Article Text

Abstract
Objective Innate lymphoid cells-2 (ILC2) were shown to be involved in the development of lung or hepatic fibrosis. We sought to explore the functional and phenotypic heterogeneity of ILC2 in skin fibrosis within systemic sclerosis (SSc).
Methods Blood samples and skin biopsies from healthy donor or patients with SSc were analysed by immunostaining techniques. The fibrotic role of sorted ILC2 was studied in vitro on dermal fibroblast and further explored by transcriptomic approach. Finally, the efficacy of a new treatment against fibrosis was assessed with a mouse model of SSc.
Results We found that ILC2 numbers were increased in the skin of patients with SSc and correlated with the extent of skin fibrosis. In SSc skin, KLRG1− ILC2 (natural ILC2) were dominating over KLRG1+ ILC2 (inflammatory ILC2). The cytokine transforming growth factor-β (TGFβ), whose activity is increased in SSc, favoured the expansion of KLRG1- ILC2 simultaneously decreasing their production of interleukin 10 (IL10), which regulates negatively collagen production by dermal fibroblasts. TGFβ-stimulated ILC2 also increased myofibroblast differentiation. Thus, human KLRG1- ILC2 had an enhanced profibrotic activity. In a mouse model of SSc, therapeutic intervention-combining pirfenidone with the administration of IL10 was required to reduce the numbers of skin infiltrating ILC2, enhancing their expression of KLRG1 and strongly alleviating skin fibrosis.
Conclusion Our results demonstrate a novel role for natural ILC2 and highlight their inter-relationships with TGFβ and IL10 in the development of skin fibrosis, thereby opening up new therapeutic approaches in SSc.
- systemic sclerosis
- fibroblasts
- inflammation
Data availability statement
Data are available on reasonable request.
This is an open access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited, appropriate credit is given, any changes made indicated, and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/.
Statistics from Altmetric.com
Key messages
What is already known about this subject?
Type 2 innate lymphoid cells (ILC2) have emerged as a player in inflammatory and fibrotic processes.
Research to date on innate lymphoid cells in systemic sclerosis (SSc) was descriptive suggesting a potential role in the disease development.
Transforming growth factor-β (TGFβ) pathway is important in SSc pathophysiology notably through its direct role on fibroblasts.
What does this study add?
We deeply described ILC2 presence and localisation in fibrotic skin.
We depicted a new indirect mechanism by which TGFβ could lead to fibrosis, triggering the switch from an ‘inflammatory’ phenotype (KLRG1high) to a ‘natural’ phenotype (KLRG1low) ILC2.
These TGFβ-activated ILC2, characterised by a diminished interleukin 10 (IL10) production, promote collagen synthesis by fibroblasts.
Using both in vitro and in vivo models, we established the importance of the combined role of TGFβ and IL10 in the fibrotic process.
How might this impact on clinical practice or future developments?
These data provide important support for the use of combination therapies in SSc.
The combined use of an antifibrotic drug such as pirfenidone and IL10 could be a new therapeutic approach in this very complicated disease.
Introduction
Systemic sclerosis (SSc) is a systemic autoimmune disorder characterised by a dysregulated extensive fibrotic process that impacts epithelial barriers, within the gut, lung and skin.1 Its pathogenesis remains poorly understood, and treatments for disease progression are limited. While the adaptive immune system has long been considered to be involved in SSc development, recent observations have established an important role of the innate immune system.2 3 As an example, a type 2 macrophage signature has been identified in both skin and lung from patients with SSc.4
Interestingly, innate lymphoid cells (ILCs) that patrol environmental interfaces to defend against infection and protect barrier integrity have emerged as crucial effectors in inflammatory and fibrotic diseases.5 6 Their cytokine production and transcription factor expression allow the identification of three distinct subsets. Type 1 ILC (ILC1) are Tbet expressing cells that produce interferon-γ and tumour necrosis factor, and are dependent on IL12 and IL18 for their generation. GATA3 expressing type 2 ILC (ILC2), which are dependent on thymic stromal lymphopoietin (TSLP), IL25 and IL33 release IL5 and IL13 whereas type 3 ILCs (ILC3) express the transcription factor RORγt, release IL17 and IL22 and are generated after IL23 and IL1β stimulation.7
The role of ILC2 was first highlighted in allergic reactions.8 9 More recently, murine studies have shown the role of ILCs in hepatic and pulmonary fibrosis, with a major effect of IL13 production. Interestingly in the context of SSc, TGFβ is another important profibrotic factor10–12 and an essential cytokine for the development of ILC2,13 suggesting a potential role of ILC2 on SSc pathogenesis.
To add another level of complexity, ILC2 constitute a heterogeneous population of cells and at least two separate clusters are described based on their differential responses to microenvironment. Inflammatory ILC2 (iILC2) respond to IL25 and produce IL17 in addition to IL13, whereas natural ILC2 (nILC2) respond to IL33 and release high levels of IL13. The differential expression of killer cell lectin-like receptor G1 (KLRG1) has been identified as a marker, with iILC2 being KLRG1+ and nILC2 being KLRG1-. iILC2 can be considered a transient progenitor based on its ability to migrate to tissue and then differentiating into nILC2 in response to activation signals.14 15 However, the relevance of this plasticity in human pathologies remains to be established.
In patients with SSc, we and others have demonstrated increased levels of homeostatic cytokines for ILC2, such as IL25, IL33 and TSLP.16–18 Furthermore, our group found that TSLP is increased in the blood and skin of patients with SSc, with levels correlating to skin fibrosis.18 In human SSc, the role of ILC2 remains elusive as only one observational study showed an increased proportion of circulating ILC2 at the blood and tissue level,19 thus questioning the potential implication of ILC2 in the fibrotic process.
In our study, we demonstrate the potential role of ILC2 in the establishment of fibrosis in human SSc. We showed that KLRG1 expression on ILC2 was linked to the fibrotic stage of the disease. Mechanistically, in vitro and in vivo studies revealed that this switch operates in a TGFβ-dependent manner, leading to a decrease in IL10 production and a profibrotic phenotype. Interestingly, while pirfenidone alone (acting partly by the inhibition of TGFβ-induced effects) failed to significantly affect the fibrotic process in vivo, the addition of IL10 synergistically alleviated fibrosis. Overall, this study unravels a new role for ILC2 in fibrotic diseases and paves the way for new therapeutic strategies for human SSc.
Results
Number of circulating ILC2 is decreased in human SSc with the extent of skin fibrosis
To investigate the potential contribution of ILC2 in SSc pathogenesis, we first monitored the total ILC population and subpopulations in the whole blood of patients with SSc (SSc, n=73) and age-matched and sex-matched healthy donors (HDs) (n=59) (table 1).
Demographic, clinical and biological characteristics of the SSc population
The gating strategy to identify ILCs in the peripheral blood is described in the online supplemental figure 1A. The frequency and absolute numbers of ILCs (defined as Lin-CD45+CD127+, figure 1A) were lower in SSc compared with HDs (0.04%±0.02% vs 0.09%±0.07%, p<0.0001 and 0.0009±0.0003 vs 0.002±0.001, p=0.0004; figure 1B and online supplemental figure 1AC, respectively), with ILC1, ILC2 and ILC3 being 74%, 15% and 11% in SSc and 55%, 18% and 27% in HDs, respectively (figure 1C). When focusing on ILC2 defined as Lin-CD45+CD127+CRTH2+, their frequency and absolute numbers were approximately three times lower in SSc compared with HDs (figure 1D and online supplemental figure 1BD, respectively). However, no differences in the frequency of KLRG1+ ILC2 were found (online supplemental figure 1CE). Decreased were also the frequency and absolute numbers of ILC3, but not of ILC1 when SSc and HDs were compared (figure 1D and online supplemental figure 1BD).
Supplemental material

Characterisation of innate lymphoid cells (ILCs) in the blood of patients with systemic sclerosis (SSc) and healthy donors (HDs). (A) Representative dot plot of circulating ILCs in the HD and SSc blood samples and (B) ILC frequency quantification. (C) Proportion of ILC subsets in the blood from HDs and patients with SSc. (D) Percentage of circulating ILC1, ILC2 and ILC3 in the HD and SSc blood. (E) Correlations between circulating ILC1, ILC2 and ILC3 with the extent of cutaneous fibrosis (modified Rodnan skin thickness score (mRSS)). Data are the mean±SEM (n=59 and 73 for HDs and patients with SSc, respectively). Comparisons between groups were calculated using Mann-Whitney U test. **P<0.01; ****p<0.0001.
To evaluate the clinical relevance of these observations, we analysed the ILC numbers to different relevant clinical parameters. We specifically observed a correlation between the quantity of circulating ILC2 and the modified Rodnan skin thickness score (mRSS), showing that lower is the amount of circulating ILC2 and higher is the cutaneous fibrosis (figure 1E and online supplemental figure 1DF). Of note, the ILC1 and ILC3 frequencies were not correlated with the mRSS. When comparing patients with or without interstitial lung disease, we found no difference on the % and the absolute count of ILC2 in the whole blood and there were also no differences according to the severity of interstitial lung disease or the disease duration (data not shown). Collectively, our data indicate that patients with SSc are characterised by a significant reduction in the proportion and number of circulating ILC2, which is correlated with the extent of skin fibrosis.
ILC2 are increased in human SSc skin and correlated with the extent of fibrosis
Since circulating ILCs were decreased, we further characterise ILCs infiltration in the skin. We first extracted cells from the skin and performed flow cytometry analysis. Representative staining depicting the gating strategy for ILCs and subpopulation categorisation is shown in figure 2A and online supplemental figure 2A. As suspected, we found that the percentage of total ILCs among CD45+ cells was increased in the SSc skin compared with that in the HD skin (figure 2B). When evaluating the repartition of ILC subsets among the total skin ILCs, we observed that 69% were ILC2 in the HDs vs 77% in the patients with SSc, 27% were ILC1 in the HDs vs 19% in the patients with SSc (figure 2C), while ILC3 were barely detectable. When analysed among CD45+ cells, a significant increase in ILC2 was observed in the SSc skin compared with the HD skin (figure 2D). The ILC1 frequency was similar in the patients with SSc and HDs (online supplemental figure 2AB), even if the proportion among ILCs was decreased in the patients with SSc compared with that in the HDs (figure 2C). The increased percentage of ILC2s among CD45+ skin cells was correlated with the extent of skin fibrosis (figure 2E), whereas the ILC1 percentage did not show this correlation (online supplemental figure 2BC).
Supplemental material

Characterisation of cutaneous innate lymphoid cells (ILC)-2 (ILC2) in patients with systemic sclerosis (SSc) and healthy donors (HDs). (A) Representative dot plot of cutaneous ILCs in the HD and SSc skin samples and (B) ILC frequency quantification. (C) Proportion of ILC subsets in the skin of HDs and patients with SSc. (D) Percentage of ILC2 among CD45+ lymphoid cells in the HD and SSc skin samples. (E) Positive correlation between the percentage of ILC2 among CD45+ cells and the extent of cutaneous fibrosis (modified Rodnan skin thickness score (mRSS)). (F) Number of ILC2 per mm2 in the dermis of HD and SSc skin. (G) Correlation between the extent of cutaneous fibrosis (mRSS) and the number of ILC2 per mm2 in the dermis. (H and I) Representative picture of an immunofluorescence assay using anti-CRTH2-based immunofluorescence (green) and antilineage (CD3, CD11b and FcεR1)-based immunofluorescence (red) performed to detect CRTH2+Lin- ILC2 in the HD and SSc skin samples (scale bars=100 µm). Bar graphs show data as the mean±SEM (n=18–20 and 17–32 for HD and SSc, respectively). Comparisons between groups were calculated using Mann-Whitney U test. *P<0.05; **p<0.01.
To validate this analysis and gain more insights on the precise localisation and quantification of ILC2 in the skin, we performed tissue immunofluorescence staining. Representative staining of ILC2 (Lin-DAPI+CRTH2+) for an HD and an SSc patient is depicted in figure 2H,I. An analysis of the ILC2 distribution and quantification revealed that the number of ILC2 per surface area (mm2) and the percentage of ILC2 (per total cell count) were increased in the patients with SSc compared with the HDs in the dermis (figure 2F and online supplemental figure 2CD). Of interest, the number of ILC2 per mm2 in the dermis was positively correlated with the extent of skin fibrosis as assessed by mRSS at the time of biopsy (figure 2G).
Overall, these results show that ILC2 are increased in human SSc skin and their frequency in the dermis is associated with skin fibrosis.
KLRG1 expression on skin ILC2 is modified over the course of the disease
To determine the phenotype of cutaneous ILC2 in the SSc skin, we studied the expression of several markers: HLA DR, OX40L, CCR10, CCR6, CLA, TSLPR and KLRG1 (figure 3A,B). The percentage of ILC2 expressing HLA-DR, OX40L, CCR6, CCR10, CLA and TSLPR was similar in the skin of the patients with SSc and HDs. In sharp contrast, the percentage of KLRG1-positive cells was significantly lower in the SSc skin (figure 3A,B). Interestingly, the percentage of ILC2 KLRG1+ tends to decrease with the extent of cutaneous fibrosis (figure 3C).
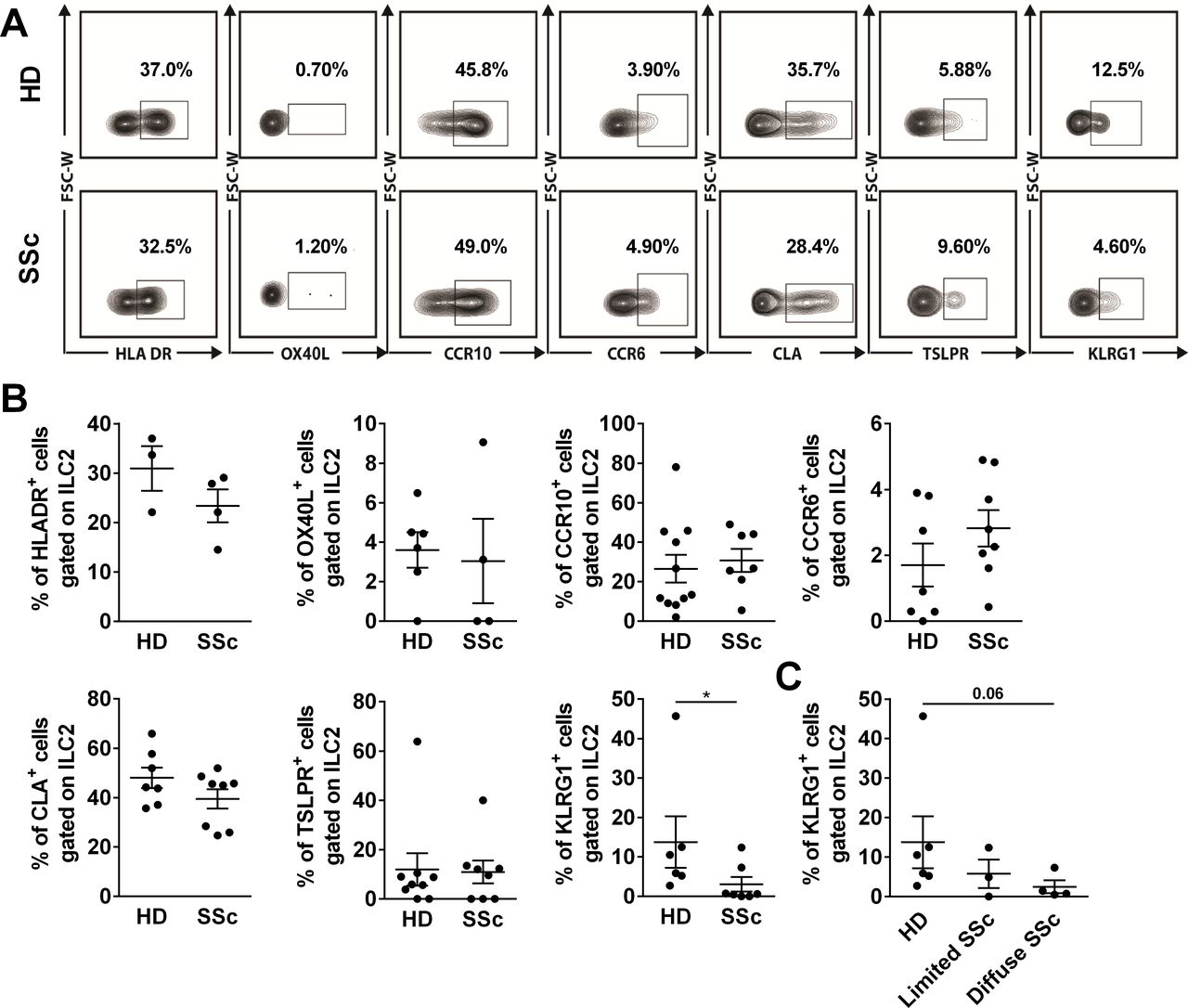
Decrease of killer cell lectin-like receptor G1 (KLRG1) expression on innate lymphoid cells (ILC)-2 (ILC2) in the skin of patients with systemic sclerosis (SSc). (A and B) Expression of HLA DR, OX40L, CCR10, CCR6, CLA, TSLPR and KLRG1 on skin ILC2 in the healthy donors (HDs) and patients with SSc. (C) Percentage of KLRG1+ cells among ILC2s in patients with SSc with limited (Rodnan score <10) and diffuse (Rodnan score >10) in patients with SSc. Bar graphs show data as the mean±SEM (n=3–11 and 4–8 for HD and SSc, respectively). Comparisons between groups were calculated using Mann-Whitney U test. *P<0.05.
Altogether, these results indicated that surface KLRG1 expression on ILC2 decreases in the SSc skin and may correlate with the extent of skin fibrosis thus raising the question of the functional relevance of this observation.
TGFβ promotes in vitro KLRG1 modulation of ILC2 and impacts IL10 secretion
The relevance of KLRG1 in ILC2 modulation are uncertain and remains to be proven, particularly in human SSc. To identify the factors implicated in KLRG1 modulation, we sorted ILC2 from the peripheral blood of HDs (online supplemental figure 3A) and expanded this population in vitro in the presence of IL1β and IL2 as previously described.20 At the end of the culture, ILC2 were consistently CD127+CRTH2+ (online supplemental figure 3B) and characterised by high expression of GATA3 (online supplemental figure 3C).21 We then tested cytokines, specifically involved in human SSc, for their ability to modulate KLRG1 expression on expanded ILC2. IL33, IL4, TSLP and IL25 did not modify KLRG1 expression while TGFβ dramatically decreased the expression of KLRG1 on ILC2 (figure 4A,B).
Supplemental material

Transforming growth factor-β (TGFβ) affects the innate lymphoid cells (ILC)-2 (ILC2) phenotype by decreasing killer cell lectin-like receptor G1 (KLRG1) expression and interleukin (IL)10 production. (A) Representative dot blot of KLRG1 expression and (B) percentage of KLRG1+ cells in unstimulated ILC2 (medium) and ILC2 stimulated with IL33, TGFβ, IL4, thymic stromal lymphopoietin (TSLP) and IL25 after 20 days of amplification. (C) Secretion of IL5, IL9, IL13 and IL10 assessed by a cytokine bead assay (CBA) in both types of ILC2 supernatants. Bar graphs show data as the mean±SEM (n=5–16 for KLRG1 expression and n=4–7 for CBA). Comparisons between groups were calculated using Kruskal-Wallis or Wilcoxon tests. *P<0.05.
We then evaluated the functional consequences of TGFβ exposure on ILC2 by analysing its impact on cytokine production. The levels of IL5, IL9 and IL13 were similar in ILC2 exposed or not to TGFβ (figure 4C). In contrast, the production of IL10 in TGFβ-primed ILC2 was significantly reduced compared with that of controls.
Overall, these data indicate that TGFβ downregulates KLRG1 expression on ILC2, whose phenotype is associated with a decreased capability to produce IL10.
IL10 decreased production by TGFβ-stimulated ILC2 leads to a profibrotic profile by fibroblasts
Our observations suggest that the KLRG1- ILC2 population could be involved in the fibrotic process. We therefore investigated the ability of TGFβ-stimulated ILC2 to modify the fibrotic response. To this end, supernatants (SN) from TGFβ-stimulated or unstimulated ILC2 were added to dermal fibroblasts. After 1 day of co-culture, the expression of type I collagen (COL1A1) and matrix metalloproteinase-1 (MMP-1) was evaluated. As expected, TGFβ alone increased COL1A1 and decreased MMP1 messenger RNA (mRNA) expression (figure 5A). TGFβ neutralisation completely blocked these changes. Interestingly, SN from TGFβ-stimulated ILC2 increased the COL1A1 mRNA expression while the SN from unstimulated ILC2 did not affect the COL1A1 mRNA levels. Importantly, TGFβ neutralisation in TGFβ-stimulated ILC2 did not affect these results, thereby ruling out the effect of the initial exogenously added TGFβ on the induction of COL1A1 mRNA in fibroblasts. Furthermore, ILC2 SN dramatically increased the MMP1 mRNA expression independently of the priming conditions (figure 5A). The profibrotic activity of TGFβ-stimulated ILC2 was emphasised by the COL1A1/MMP1 ratio as a surrogate of collagen turnover, which was increased, suggesting an enhanced collagen deposition over degradation (figure 5B). While no difference of expression was observed for COL1A2, the SN of TGFβ-stimulated ILC2 also increased the mRNA of fibronectin (figure 5C).

Transforming growth factor-β (TGFβ) exerts an indirect fibrotic role through innate lymphoid cells (ILC)-2 (ILC2) and interleukin (IL)10 secretion. (A) Q-RT-PCR analysis of type I collagen (COL1A1) and matrix metalloproteinase-1 (MMP-1) messenger RNA (mRNA) expression and (B) COL1A1/MMP1 ratio, in fibroblasts from healthy donors (HDs), incubated with TGFβ or supernatant of unstimulated ILC2 or supernatant of TGFβ-stimulated ILC2 with or without blocking antibodies against TGFβ. (C) Q-RT-PCR analysis of COL1A2 and FN1 mRNA expression. (D) Q-RT-PCR analysis of COL1A1 expression in fibroblasts incubated with supernatant of unstimulated ILC2 (in the presence or absence of anti-IL10 antibodies) and supernatant of TGFβ-stimulated ILC2 (with or without IL10). (E) Human pro-collagen I α1 concentration were analysed in the supernatants of fibroblasts from HDs after 48 hours of activation with ILC2 supernatants. (F) Average growth rate of fibroblasts were analysed after 48 hours of proliferation. (G) The percentage of α-smooth muscle actin positive fibroblasts were analysed after 72 hours of activation. Bar graphs show data as the mean±SEM (n=7–15). Comparisons between groups were calculated using the paired Wilcoxon (#) or the Kruskal-Wallis tests (*). *P<0.05; **p<0.01.
Since the production of IL10 was specifically reduced in TGFβ-stimulated ILC2, we next evaluated the role of IL10 on their profibrotic activity. Of interest, when IL10 was neutralised in control ILC2 SN we observed enhanced COL1A1 mRNA expression of dermal fibroblast (figure 5D). Conversely, the addition of IL10 to TGFβ-stimulated ILC2 SN led to a dramatic decrease in COL1A1 mRNA expression. We then confirmed these results at the protein level (figure 5E). To further explore the fibroblast activation, we measured the proliferation and the differentiation into myofibroblasts. While no difference was observed regarding the proliferation (figure 5F), we found a significant increase expression of α-smooth muscle actin when fibroblasts were incubated with the SN of TGFβ-stimulated ILC2, independently of IL10 (figure 5G). As previously, we paid attention to incubate the SN of TGFβ-stimulated ILC2 with anti-TGFβ blocking antibody, excluding the possibility that residual TGFβ from ILC2 activation may have mediated myofibroblast differentiation.
Collectively, our data show that TGFβ favours the generation of KLRG1- ILC2 characterised by low IL10 production capacity, which simultaneously results in their enhanced profibrotic capacity.
Transcriptomic analysis reveals additional fibrotic potential of TGFβ-stimulated ILC2
To look further on the fibrotic potential of human ILC2, we evaluated the transcriptome of TGFβ-stimulated ILC2 compared with unstimulated ILC2 by RNAseq (online supplemental figure 4A). The heat map shows modification in gene expression of ILC2 triggered by TGFβ activation, with 2840 genes being differentially expressed (p-adjusted value <0.01). The enrichment analysis with R library Gprofiler2 (V.0.2.0), using Gene Ontology databases, indicated SMADs activation (‘heteromeric SMAD protein complex’; GO:0071144; p adjusted value=4.01E-2 and ‘SMAD protein complex’; GO:0071141; p adjusted value=4.45E-2) and immune regulation (online supplemental table E3, E4, E5, E6 and E7). In the first 60 most upregulated genes (online supplemental table E8), we look at genes already described in fibrosis mechanisms. Interestingly, we observed an upregulation of LTC4S (leukotriene C4 synthase) (log2 fold change=2.71; p adjusted value=9.89E-22). Regarding downregulated genes, we found a decrease of IL10 (log2 fold change=−3.71; p adjusted value=1.36E-51), in agreement with the low IL10 expression at the protein level (figure 4C). Transcriptomic results for IL10 and LTC4S were confirmed by Q-RT-PCR (online supplemental figure 4B).
Supplemental material
KLRG1- ILC2 are enriched in the skin of hypochlorous acid-treated mice and their numbers correlate with the extent of skin fibrosis
To assess the in vivo relevance of our findings, we monitored ILC2 in the hypochlorous acid (HOCl)-induced mouse model of SSc, which recapitulates the main hallmarks of SSc.22 At day 42, parallel to an increase in dermal thickness (figure 6A for a representative histology of the skin, figure 6B,C for skin thickness and collagen content, respectively), we observed a significant enrichment of the lymphoid cell infiltrate (figure 6D,E). Remarkably, while the frequency of ILCs was not changed (figure 6F), the frequency of ILC2 cells was more than 5 times greater in HOCl-treated skin mice compared with controls as detected by flow cytometry (figure 6G). In addition, the absolute value of cutaneous ILC2 was significantly higher in HOCl-treated mice compared with control mouse (online supplemental figure 5A). This ILC2 enrichment was specific, since the frequency and absolute count of total ILCs was not different in HOCl-treated and non-treated mice (figure 6F and online supplemental figure 5B). A slighter increase in ILC2 number and absolute count was already observed even before complete establishment of skin fibrosis (online supplemental figure 5C,D and E,F for skin thickness and collagen content, respectively). Furthermore, the frequency of KLRG1 in ILC2 was fourfold lower in HOCl-treated compared with control mice (figure 6H for a representative staining of ILC2 and figure 6I for quantification). Interestingly, we observed a strong positive correlation between the proportion of KLRG1- ILC2 and the extent of skin fibrosis (figure 6J).
Supplemental material

Characterisation of innate lymphoid cells (ILC)-2 (ILC2) in the skin of phosphate-buffered saline (PBS)-treated and hypochlorous acid (HOCl)-treated mice. (A) Picrosirius red staining of the control and systemic sclerosis (SSc) skin mice at day 42 (scale bar=10 µm). (B and C) Measure of skin thickness (μm) and collagen (μg) in the skin of PBS-treated and HOCl-treated mice at day 42. (D) Representative dot plot of cutaneous ILCs (CD45+, Lin-, CD127+) and ILC2 (CD45+, Lin-, CD127+, CD25+) in both mice skin. (E) Percentage of CD45+ cells in the skin of PBS-treated and HOCl-treated mice at day 42. (F) Percentage of ILC and (G) ILC2, among lymphoid cells in the skin of PBS-treated and HOCl-treated mice at day 42. (H and I) Percentage of KLRG1+ cells among ILC2 in the skin of PBS-treated and HOCl-treated mice at day 42. (J) Positive correlation between the quantity of collagen (μg) and the percentage of KLRG1- ILC2. Bar graphs show data as the mean±SEM (n=10 per groups). Comparisons between groups were calculated using Mann-Whitney U test. **P<0.01; ***p<0.001; ****p<0.0001.
The combined administration of IL10 and TGFβ inhibition is required to restore skin KLRG1+ ILC2 and to prevent skin fibrosis
We have shown that the TGFβ-priming of human KLRG1- ILC2 enhances their profibrotic potential by specifically reducing their production of IL10, thus attenuating a negative feedback loop. We then addressed the question of the in vivo relevance of these findings by taking advantage of the HOCl mouse model of SSc. To inhibit TGFβ signalling, HOCl-treated mice received daily oral pirfenidone for 42 days, or PBS as control. In addition, they were injected or not with daily IL10. At day 42, the treatment with pirfenidone alone moderately reduced, without reaching statistical significance, the fibrotic response assessed as skin thickness and collagen content (figure 7A,B). The treatment with IL10 by itself had no effect on skin fibrosis. In contrast, when pirfenidone and IL10 were administered jointly we observed a dramatic and statistically significant reduction in both the skin thickness and collagen content (figure 7B). Interestingly, the proportion of ILC2 was significantly lower and the frequency of their KLRG1+ subset was significantly higher in HOCl mice that received the combined treatment with pirfenidone and IL10 when compared with all the other treatments (figure 7C). Not the least, the extent of collagen content in the skin was inversely correlated with the frequency of KLRG1+ ILC2 (figure 7D). Altogether, these results demonstrate that the combined treatment with pirfenidone and IL10 strongly reduces skin fibrosis in vivo simultaneously reducing the number of ILC2 infiltrating the skin and enhancing their expression of KLRG1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Interleukin (IL)10 and pirfenidone (Pirf) reduce fibrosis and affect the number of innate lymphoid cells (ILC)-2 (ILC2) in the skin of hypochlorous acid (HOCl)-treated mice at day 42. (A) H&E saffron staining of HOCl skin mice treated or untreated (scale bar=10 µm). (B) Measure of skin thickness (μm) and collagen (μg) in the skin of HOCl-untreated and HOCl-treated mice at day 42. (C) Percentage of ILC2 among lymphoid cells and percentage of KLRG1+ cells among ILC2 in the skin of HOCl-untreated and HOCl-treated mice at day 42. (D) Negative correlation between collagen assay (μg) and the percentage of KLRG1+ ILC2 in the skin. Bar graphs show data as the mean±SEM (n=10 per groups). Comparisons between groups were calculated using Kruskal-Wallis test. *P<0.05; **p<0.01.
Discussion
ILCs were recently identified as new important actors of the innate arm of the immune system. ILCs have been characterised and classified in recent years, and they have been directly implicated in many inflammatory conditions, including fibrosis, atopic dermatitis, asthma and inflammatory bowel disease.23–26 SSc is a complex autoimmune connective tissue disease characterised by autoimmunity, widespread tissue fibrosis of the skin and internal organs and vasculopathic alterations. Its pathogenesis remains poorly understood, and a single treatment has not been approved over the last 50 years.
Our study directly implicates the impairment of ILC homeostasis as a potential contributor and new therapeutic strategy for SSc. Specifically, we observed that human skin biopsies from patients with SSc were characterised by a higher proportion of KLRG1- ILC2 positively correlated to the severity of the fibrotic process. Mechanistically, we demonstrated in vitro that TGFβ decreased KLRG1 expression on ILC2 and enhanced their profibrotic function through IL10 downregulation. Interestingly, in a mouse model of SSc recapitulating human findings, TGFβ inhibition associated with IL10 administration prevented the development of fibrosis while repopulating the skin via KLRG1+ ILC2 in a synergistic manner. Altogether, those results revealed a previously undescribed mechanism in human SSc pathogenesis and paved the way for potential alternative therapeutic strategies based on TGFβ blocking associated with IL10 administration.
Few previous studies have investigated the role of ILCs in human SSc. One group described an increased number of CD4+ group 1 ILCs,27 while another group reported an increased proportion of ILC2 in the peripheral blood.19 In contrast, our data showed a decrease in ILC2 numbers/proportions at the blood level and did not identify a difference for ILC1. We cannot exclude that immunomodulatory agents may have participated to the decrease of ILC2 in the blood. However, in the limit of the number of analysed patients, no impact on the increase amount of ILC2 in the skin of patients with SSc was observed. These contradictory observations also raised specific questions about the phenotypic definition of low-number circulating cells. Although key researchers in the field have reviewed this important question,28 differences in gating strategies, antibodies or even methods may explain these discrepancies. In our gating strategy, one may argue that the gate for ILC is too close from the lineage positive cells. As shown in the online supplemental figure 1B, we pay attention to evaluate that the population edging the lineage positive cells also contained ILC1, ILC2 and ILC3 subsets. Regarding selected markers in our experiments, we used the CD5 in the lineage marker to remove CD4+ T lymphocytes. A recent paper showed that CD5+ ILCs are functionally immature and very close to progenitor cells.29 Although CD5 could be expressed in a small proportion of ILC1, ILC2 and ILC3, these cells become functionally active cytokine-secreting ILCs when they downregulate CD5 and migrate to tissues. Therefore, using CD5 as we have done, appears to be a reasonable trade-off for eliminating CD4 T cells without losing mature cells. Finally, by using two complementary strategies, we observed an increase in the proportion of ILC2 in the SSc skin in accordance with a previously published report.19
Extensive characterisation of human skin-homing ILC2 from patients with SSc revealed subtle phenotypic changes associated with the fibrotic process. SSc skin is populated by KLRG1- ILC2, whereas skin-resident ILC2 express KLRG1 in normal skin from humans and mice. Interestingly, KLRG1 expression levels have been described as a marker to separate iILC2 (KLRG1+) and nILC2 (KLRG1-). iILC2 could be transient progenitors and develop into nILC2 or ILC3-like cells.30 Although this dichotomy has mainly been described in the field of infection in mice, our results show that this observation is true in humans and would be directly associated with the fibrotic process. Our results in SSc suggest KLRG1+ ILC2 could migrate from the blood to the skin and switch into profibrotic KLRG1- ILC2. Functional studies in vitro revealed that TGFβ, which is known to be elevated in patients with SSc, was a key cytokine involved in KLRG1 downregulation, thus mirroring the effect of TGFb on KLRG1-expressing CD8+ T cells.31 Unexpectedly, we observed that KLRG1- ILC2 triggered profibrotic responses on dermal fibroblast, which was mainly due to an imbalance in production between profibrotic factors and antifibrotic factors. Moreover, we observed that the low capability to secrete IL10 was directly responsible for the profibrotic effect. The role of IL10 as an antifibrotic agent has been previously demonstrated. Indeed, two distinct models of fibrosis, one in chronic renal insufficiency and another in hepatic damage induced by biliary duct ligation, provide evidence for the impact of decreased levels of IL10 in the occurrence and maintenance of fibrosis.32 33 Moreover, the genetic delivery of IL10 significantly attenuated TGFβ production in the lungs of mice subjected to bleomycin-induced pulmonary fibrosis. This effect was still observed when IL10 was delivered at later time points when fibrosis was already established.34 Therefore, our results established a direct link between TGFβ and low secretion of IL10 at the tissue level through ILC2.
To look further on the fibrotic potential of ILC2, we evaluated the transcriptome of TGFβ-stimulated ILC2 compared with unstimulated ILC2 and found an increase of the leukotriene C4 synthase expression, suggesting that activated ILC2 may release leukotriene C4 (LTC4). Since LTC4 is a potent inducer of collagen synthesis by dermal fibroblasts,35 the ILC2-LTC4 axis in SSc skin fibrosis will require further investigations.
Different SSc mice models have been used to evaluate different therapeutic strategies; however, none of them recapitulates the human observations in integro. The HOCl-treated mouse model has the advantage of summarising the three main characteristics of SSc: vasculopathy, deregulation of the immune response (including the production of autoantibodies) and cutaneous and pulmonary fibrosis. In our study, we only tested our hypothesis in one mouse model that constitutes a limitation. However, our data indicated that this model recapitulated the human skin findings, including the increased proportion of KLRG1- skin ILC2 proportions correlated to the fibrotic process. We were not able to increase lung fibrosis, precluding any further pulmonary investigation. Our work is thus limited to only one mouse model exploring cutaneous fibrosis. Many mouse models potentially useful to further assess the relevance of ILC in experimental animals could be considered, of which none fully recapitulates the features of human systemic sclerosis.36 However, we believe that, within its limits, the HOCl mouse model we adopted, strongly support the results of our research principally and extensively made in humans. In detail, it met several important requirements for our demonstration: (i) development of skin fibrosis after an inflammatory period; (ii) accompanied by an increase of ILC2 in the skin; (iii) a positive correlation between skin fibrosis and number of ILC2 in the skin, as we have shown in humans. From a therapeutic perspective, most of the mouse models have implicated ILC2 through its deletion using Rorasg/floxIl7rCre/+ mice37 or Rag1-/- mice, for instance, coupled with the injection of depleting anti-Thy1 antibody.38 Although these strategies offer a clear view of the global implication of ILC2 in a model, it is not a feasible strategy in humans and does not permit the characterisation of the implication of ILC2-skewed functionalities in a disease process, as it is the case in our setting. Therefore, we decided to adopt an alternative strategy that combines a well-accepted TGFβ inhibitor and IL10 supplementation, according to our in vitro data. Our results revealed a dramatic synergic effect on the fibrotic process while KLGR1+ ILC2 were repopulating the skin. Although this therapeutic strategy does not formally directly prove the involvement of the KLRG1- population in the pathogenesis, it provides indirect evidence on the pathogenic loop implicating ILC2, IL10 and TGFβ. Moreover, our results could constitute the groundwork for proposing a clinical trial testing the combination of the TGFβ inhibitor and IL10 in patients with SSc. The US Food and Drug Administration recently approved targeting the TGFβ pathway for the treatment of idiopathic pulmonary fibrosis (IPF). In a phase III clinical trial, pirfenidone successfully reduced the progression of IPF and was associated with fewer deaths.39 For unknown reasons, this effect in the patients with SSc seemed to be milder; thus, combining another strategy in this multifactorial disease could be an option. After the successful preclinical experiments, clinical studies using human recombinant IL10 are already in progress for the treatment of inflammatory bowel diseases with an acceptable safety profile.40 Future studies using nintedanib instead of pirfenidone are also of interest since this molecule is used in interstitial lung disease in SSc.41
In conclusion, our study revealed that ILC2 may contribute to the fibrotic process observed in human SSc and the combination of the TGFβ inhibitor and IL10 could be a promising therapeutic strategy.
Supplemental material
Data availability statement
Data are available on reasonable request.
Ethics statements
Ethics approval
Ethical approval information: human participants: Patients were included in the context of the VISS (Vasculopathy and Inflammation in Systemic Sclerosis) biomedical research project founded in 2012 and approved by the institutional ethical committee (CPP, 2012-A00081-42, Aquitaine). All participants provided written informed consent before inclusion. Ethical approval information: animals: Comité d'éthique pour l'expérimentation animale de Bordeaux (CE50) (project n°15544).
Acknowledgments
Some of the data were presented in an abstract at the EULAR annual meeting in 2016. Victor Racine and the QuantaCell society are acknowledged for their help with the analysis of the human imaging data. The histological studies were performed at the Experimental Histopathology Platform, INSERM US 005-CNRS UMS 3427-TBM CORE, a service unit of the CNRS-INSERM and Bordeaux University. The help of Nathalie Dugot-Senant is acknowledged. The microscopy analyses were performed at the Bordeaux Imaging Centre, a service unit of the CNRS-INSERM and Bordeaux University, member of the national infrastructure France BioImaging supported by the French National Research Agency (ANR-10-INBS-04). The help of Sébastien Marais is acknowledged. We are also grateful to Atika Zouine and Vincent Pitard for their expert assistance in flow cytometry (Plateforme de Cytométrie en Flux, SFR TransBioMed, Université Bordeaux), Xavier Gauthereau for assistance in PCR (Plateforme de Q-PCR SFR TransBioMed, Université Bordeaux) and Benoit Rousseau for help in the animal facility (Animalerie A2, Université Bordeaux). RNAseq experiment were realised with the genom'IC platform (INSERM U1016, Institut Cochin). Part of the computational resources and infrastructure used in present publication were provided by the Bordeaux Bioinformatics Centre (CBiB). Eric Vivier and Frédéric Vély are acknowledged for helping us in the setting of ILC staining. Andreas Ramming and Jörg Distler are acknowledged for providing stimulating discussions on the data.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
PL and BA are joint first authors.
Handling editor Josef S Smolen
CC-B and M-ET contributed equally.
Collaborators Victor Racine, Nathalie Dugot-Senant, Sébastien Marais, Atika Zouine, Vincent Pitard, Xavier Gauthereau, Benoit Rousseau, Eric Vivier, Frédéric Vély, Andreas Ramming, Jörg Distler.
Contributors PL and BA jointly designed the research, performed the experiments, collected, analysed and interpreted the data, wrote and revised the manuscript. MJ, PM, EL, DL, AG and VJ performed and analysed some experiments. EL, PL, JI, MJ and FB performed mice experiments. TS, CC, TP and PB provided intellectual input and edited the manuscript. PM, DL, PH, JS, JC, CR, PD, TS, EL and EF recruited the study participants and provided patients’ samples and clinical data. CC-B and M-ET jointly designed and supervised the study and wrote the manuscript.
Funding This research was supported by grants from the Société Française de Rhumatologie (SFR), Rhumato-Network. It was also supported by the Association des Sclérodermiques de France (ASF).
Competing interests None declared.
Provenance and peer review Not commissioned; externally peer reviewed.
Supplemental material This content has been supplied by the author(s). It has not been vetted by BMJ Publishing Group Limited (BMJ) and may not have been peer-reviewed. Any opinions or recommendations discussed are solely those of the author(s) and are not endorsed by BMJ. BMJ disclaims all liability and responsibility arising from any reliance placed on the content. Where the content includes any translated material, BMJ does not warrant the accuracy and reliability of the translations (including but not limited to local regulations, clinical guidelines, terminology, drug names and drug dosages), and is not responsible for any error and/or omissions arising from translation and adaptation or otherwise.