Article Text

Abstract
Objective Comprehending the mechanisms that regulate activation of autoreactive T cells and B cell antibody production is fundamental for understanding the breakdown in self-tolerance and development of autoimmunity. Here we studied the role of Fms-like tyrosine kinase 3 ligand (Flt3L) signalling in the pathogenesis of collagen-induced arthritis (CIA).
Methods CIA was induced in mice lacking Flt3L (Flt3L−/−) and wild-type (WT) littermates (C57/BL6, 8–10 weeks old). Mice were killed in the initial phase (acute phase: experiment 1) and late phase (chronic phase: experiment 2) of the disease. Arthritis severity was assessed using a semiquantitative scoring system (0–4), and histological analysis of cellular infiltration, cartilage destruction and peptidoglycan loss was performed. Phenotypic and functional analysis of T and B cells, FoxP3 expression, activation and lymphocyte costimulatory markers, and cytokine production were performed ex vivo by flow cytometry in lymph nodes. Serum collagen type II (CII)-specific antibodies were measured by ELISA.
Results Flt3L−/− mice showed a marked decrease in clinical arthritis scores and incidence of arthritis in both acute and chronic phases of CIA compared with WT mice. Moreover, decreased synovial inflammation and joint destruction was observed. Both the magnitude and quality of T cell responses were altered in Flt3L−/−. In the acute phase, the amount of CII-specific IgG2a antibodies was lower in Flt3L−/− than WT mice.
Conclusions These results strongly suggest a role for Flt3L signalling in the development of arthritis.
- Arthritis
- Inflammation
- Cytokines
- T Cells
Statistics from Altmetric.com
Introduction
Dendritic cells (DCs) are key players in the induction and maintenance of adaptive immunity. Interestingly, although the role of T and B lymphocytes in autoimmunity is well described in both humans and mice,1–4 and indeed biological agents targeting these cell lineages are currently used in the treatment of rheumatoid arthritis (RA) and systemic lupus erythematosus,5 ,6 far less is known about the specific involvement of DCs in autoimmune disease. In this respect, it is widely assumed that, because of the fundamental role of DCs in initiating T cell responses, DCs might play an essential pathological role in autoimmune disease by presenting self-antigens to autoreactive T cells. Primary immune responses are initiated in the periphery, where DCs capture and process antigens, increase expression of lymphocyte costimulatory molecules, migrate to lymphoid organs, and secrete cytokines that differentiate naïve T cells.7 It was shown that adoptive transfer of DCs pulsed with collagen type II (CII) was able to initiate experimental arthritis in mice,8 thus showing the crucial role of these cells in the induction of arthritis. However, DCs not only activate lymphocytes, but are also capable of tolerising T cells to self or innocuous antigens, thereby minimising autoimmune reactions.9 These dual functions of DCs depend on DC maturation status, cytokine profile and DC subset. DCs are a very heterogeneous population of cells, and different subsets can have very distinct functions. They are important in the breaching of self-tolerance in autoimmunity,10 and plasmacytoid (p)DCs play an important anti-inflammatory role in the context of articular breach of tolerance.11 It was recently shown that compounds that selectively deplete DCs can ameliorate arthritis12 and that blocking tyrosine kinases may have beneficial effects in arthritis by modulating DC numbers.13 DCs can be efficiently differentiated in vitro by stimulating monocytes or haematopoietic progenitors with granulocyte macrophage colony-stimulating factor (GM-CSF).14 ,15 Surprisingly, mice lacking GM-CSF or its receptor showed only a small decrease in lymphoid-organ DCs, with a maximum reduction of threefold in lymph node conventional/classical DCs (cDCs) and only a modest reduction in langerhans cells (LC), whereas GM-CSF transgenic mice showed reciprocal effects.16 ,17 Therefore, at least in the presence of compensatory cytokines, GM-CSF seemed to have little influence on steady-state DC maintenance, and it has been suggested that GM-CSF mostly contributes to inflammatory DC generation, potentially from monocytes, in vivo.18–21 The Fms-like tyrosine kinase 3 (Flt3)/Flt3L axis is crucial for DC development from bone marrow progenitors but not from monocytes.22–24 In line with this, it has been shown that DC development is confined to haematopoietic precursors in bone marrow expressing Flt3.22 ,25 Flt3 signalling can also instruct Flt3-negative precursors to differentiate into both pDCs and cDCs.26 Flt3L is involved in all lymphoid-organ DC development and expansion, from early progenitors in the bone marrow to immediate DC progenitors in lymphoid tissues.22 ,27 ,28 Flt3L is expressed as a membrane-bound protein on the cell surface and can be proteolytically cleaved to generate a soluble protein. Both the membrane-bound and soluble forms are biologically active.29 Administration of Flt3L induces a large increase in the number of DCs in the spleen and lymph nodes (LNs) as well as in the peripheral blood, lungs, liver, Peyer's patches and thymus.30–32 Flt3L-expanded DCs can process and present antigen to naive T lymphocytes both in vitro and in vivo.30 In Flt3L−/− mice, B cell precursors, splenic NK cells and lymphoid tissue DCs are severely reduced.33 Leucocyte cellularity is reduced in the bone marrow, peripheral blood, LNs and spleen. Thymic cellularity, blood haematocrit and platelet numbers are not altered. T and B cells from spleen and LNs are reduced in number, but the ratio of T cells to B cells is unchanged in Flt3L−/− mice compared with wild-type (WT) mice. Histochemical analyses of spleen, thymus and LNs revealed no disruption to normal architecture in the tissues of Flt3L−/− mice.33
It has been shown that Flt3L- and GM-CSF-derived DCs are phenotypically and also functionally different,34 and they can activate different T cell subsets in vivo and in vitro.28 ,34
Flt3L accumulates in RA synovial fluid and induces arthritis when injected into healthy mouse knee joints. In addition, administration of Flt3L worsens experimental arthritis.35 However, the exact mechanism by which Flt3L affects the development of arthritis remains unknown.
Here we took advantage of animals with targeted deletion of Flt3L to provide more insight into the role of Flt3L signalling in the pathogenesis of collagen-induced arthritis (CIA).
Materials and methods
Animals
Male and female Flt3L−/− mice and WT littermates about 8–10 weeks old were used for the experiments. Further details are described in online supplementary methods.
Induction and assessment of CIA
We evaluated the role of Flt3L in CIA using Flt3L−/− and WT mice (n=10–15 per group). Mice were immunised as previously described.36 ,37 Two independent experiments—acute (day 43) and chronic (day 60) stages—were performed. Further details are given in online supplementary methods.
Histological analysis
Hind paws were fixed for 24 h in 4% buffered formalin and decalcified in Osteosoft (Merck, Darmstadt, Germany). After embedding in paraffin, serial 5 μm sagittal sections of whole hind paws were cut and stained with hematoxylin/eosin (H&E), toluidine blue and safranin O-fast green. Further details are described in online supplementary methods.
Cell staining and flow cytometry
Single-cell suspensions were obtained from spleen and axillary and inguinal LNs. After erythrocyte lysis, cells were stained with the indicated fluorochrome-conjugated antibodies for surface markers and intracellular cytokines. Further details are described in online supplementary methods.
Measurement of antigen-specific antibodies and their affinity in serum
Serum levels of antibodies against chicken collagen type II (cCII) were measured by ELISA. Further details are described in online supplementary methods.
T cell proliferation in vitro
Carboxyfluorescein diacetate, succinimidyl ester (CFSE)-labelled ovalbumin (OVA)-specific CD4 T cells from OT II mice (2×105 cells) were stimulated for 3 days with OVA-pulsed bone marrow-derived DCs from WT and Flt3L−/− mice. T cell proliferation was measured by fluorescence-activated cell sorting (FACS). Further details are described in online supplementary methods.
Statistical analysis
Areas under the curve for the change in arthritis scores were calculated. Incidence was compared using Kaplan–Meier survival analysis (GraphPad Prism V.5 ). The mean changes in scores (clinical, histological and immunohistochemical), total cell numbers, and cytokine- and collagen-specific IgG1 and IgG2a antibody levels between groups were determined by Mann–Whitney U test. p<0.05 was considered significant.
Results
Reduced incidence and severity of CIA in Flt3L-deficient mice
In agreement with the original description of these mice,33 mice lacking Flt3L developed normally and showed no gross anatomical defects or phenotypic abnormalities. Flt3L−/− mice in homoeostatic conditions showed a dramatic reduction in DC numbers in lymphoid organs (see online supplementary figure 1A, B p=0.0079). After induction of arthritis, all mice were killed at day 60 (the point when arthritis reached the chronic phase) or at day 43 (during the acute phase of the disease). A dramatic improvement in clinical signs of arthritis was seen in arthritic Flt3L−/− mice compared with arthritic WT mice (figure 1A, p=0.0003, and 1C, p<0.0001) for both chronic and acute phases of the disease, respectively. In addition, WT mice showed accelerated onset of disease and significantly higher disease incidence (figure 1B, p<0.0001, and 1D, p<0.0001).
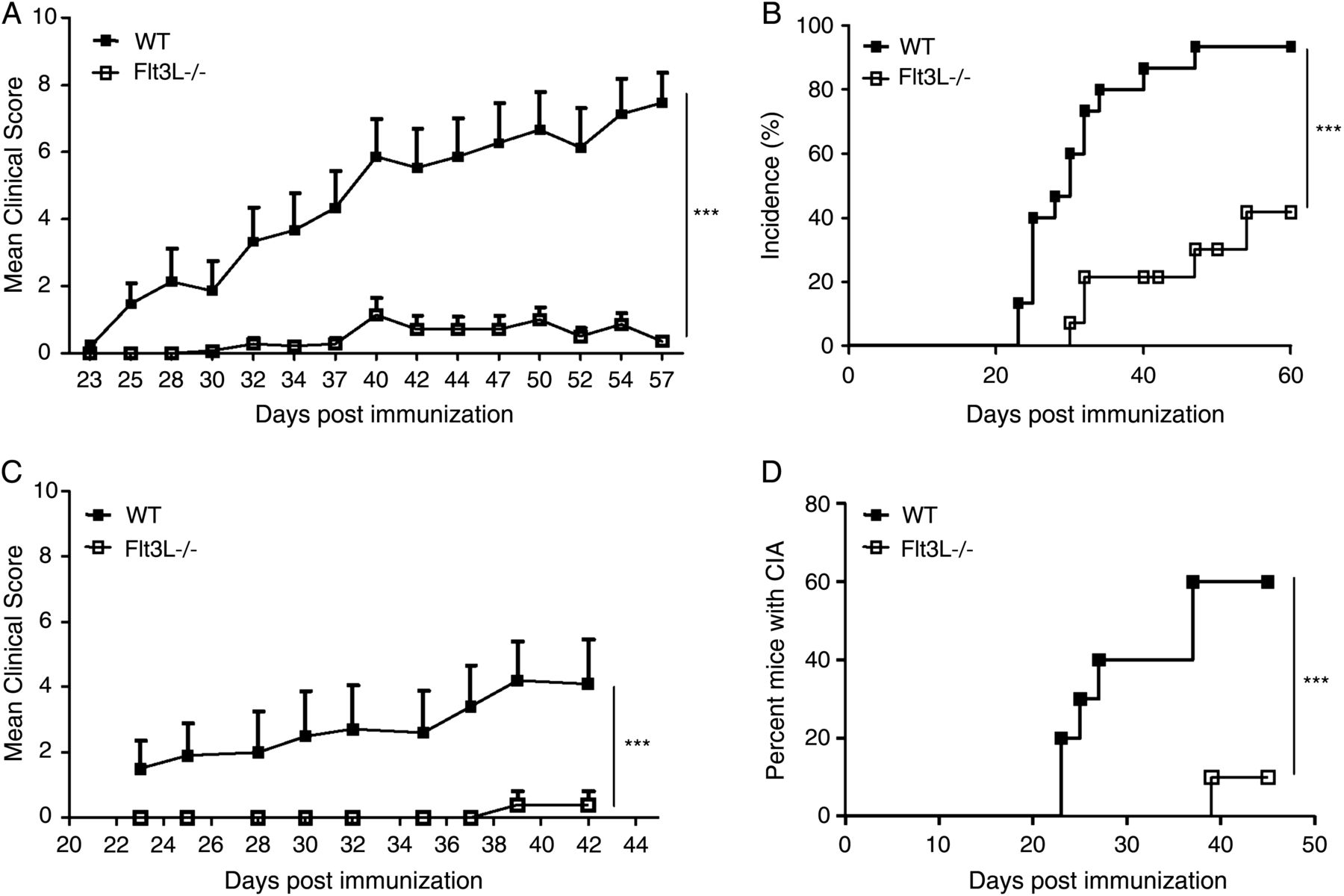
Development of collagen-induced arthritis (CIA) in wild-type (WT) and Flt3L−/− mice in the chronic (experiment 1; day 60; WT, n=14; Flt3L−/−, n=14) and acute (experiment 2; day 43; WT, n=10; Flt3L−/−, n=10) phases of the disease. (A) Flt3L−/− mice showed a dramatic decrease in arthritis scores (chronic phase) compared with WT mice. All mice are shown. Values are presented as the mean±SEM. (B) Chronic phase cumulative disease incidence scores in WT and Flt3L−/− mice calculated as the percentage of affected animals. Arthritic Flt3L−/− mice showed a significant decrease in disease incidence compared with WT mice. (C) Flt3L−/− mice showed a striking decrease in arthritis scores (acute phase) compared with WT mice. All mice are shown. Values are presented as the mean±SEM. (D) Acute phase cumulative disease incidence scores in WT and Flt3L−/− mice calculated as the percentage of affected animals. Arthritic Flt3L−/− mice showed a significant decrease in disease incidence compared with WT mice. ***p<0.0001.
Decreased cellular infiltration and cartilage degradation in Flt3L−/− mice in the chronic phase of the disease
We next analysed whether Flt3L deficiency would affect synovial inflammation. Flt3L−/− mice showed a significant decrease in cellular influx (figure 2A, D; p=0.0018) into the synovium at day 60 compared with WT mice (figure 2A, B, D). In addition, depletion of proteoglycans, assessed by safranin O-fast green staining, was significantly less in the knee joints of Flt3L−/− mice compared with WT mice (figure 2C, D; p=0.0339). We did not observe any significant differences in cartilage damage in Flt3L−/− mice and WT mice (figure 2B).

Histological analysis of wild-type (WT) and Flt3L−/− knee joints in the chronic (day 60; WT, n=14; Flt3L−/−, n=14) phase of the disease. (A) Representative H&E staining of the knee joints of a WT mouse and a Flt3L−/− mouse. (B) Representative toluidine blue staining of the knee joints of a WT mouse and an Flt3L−/− mouse. (C) Representative safranin O-fast green staining of the knee joints of a WT mouse and an Flt3L−/− mouse. (D) Semiquantitative scores for synovial infiltration, cartilage damage and proteoglycan (PG) depletion assessed by H&E, toluidine blue and safranin O-fast green staining, respectively. Flt3L−/− mice show less synovial infiltration and less proteoglycan depletion compared with WT mice. Data are represented as the mean±SEM. *p<0.05, ***p<0.0001. Magnification ×100.
T cell responses during CIA in Flt3L−/− mice: reduced cytokine production, activation and costimulatory markers
Total CD4 and CD8 T cell numbers and phenotype in inguinal and axillary LNs during the chronic phase of CIA on day 60 were assessed by FACS analysis. We observed a significant reduction in CD8 T cell numbers (figure 3A, p=0.0259) and a trend towards lower CD4 T cell numbers. We phenotypically analysed CD4 and CD8 compartments for naïve and effector memory based on the expression of CD44 and CD62L. While there were no differences in the amount of CD4 and CD8 naive (CD44−CD62L+) T cells between Flt3L−/− and WT mice, we observed a reduction in the CD4 effector memory (CD44+CD62L−) (figure 3B, p<0.0001) and CD8 effector memory (CD44+CD62L−) compartments (figure 3C, p=0.0008). In addition, a reduction in total B cell numbers (figure 3D, p<0.0001) was observed in Flt3L−/− compared with WT mice.

Phenotypic characterisation of T and B cells in wild-type (WT) and Flt3L−/− mice in chronic collagen-induced arthritis (WT, n=14; Flt3L−/−, n=14). (A) Total cell numbers of CD4 and CD8 T cells in inguinal and axillary lymph nodes (LNs). (B) Naive (N) CD44−CD62L+ and effector memory (EM) CD44+CD62L+ CD4+ T cells present in the LNs. We observed a reduction in EM in the CD4 T cell compartment in Flt3L−/− mice compared with WT mice. (C) N CD44−CD62L+ and EM CD44+CD62L−, CD8 T cells present in the LNs of WT and Flt3L−/− mice (n=14 per group). We observed a reduction in EM in the CD8 T cell compartment in Flt3L−/− mice compared with WT mice. (D) Reduced number of total B220+ B cells in the LNs. Data are represented as the mean±SEM. *p<0.05, ***p<0.0001.
Next, the production of Th1 and Th17 cytokines, involved in the development of CIA, was assessed by intracellular staining of phorbol myristate/ionomycin-stimulated LN cells. We observed a significant reduction in the number of CD4 T cells that produced interleukin (IL)-17 (p=0.0014) and tumour necrosis factor (TNF)α (p=0.0072) in Flt3L−/− mice compared with WT mice (figure 4A). In addition, a significant reduction in the number of CD8 T cells that produced interferon (IFN)γ (p=0.029) was observed in Flt3L−/− mice compared with WT mice (figure 4A). Not only was the number of T cells that produced cytokines affected in Flt3L−/− mice, but the amount produced per cell basis was also significantly lower. The mean fluorescence intensity is a measurement of expression per cell basis. It was calculated only for the positive cell populations for each cytokine. In this respect, CD4 T cells in Flt3L−/− mice produced significantly less IFNγ (p=0.01) and TNFα (p=0.0003), and CD8 T cells had a higher capacity to produce IL-17 (p=0.0067) than WT mice (figure 4B). No differences were observed between WT and Flt3L−/− mice with respect to IL-2 or IL-10 production (figure 4A, B).

Changes in cytokine responses of T cells in wild-type (WT) and Flt3L−/− mice during the chronic phase of CIA. Lymph nodes (LNs) of WT mice (n=14) and Flt3L−/− (n=14) were collected 60 days after primary immunisation, and intracellular cytokine production was assessed by flow cytometry. (A) Total cell numbers of cells producing interleukin (IL)-2, interferon (IFN)γ, tumour necrosis factor (TNF)α, IL-10 and IL-17 in CD3+CD4+ and CD3+CD8+ T cells after 2 h of stimulation with phorbol myristate/ionomycin plus 4 h with brefeldin are shown. A significant reduction in the numbers of CD4 T cells that produced IL-17 (p=0.0014) and TNFα (p=0.0072) was observed in Flt3L−/− mice compared with WT. In addition, a significant reduction in CD8 T cells that produced IFNγ (p<0.0001) was observed in Flt3L−/− mice compared with WT. (B) Mean fluorescence intensity (MFI) for IL-2, IFNγ, TNFα, IL-10 and IL-17 in CD3+CD4+ and CD3+CD8+ T cells. (C) Total CD4 T cell numbers expressing CD25. Flt3L−/− mice have reduced numbers of CD4+CD25+ T cells compared with WT (p<0.0001). (D) Total CD25+FoxP3+ regulatory T cells numbers. A significant reduction in the number of T regs was observed in Flt3L−/− animals compared to WT. *p<0.05, ***p<0.0001.
To compare the overall state of activation of T cells, we calculated the percentage of activated CD4+CD25+ T cells in the LN of Flt3L−/− mice and compared this with WT mice. We observed a significant reduction (p=0.03) in total cell numbers of activated CD4 T cells (figure 4C). The total number of FoxP3+ regulatory T cells (T regs) is also diminished in Flt3L−/− animals compared with WT animals (figure 4D).
Reduced DC numbers and changes in DC subsets in Flt3L−/− mice during the acute phase of CIA
The half-life of mature DCs is short, and, since cytokine production is transient, the number and type of DCs present in the T cell areas will mirror the conditions of the tissues from which the lymph is drained.38 To study the number and diversity of the DC populations in CIA, LNs were collected during the acute phase of the disease (day 43; see online supplementary methods), and cells were stained for cellular markers present on DC cells. As previously shown for Flt3L−/− mice in the steady-state33 (see online supplementary figure 1A, B), in CIA conditions also, these mice show a significant reduction in the percentage of total DC numbers, CD11c+MHCII+ in the LNs compared with WT mice (figure 5A, B, p=0.0015). To assess LN DCs in detail, we analysed various DC subsets by further flow cytometric staining. cDCs in the LN are CD11chi cells that differentially express CD8α and CD11b. We also analysed for the presence of migratory CD103 DCs. Since it has been previously shown that pDCs are reduced in Flt3L−/− mice and play an anti-inflammatory role in experimental arthritis,11 ,39 either by suppressing arthritogenic autoimmunity preventing exacerbated breach of tolerance to self-antigens or by inducting IL-10-producing regulatory T cells, we chose to focus on the other DC subsets that are more important for disease induction. Myeloid-related DCs were identified as low-density cells expressing high levels of CD11c and lacking expression of CD8α, whereas lymphoid-related DCs were identified as cells expressing CD11c and being positive or not for CD8α. All CD11cbright cells expressed high levels of major histocompatibility complex class II (IAb). Both the percentages of myeloid-related and lymphoid-related DCs and migratory CD103 DCs were reduced in the LNs of Flt3L−/− mice compared with WT mice (figure 5A, B; p=0.0039, p=0.0007, p<0.0001 and p<0.0001, respectively). Other CD11cint or CD11c populations that expressed high CD11b were likely LN monocytes and macrophages,27 ,40 and these populations were not affected (data not shown). Bone marrow-derived DCs from Flt3L−/− are able to efficiently stimulate antigen-specific T cell proliferation in vitro (see online supplementary figure 2).

Phenotypic characterisation of dendritic cells (DCs) in wild-type (WT) and in Flt3L−/− mice in the acute phase of collagen-induced arthritis (CIA). Inguinal and axillary lymph nodes (LNs) of WT mice (n=10) and Flt3L−/− mice (n=10) were collected 43 days after primary immunisation, and DC subsets were identified by flow cytometry. (A) Representative FACS plot of total DCs (CD11c+MHCII+), myeloid-related DCs (CD11b+CD11c+), lymphoid-related DCs (CD8α+/CD8α− CD11c+) and migratory DCs (CD11c+CD103+) in Flt3L−/− and WT mice are shown. (B) Quantification of total DCs (CD11c+MHCII+), myeloid-related DCs (CD11b+CD11c+) lymphoid-related DCs (CD8α+/CD8α− CD11c+) and migratory DCs (CD11c+CD103+) in Flt3L−/− and WT mice. Flt3L−/− mice showed a reduction in the total CD11c+MHCII+ DC numbers (p=0.0015) compared with WT animals. Both the numbers of CD11b+CD11c+ DCs (p=0.0039), CD8α+/CD8α− CD11c+ DCs (p=0.0007 and p<0.0001) and CD11c+CD103+ DCs (p<0.001) were reduced in the LNs of Flt3L−/− mice compared with WT mice. ***p<0.0001.
CII-specific antibody production
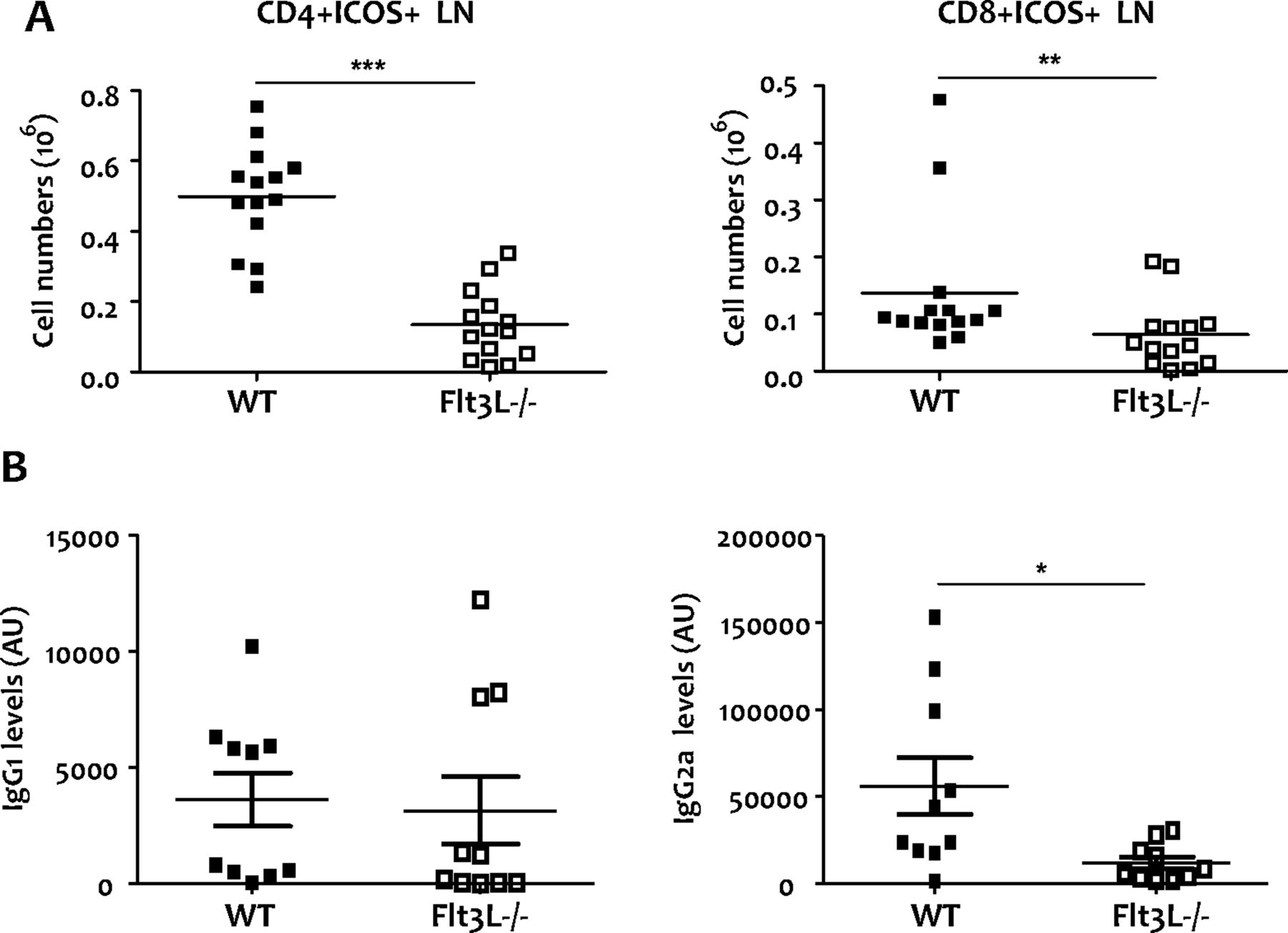
It has previously been reported that expression of the inducible T cell costimulator (ICOS) protein and CD40 ligand play an essential role in promoting effective B cell activation and Ig class switching.41 Accordingly, we observed a significant reduction in ICOS in both CD4 and CD8 T cells from Flt3L−/− mice compared with WT mice (figure 6A, p=0.0001 and p=0.0094, respectively).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of costimulatory molecules in chronic collagen-induced arthritis (CIA) (WT, n=14; Flt3L−/−, n=14) and serum-specific collagen type II (CII) antibodies in wild-type (WT) and Flt3L−/− mice during the acute phase of CIA as assessed by ELISA (WT, n=10; Flt3L−/−, n=10). (A) Total CD4 and CD8 T cells expressing inducible T cell costimulator (ICOS). In Flt3L−/− mice, reduced numbers of CD4 (p<0.001) and CD8 T cells (p=0.0094) that express ICOS were observed compared with WT animals. Data are represented as the mean±SEM. (B) CII-specific IgG1 and IgG2a antibodies during the acute phase. At day 43 we observed similar IgG1 levels but increased IgG2a antibodies in WT compared with Flt3L−/− mice (p=0.02). Levels are shown as arbitrary units (AU). *p<0.05, **p<0.001; ***p<0.0001.
The severity of CIA is reflected by the switch from a Th2 to a Th1 response,42 which is accompanied by a switch from IgG1 to IgG2a antibody production. At day 43, we observed similar IgG1 levels but increased IgG2a antibodies in WT compared with Flt3L−/− mice (figure 6B, p=0.0232). There were no differences in CII-specific IgG2a and IgG1 antibodies at day 60 (see online supplementary figure 3).
Discussion
In this study, we show that CIA in Flt3L-deficient mice is impaired, with a marked decrease in clinical arthritis scores (both in acute and chronic phases of the disease) compared with WT littermates. This was accompanied by a decreased incidence of arthritis, synovial inflammation and joint destruction and diminished levels of proinflammatory cytokines and CII-specific IgG2a antibodies.
Both the magnitude and quality of a T cell response are considered important metrics in evaluating the efficacy of an immune response. The number of responding cells provides a measure of the magnitude, whereas the nature and diversity of the functional responses have been associated with measures of quality. These functions include expression of activation and costimulatory markers and the release of one or more cytokines that induce proliferation and modulate inflammation.43 Here we show changes in both magnitude and quality of the T cell response in the LNs of Flt3L−/− mice compared with WT mice. In this respect, Flt3L−/− mice showed reduced cytokine production and total numbers of activated CD4+CD25+ T cells compared with WT mice. Given that clinical assessment of the disease is made by quantifying local inflammation—by scoring paw swelling—studying the balance between cytokine-producing Th1/Th17 and regulatory T cells at the local site of inflammation would provide clearer results.
Immune responses are initiated in the T cell areas of secondary lymphoid organs where naive T lymphocytes meet DCs, which present antigens taken up in peripheral tissues.7 As shown previously,33 Flt3L−/− mice show a large reduction in DC numbers in lymphoid organs. Here we show that the density and quality of DCs present in the secondary lymphoid organs might account for the changes in the magnitude and class of the T cell response that we observed in Flt3L−/− mice during CIA. We observed reductions in MHCII+CD11C+ DCs as well as specific reductions in CD11b+CD11c+ DC, CD8α+ and CD8α− DC and CD103 DC subsets both in the LNs and spleen (data not shown). In the steady-state, these subsets require Flt3L for their generation.
The availability and quality of antigen-presenting DCs and antigen-specific T cell precursors are limiting factors in the generation of a proper immune response. There is evidence that responding T cells compete in vivo for access to DCs and that this competition can be relieved by increasing total DC numbers44 ,45 and/or increasing specific subsets that can shape the immune response.46 During the initial phase of a primary response, the frequency of naive T cells specific for a given antigen is low, which makes competition among responding cells unlikely. However, as the responding cells proliferate, competition for sustained T cell receptor stimulation will increase, particularly among cells of the same clone, which have the same affinity or avidity and occupy the same niche. Knowing that Flt3L−/− mice have severely reduced numbers of DCs in their secondary lymphoid organs, and given the importance of these cells for the initiation of a proper T cell response, we hypothesised that these mice would have problems in mounting a proper immune response. However, not all DC subsets are dependent on Flt3L for their generation,38 and not all DC populations in these mice are affected,47 making Flt3L-dependent DCs a very important cell type in CIA induction. There are two major pathways of DC development: one driven by Flt3L and its receptor, Flt3, acting on monocyte-independent precursors,19 ,26 ,40 and the other from monocytes independently of the Flt3L/Flt3 system.48 Great emphasis has been given to inflammatory DCs arising from monocytes through the control of GM-CSF-, inflammation-driven monocyte conversion into murine splenic DCs,19 ,20 and DC generation during acute inflammatory arthritis and antigen-induced peritonitis49 have been reported to depend on GM-CSF, emphasising the increasing importance of GM-CSF under inflammatory conditions. Nevertheless, we show that steady-state Flt3L-dependent DCs play a crucial role in the induction of CIA, since Flt3L−/− mice show a dramatic decrease in incidence and severity of arthritis.
Flt3L−/− mice also show a large reduction in NK cell populations. NK cells have been described as having both a protective50 and a pathogenic51 role in autoimmunity, and how they contribute to RA pathology remains to be fully elucidated. Since, in Flt3L−/− mice, NK numbers are reduced, we cannot exclude a possible role for these cells in CIA.
B cell-deficient mice are resistant to CIA, indicating that CII-specific antibodies are important for disease induction.52 It has been reported that expression of the costimulatory proteins, ICOS and CD40 ligand, plays a critical role in promoting effective B cell activation and Ig class switching.41 We observed a reduction in the total numbers of CD4 and CD8 T cells expressing ICOS in Flt3L−/− mice compared with WT animals. In the acute phase of CIA, the amount of IgG2a was significantly lower in Flt3L−/− mice compared with WT animals. Switching to the IgG2a isotype is strongly associated with a typical Th1 response in mice.53 ,54 In agreement with this notion, we observed reduced levels of the prototypic Th1 cytokine, IFNγ, in Flt3L−/− mice. Overall, these results strongly suggest a role for Flt3L signalling in the development of arthritis.
Acknowledgments
We thank Drs K A Reedquist and J Hamann for critically reading the manuscript, and Dr Louis Boon for providing reagents.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement 1
- Data supplement 2 - Online figures
Footnotes
Handling editor Tore K Kvien
Correction notice This article has been corrected since it was published Online First. Figure 5A labels have been corrected.
Contributors MIR drafted the manuscript. All authors revised the manuscript critically for important intellectual content and approved the final version. Study conception and design: MIR, ONK, PB, SA, SEJ, PPT and MCL. Acquisition of data: MIR, ONK, PB and MCL. Analysis and interpretation of data: MIR and MCL.
Funding Supported by grants from the Fundação para a Ciência e a Tecnologia (SFRH/BD/47257/2008 to MIR), Portugal and Dutch Arthritis Association (Reumafonds, project No 7-1-301 to PPT and MCL), The Netherlands.
Competing interests None.
Provenance and peer review Not commissioned; externally peer reviewed.