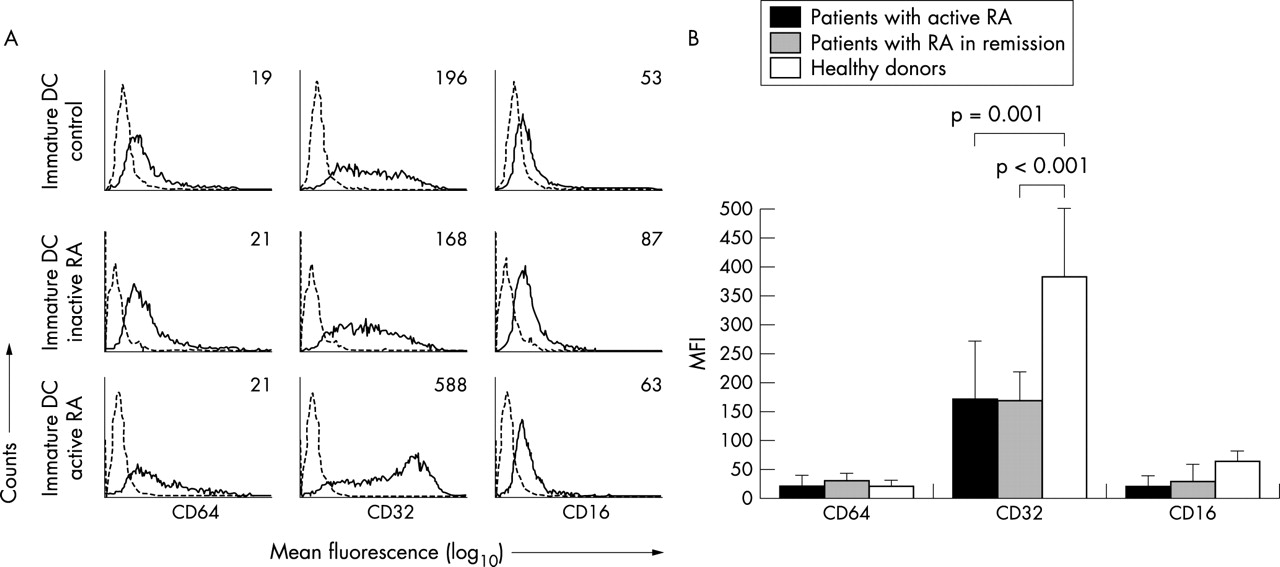
We then studied the influence of disease activity on the expression of FcγR by DC. The latter was measured in a new group of subjects comprising eight patients with active RA, six patients with RA in remission, and 10 healthy controls. Only patients with active disease showed an increased expression of FcγRII when compared with patients in remission (382 (120) v 170 (50), p<0.001) and healthy controls (382 (120) v 172 (100), p<0.001) (fig 3).

FcγRI, II, and III expression by iDC and influence of RA disease activity. (A) FACS analysis of FcγR subtypes (solid line) and isotype control (dotted line) on iDC from a healthy donor, a patient with active RA, and a patient with RA in remission. The numbers within the histograms indicate the mean fluorescence. Each histogram displays one representative person. (B) Averaged mean expression of CD64, CD32, and CD16 (FcγRI, II, and III, respectively) on iDC from healthy donors (n = 10), patients with RA in remission (n = 6), and patients with RA with active disease (n = 8).
FcγR mRNA expression
To determine the relative expression of both FcγRIIa and FcγRIIb, semiquantitative RT-PCR techniques were performed. Therefore, we compared the expression levels of FcγRIIa and FcγRIIb from eight patients with active and six with inactive disease and 10 healthy controls. Figure 4 shows that iDC from patients with active RA expressed the FcγRIIa at comparable levels to patients with inactive RA and healthy controls. In contrast, the median mRNA expression of FcγRIIb was significantly higher in iDC (p<0.001) and mDC (p<0.004) from patients with active disease. The mRNA expression levels of the FcγRII subtypes in DC from patients with inactive RA were not significantly different from those of healthy controls and were clearly down regulated during DC maturation (fig 4).

mRNA expression of FcγRIIa and FcγRIIb by iDC and mDC from patients with RA and healthy controls. The bars represent the median level of FcγRII mRNA analysed with PCR techniques, corrected for the expression of the housekeeping gene GAPDH. Eight patients with active RA, six patients with RA in remission (n = 6), and 10 healthy donors (n = 10) were studied.
Opposing effects on TNFα production after addition of IC to DC from patients with RA and controls
We showed that both the FcγRII protein and FcγRIIb mRNA expression were increased on DC from patients with RA. To investigate whether this increased FcγRII expression has functional consequences for the behaviour of these DC we tested their response to stimulation through the FcγR dependent pathways. As a measure of FcγR activation, we determined the TNFα production of both iDC and mDC from patients with RA and controls with and without stimulation with HAGGs.
Figure 5A shows that the basal TNFα production by iDC was low and similar for patients with RA (n = 21) and healthy controls (n = 18) (28 (0–470) v 35 (0–270) (median (range)). However, the production of TNFα was strongly increased after maturation of DC in both patients with RA (p<0.001) and healthy controls (p<0.001) and was significantly higher in RA (n = 21) than in controls (n = 18) (2400 (1500–9900) v 352 (0–1700), p<0.0001). As expected, addition of IC to mDC increased production of TNFα in controls (n = 8; 278 (40–1500) v 1500 (345–3500), p = 0.007) (fig 5C). In contrast, the presence of IC had the opposite effect on DC from patients with RA (n = 10; 3500 (1541–6500) v 1825 (580–3250), p = 0.01), (fig 5B).

(A) TNFα production by iDC and mDC. TNFα production (pg/ml) by iDC (n = 21) and mDC (n = 21) from patients with RA (RA DC) and DC from healthy controls (n = 18) (C DC). Full maturation was achieved by adding LPS on day 6. (B) TNFα production by mDC after stimulation with anti-IgG complexes (HAGGs). TNFα production of mDC with and without HAGGs stimulation from patients with RA (n = 10, B) and from healthy subjects (n = 8, C). Full maturation was achieved by adding LPS on day 6.
Mature DC expressing FcγRII are present in RA synovial tissue
To analyse whether the differences between monocyte derived DC from patients with active RA and healthy controls are also found in vivo we studied the presence of mDC with FcγRII expression in the target organ, the synovial tissue. For this, consecutive sections of synovium obtained from three patients with RA and three healthy controls were stained with FcγRII and DC-LAMP, the mDC marker. Figures 6A and B show overlapping expression patterns of DC-LAMP and FcγRII in rheumatoid synovium, suggesting that the mDC that are scattered throughout the perivascular regions also express high levels of FcγRII. The expression of FcγRII was as expected, not limited to DC-LAMP positive cells but also present in other inflammatory cells in the lining, sublining, and deeper regions. In contrast, the synovial tissue from healthy donors showed hardly any FcγRII staining and a complete absence of DC-LAMP positive cells (figs 6C and D).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Co-expression of FcγRII and CD83 in synovial tissue. (A) and (B) show immunostaining of RA synovial tissue with CD32 and DC-LAMP, respectively. Immunostaining with the same set of markers is shown for synovial tissue of a healthy control (HC), respectively.
DISCUSSION
This study provides evidence for an increased expression of FcγRII on DC in patients with active RA. Our results show that among a wide panel of DC markers, this was the only phenotypic difference between RA and control DC. The highly increased expression of FcγRII in RA was seen not only on iDC but also on fully matured DC. This lack of FcγRII down regulation on mature RA DC is in contrast with the observed FcγRII down regulation on mDC from healthy subjects. The medication used did not differ between patients with active and inactive disease and could thus not explain this phenomenon. Furthermore, mDC are abundantly present in the perivascular infiltrates of the synovial tissue of patients with RA, whereas they are not present in synovial tissue from healthy donors. Intriguingly, overlapping expression patterns of FcγRII and DC-LAMP exist in RA but not in healthy synovial tissue, suggesting that mDC do express FcγRII in RA synovium in vivo. Finally, our results indicate that overexpression of FcγRIIb has functional implications for DC, affecting the production of proinflammatory cytokines.
DC are professional antigen presenting cells and there is increasing evidence that DC have a substantial role in the pathogenesis of RA.9,15,17,35,36 It is now well appreciated that DC have specific functions according to their state of maturation, leading to either immunity or tolerance.37,38 Immature DC mainly reside in the periphery and are highly specialised in antigen uptake, endocytosis, and processing, whereas mDC normally lose this ability. mDC, however, are fully equipped for optimal antigen presentation and migrate to the draining lymph nodes, where they activate T cells. The presence of mDC in RA synovium and their preferential location in the perivascular infiltrates, where the interaction with T and B cells takes place, is therefore intriguing, but in line with previous studies.15,17,35 We now demonstrate that these synovial mDC are unique in that they still express high levels of the FcγRII.
FcγR play an important part in antigen uptake and cross presentation, which are key processes and unique features of DC.24,25 DC use the FcγRII for uptake of antigens and IgG immune complexes (IgG IC), which are abundantly present in several RA compartments such as peripheral blood, synovial fluid, and tissue. Two FcγRII subtypes exist, the stimulatory receptor FcγRIIa and the inhibitory receptor FcγRIIb. Factors such as IL4 and interferon gamma determine the balance between these FcγRII subtypes, as recently demonstrated by Pricop et al.39 An altered balance of these FcγRII subtypes during disease could result in altered behaviour of DC and macrophages. Previously, we demonstrated that macrophages from patients with RA express higher levels of FcγRII and FcγRIII and produce increased levels of TNFα and matrix metalloproteinase relative to controls upon addition of IC.34
Our data now indicate that both FcγRII mRNA subtypes are also increased in DC derived from patients with active RA, and show that the FcγRII balance is shifted towards the inhibitory FcγRII subtype (FcγRIIb). This observation is supported by the finding that the addition of IC decreases TNFα production by DC from patients with RA, whereas a decrease was found after FcγR dependent stimulation of DC from healthy subjects. This, together with the PCR data implies that the higher FcγRII expression is also functionally balanced towards the inhibitory FcγRII subtype in patients with RA. However, we also observed that mDC derived from patients with RA produce significantly larger amounts of TNFα than mDC from healthy subjects. TNFα is a key proinflammatory cytokine in RA and the therapeutic effect of TNFα blockade in this and other inflammatory diseases has been clearly demonstrated.40,41
The observations in our study may appear contradictory and raises a question about how DC are entangled in the autoimmune phenomena seen in RA. One possible clue comes from the finding that the increased FcγRII expression is restricted to the active phase of RA. Possibly, this might reflect a protective mechanism in patients with active RA to counteract harmful inflammation. The balance towards the inhibitory FcγRII subtype during active RA might be the result of high levels of IL13 present in RA synovial fluid.42,43 IL13 shares many functional properties of IL4 and might possibly, like IL4, induce the inhibitory FcγRIIb. The importance of inhibitory FcγR in the clearance of IC and the inflammatory cascade has already been elegantly shown in several in vivo and in vitro studies.44–46 In patients with inactive RA, the DC might be involved in the transition to active RA by releasing large amounts of TNFα after activation. The release of TNFα would subsequently trigger the inflammatory cascade leading to the production of cytokines such as IL13, which in turn leads to increased levels of the inhibitory FcγRII. Such a dual role of DC in RA might also explain the changing pattern of active and inactive disease, which is characteristic for patients with RA.
In summary, we demonstrate that DC from patients with active RA have an increased expression of FcγRII. The inhibiting FcγRIIb, especially, was increased, and addition of IC altered the response of the DC. Additional studies are required to elucidate the local factors which play a part in the regulation of the balance between inhibitory and activating FcγR in RA.
Acknowledgments
We are indebted to all those who participated in the collection of peripheral blood specimens. We thank Dr M de Waalmalefijt and Dr W Schreurs from the department of orthopaedic surgery for their participation in the collection of synovial tissue.