Article Text

Abstract
Objectives Patients with systemic lupus erythematosus (SLE) and rheumatoid arthritis (RA) have increased risk of cardiovascular disease (CVD). We investigated whether single nucleotide polymorphisms (SNPs) at autoimmunity risk loci were associated with CVD in SLE and RA.
Methods Patients with SLE (n=1045) were genotyped using the 200K Immunochip SNP array (Illumina). The allele frequency was compared between patients with and without different manifestations of CVD. Results were replicated in a second SLE cohort (n=1043) and in an RA cohort (n=824). We analysed publicly available genetic data from general population, performed electrophoretic mobility shift assays and measured cytokine levels and occurrence of antiphospholipid antibodies (aPLs).
Results We identified two new putative risk loci associated with increased risk for CVD in two SLE populations, which remained after adjustment for traditional CVD risk factors. An IL19 risk allele, rs17581834(T) was associated with stroke/myocardial infarction (MI) in SLE (OR 2.3 (1.5 to 3.4), P=8.5×10−5) and RA (OR 2.8 (1.4 to 5.6), P=3.8×10−3), meta-analysis (OR 2.5 (2.0 to 2.9), P=3.5×10−7), but not in population controls. The IL19 risk allele affected protein binding, and SLE patients with the risk allele had increased levels of plasma-IL10 (P=0.004) and aPL (P=0.01). An SRP54-AS1 risk allele, rs799454(G) was associated with stroke/transient ischaemic attack in SLE (OR 1.7 (1.3 to 2.2), P=2.5×10−5) but not in RA. The SRP54-AS1 risk allele is an expression quantitative trait locus for four genes.
Conclusions The IL19 risk allele was associated with stroke/MI in SLE and RA, but not in the general population, indicating that shared immune pathways may be involved in the CVD pathogenesis in inflammatory rheumatic diseases.
- rheumatoid arthritis
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Introduction
Patients with systemic lupus erythematosus (SLE) have a 2–10 fold increased risk of cardiovascular disease (CVD) compared with the general population, with the highest relative risk in younger patients and highest absolute risk in older individuals.1–3 Women with SLE in the age of 35–45 years have a 50-fold increased risk of myocardial infarction (MI) compared with the general population.4 Death related to active disease and infections have decreased, but mortality related to CVD shows no such decline.5 Instead, a slight increase in standardised mortality ratio due to vascular diseases has been reported.6 Today SLE is acknowledged as a unique risk factor for CVD by the American Heart Association.7
Traditional risk factors cannot fully explain the increased risk for CVD in SLE, and a number of SLE-related risk factors have been identified, such as antiphospholipid antibodies (aPL) and renal impairment.8–10 Genetic predisposition is an important risk factor for SLE, and different risk genes are also connected to CVD. We have previously shown that a variant of interferon regulatory factor 8 (IRF8)11 is associated with development of ischaemic heart disease in SLE and that a variant of signal transducer and activator of transcription 4 (STAT4) is associated with aPL and ischaemic stroke.12 Other risk genes shown to be associated with CVD in SLE include mannose-binding lectin, C reactive protein and HLA-DRB1*04/*13.13–15 Recently, a large international association study identified 24 new SLE risk loci using Immunochip genotype data.16 17 In the present study, we examined if single nucleotide polymorphisms (SNPs) analysed by the Immunochip were associated with CVD in two large SLE cohorts and investigated possible functional effects of associated gene variants. We also investigated if identified risk gene variants were associated with CVD in patients with rheumatoid arthritis (RA) and in the general population.
Patients and methods
Patients and controls
The discovery cohort included 1045 patients with SLE from rheumatology clinics in Sweden. All patients fulfilled ≥4 American College of Rheumatology (ACR) 1982 criteria for SLE.18 The replication cohort included 1043 patients with SLE from the University of California, San Francisco (UCSF) Lupus Genetics project.19 All patients completed an extensive questionnaire, and the SLE diagnosis was confirmed by medical record review according to the ACR criteria.18 The SLE patients were all of European decent, and age at diagnosis was similar in both populations, but patients in the replication cohort were younger at follow-up and had less CVD events (table 1). In the discovery cohort, the average age at first stroke/transient ischaemic attack (TIA) was 52 years and at first stroke/MI was 54 years (online supplementary table S1). Information regarding stroke and TIA was available for 814 patients in the SLE discovery cohort and data regarding TIA, and angina was not available for the replication cohort. Patients with RA (n=824) all fulfilled the 1987 RA classification criteria20 (online supplementary method/table S2). Healthy blood donors (n=2711) were recruited as previously described.21 Publicly available data from the CARDIoGRAMplusC4D Consortium22 and the International Stroke Genetics Consortium23 were used for analyses of CVD in the general population (online supplementary methods). All participants gave their informed consent. Definitions for CVD in SLE are in online supplementary table S3 and in RA in supplementary methods.
Supplementary file 1
Clinical characteristics of the patients with SLE
Genotyping and quality control
Genotyping of the SLE discovery cohort, SLE replication cohort and the RA cohort was performed using the Illumina Immunochip (for quality control, see supplementary methods).16 17 24
Statistical analysis
A logistic regression model with sex and disease duration included as covariates was used to test association between SNPs and CVD in the SLE discovery cohort. For the American–European population sex, disease duration and the first principal component for population stratification were included as covariates. The identification of SNPs associated with CVD in SLE included three steps. First, the four CVD variables were tested for association with the Immunochip SNPs using data from the discovery cohort. Next, the top 100 associated SNPs per variable were tested for association with CVD in the SLE replication cohort, and SNPs not achieving nominal significance (P>0.05) were excluded. Finally, SNPs with meta-analysis P<0.001 were chosen as candidates for functional follow-up. The identified interleukin 19 (IL19) and signal recognition particle 54 – antisense 1(SRP54-AS1) SNPs were analysed in the RA cohort, including sex and disease duration as covariates. Resulting significant variants were included in a cross-disease meta-analysis of SLE and RA. In this meta-analysis, P<1.0×10−6 adjusting for 48 000 independent SNPs on the Immunochip were considered significant. In other analyses, p values <0.05 were considered significant. For more information on statistical and bioinformatic analyses, see supplementary methods.
Functional analysis
Electrophoretic mobility shift assays (EMSA) were performed using nuclear extract from Jurkat, LCL, K562, HUVEC cell lines and peripheral blood mononuclear cells (PBMCs) from healthy individuals (supplementary methods). Expression quantitative trait locci (QTLs) were analysed using GTEx datasets,25 and IL19 (R&D Systems), IL10 (Mesoscale Discovery) and aPL levels were measured (supplementary methods).12
Carotid ultrasound
Ultrasound scans were performed as previously described.26
Results
Identification of risk loci for CVD in SLE
The allele frequencies of 137 213 SNPs were compared between patients with SLE in the discovery cohort with and without CVD. Initially, we asked if there are general risk genes for CVD in SLE and consequently analysed ischaemic stroke and/or MI (stroke/MI) as one variable. Next, we looked for subtype-specific risk gene variants analysing MI and/or angina (MI/angina) and ischaemic stroke and/or TIA (stroke/TIA) separately. Finally, venous thrombosis defined as pulmonary embolism and/or deep vein thrombosis (PE/DVT) was analysed. We identified eight SNPs at three different loci demonstrating an association with CVD in both the discovery cohort and in the replication cohort, all reaching P<0.001 in the meta-analysis (tables 2 and 3). The IL19 risk alleles were associated with stroke/MI, the SRP54-AS1 risk alleles, and the IL7 receptor (IL7R) risk alleles were associated with stroke/TIA. Functional studies did not indicate the IL7R risk allele to be important for CVD development (see supplementary file). No SNPs were associated with MI/angina or PE/DVT (data not shown).
SNP variants associated with stroke/MI
SNP variants associated with stroke/TIA
The IL19 locus
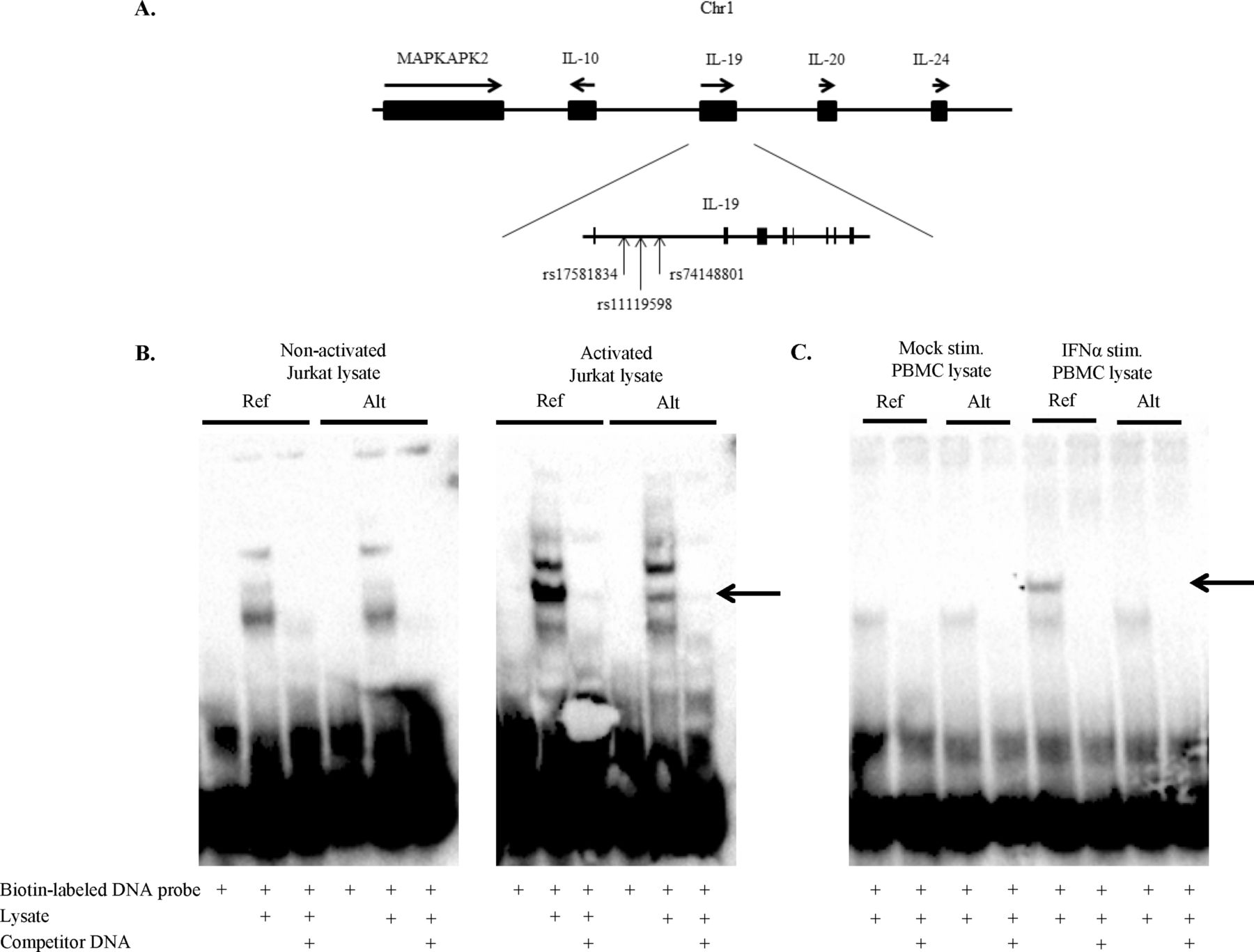
Three SNPs in the IL19 gene (rs74148801, rs17581834 and rs11119598) showed an association with stroke/MI in both SLE cohorts and in the meta-analysis (OR 2.3 (1.5 to 3.4), P=8.5×10−5; table 2). All three SNPs are located in intron 1 of the IL19 gene and are in high linkage disequilibrium (LD) with each other (1000 Genomes, r2=1) (figure 1A). The association between stroke/MI and the IL19 risk allele remained significant when adjusting for known cardiovascular risk factors in a multivariable regression analysis (OR 2.03 (1.07 to 3.84), P=3.09×10−2; online supplementary table S4). The IL19 risk allele was not associated with SLE per se (online supplementary table S5), the ACR criteria (online supplementary table S6)18 or the SLICC-DI (SDI) (OR 1.30 (0.94 to 1.79), P=0.12).24 There was no association between the IL19 risk allele and intima-media thickness (IMT) (0.059 vs 0.063 mm, P=0.92) or presence of carotid plaque (27% vs 21%, P=0.47) in a subgroup of patients with SLE (n=202) examined by carotid ultrasound.

Differential binding to the IL19 risk allele of a protein produced by PBMCs stimulated with IFN-α and Jurkat cells stimulated with PMA/ionomycin. (A) Schematic picture of the IL19 gene region on chromosome 1 with the exons shown as vertical bars and the position of the SNPs are indicated by arrows, dbSNP Build 147, release 108. The SNPs rs17581834, rs11119598 and rs74148801 are associated with stroke/MI in SLE. (B) EMSA of the IL19 locus, rs74148801 using nuclear extract from non-stimulated Jurkat (non-activated) cells and Jurkat cells stimulated with PMA/ionomycin (activated). (C) EMSA of the IL19 locus, rs74148801 using nuclear extract from non-stimulated PBMCs (Mock stim.) and PBMCs stimulated by IFN-α for 20 hours (IFNα stim.). Arrows point at positions of potential differential binding. Biotin-labelled DNA probe is probes of reference and alternative alleles. Lysate is nuclear extract. Competitor DNA is unlabelled DNA probes as competitors in 100-fold excess. Ref.=reference allele (C); Alt.=alternative allele (T). dbSNP, The Single Nucleotide Polymorphism Database; EMSA; electrophoretic mobility shift assay; IFN, interferon; IL, interleukin; MI, myocardial infarction; PMA, phorbol 12-myristate 13-acetate; PBMCs, peripheral mononuclear cells; SLE, systemic lupus erythematosus; SNPs, single nucleotide polymorphisms.
To clarify the function of the IL19 risk allele, we initially performed EMSAs to investigate effects on transcription factor (TF) binding at the locus. Nuclear extract from Jurkat cells stimulated with PMA/ionomycin or PBMC stimulated with interferon-α (IFN-α) demonstrated binding of a protein to the reference allele (C) but not the risk/alternative allele (T) (figure 1B,C). Nuclear extract from unstimulated Jurkat cells or PBMCs displayed no differential binding between the two alleles. Next, protein expression of genes located at the IL19 locus, including IL19 and IL10 (figure 1A), were examined. Serum-IL19 was measured in 394 SLE patients, but no significant difference between patients with and without the IL19 risk allele was observed (24% vs 19%, P=0.41). Plasma-IL10 was measured in 243 patients and patients with the IL19 risk allele more often had elevated IL10 compared with patients without the IL19 risk allele (50% vs 22%, P=0.0038). SLE patients with high IL10 more often had elevated levels of anticardiolipin (aCL) IgM antibodies compared with patients with low IL10 (31% vs 14%, P=0.01). Given these results, we measured the levels of aPL in 781 patients with SLE and observed an association between the IL19 risk allele and elevated levels of aCL IgG (P=0.002), aCL IgM (P=0.002), anti-β2 glycoprotein-I (anti-β2GPI) IgG (P=0.0004) and antiprothrombin (n=494) IgG (P=0.04) antibodies. Furthermore, the IL19 risk allele showed an association with positive lupus anticoagulant (LA; n=311) test (P=0.03).
The Signal recognition particle 54 – antisense 1 locus
Three SNPs (rs799454, rs1712349 and rs712308) located in the SRP54-AS1 gene and in high LD with each other (r2=1) (figure 2A) were associated with stroke/TIA (table 3) with an OR of 1.7 (1.3 to 2.2), P=2.5×10−5 in the SLE meta-analysis. When including the SRP54-AS1 risk allele together with traditional cardiovascular risk factors, the association remained (OR 1.65 (1.15 to 2.37), P=6.10×10−3; online supplementary table S7).The SRP54-AS1 risk allele was not associated with SLE (online supplementary table S5), the ACR criteria, levels of aPL, IMT or carotid plaque (data not shown, all P>0.05).

{kind=link}
{kind=link}
Using recombinant protein, STAT1 is demonstrated to be one of the transcription factors binding at the SRP54-AS1 locus. (A) Schematic picture of the SRP54-AS1 gene region on chromosome 14. The exons are shown as vertical bars, and the position of the SNPs are indicated by arrows, dbSNP Build 147, release 108. The SNPs rs799454, rs1712349 and rs712308 are associated with stroke/TIA. (B) EMSA analysis of the SRP54-AS1 locus, rs1712349. Arrow points at position of potential differential binding of STAT1. Biotin-labelled DNA probe are probes of reference and alternative alleles. STAT1 protein is recombinant STAT1 protein. Lysate is nuclear extract from non-activated Jurkat cells. Competitor DNA is unlabelled DNA probes as competitors in 100-fold excess. Ref.=reference allele (C); Alt.=alternative allele (T). dbSNP, The Short Genetic Variations database; EMSA; electrophoretic mobility shift assay; SNP, single nucleotide polymorphisms; SRP54-AS1, signal recognition particle 54 – antisense 1; STAT1, signal transducer and activator of transcription 1; TIA, transient ischaemic attack.
EMSA analysis using nuclear cell extract from Jurkat, LCL, K562 and PBMCs showed protein binding to the SRP54-AS1 risk locus but no distinct allele specific difference (data not shown). According to chromatin immunoprecipitation with DNA sequencing (ChIP-Seq) data from the ENCODE project, Signal transducer and activator of transcription 1 (STAT1) binds to the SRP54-AS1 locus. As STAT1 is crucial for IFN-α signalling, we tested recombinant STAT1 in the EMSA and showed that STAT1 is one of the proteins binding at the SRP54-AS1 risk locus, with a lower affinity to the risk allele (figure 2B). According to GTEx data, the three SNPs at the SRP54-AS1 locus affect the expression of signal recognition particle 54 (S54), protein phosphatase 2 regulatory subunit B gamma (PPP2R3C), SRP54-antisense 1 (SRP54-AS1) and family with sequence similarity 177 member A1 (FAM177A1) in multiple tissues (online supplementary figure 1A–D).25 These genes are all located close to the SRP54-AS1 risk locus (figure 2A). In addition, the SNP rs712306, in high LD (r2=0.7), with the SRP54-AS1 risk allele showed differential expression of FAM177A1 in the artery wall (P=3.9×10−6) (GTEx, online supplementary figure 1E).25
The IL-19 and the SRP54-AS1 loci in RA and in the general population
In order to investigate if the six identified CVD risk variants were confined to patients with SLE, or could constitute a risk factor for patients with other inflammatory rheumatic diseases, we compared the allele frequencies of the risk gene variants in RA patients with, and without, CVD (table 4). Results show that all 3 IL19 gene variants were associated with stroke/MI in RA and that allele frequencies of the risk gene variants in patients with and without stroke/MI were similar in RA and SLE (table 4). In addition, the IL19 risk alleles showed association with stroke/TIA in RA (OR 2.8 (1.4 to 5.6), P=3.8×10−3). The SRP54-AS1 risk allele did not show an association with CVD in RA (data not shown). When including patients with SLE and RA in a meta-analysis, the association with stroke/MI remained (OR 2.45 (1.96 to 2.94), P=3.5×10−7, after Bonferroni adjustment for 48 000 independent SNPs16 on the Immunochip Padjusted=0.017). Publicly available data of individuals in the general population showed no association between the IL19 and SRP54-AS1 risk variants and ischaemic stroke or coronary artery disease (see supplementary results).
IL19 SNPs associated with stroke/MI in rheumatoid arthritis
Discussion
The objective of the present study was to investigate possible associations between genetic variation and CVD in SLE, and we identified two new risk loci, namely IL19 and SRP54-AS1. The IL19 risk locus is located in an interleukin gene cluster coding for several cytokines, including IL19 and IL10. Several SNPs in this region have previously been associated with SLE27 28 and CVD in both the general population29 30 and in a small SLE study of 52 individuals.31 However, none of these earlier identified SNPs are in high LD with the SNPs at the IL19 risk locus identified here, and none of the previously described SNPs were associated with CVD in our dataset. The observation that our identified risk gene variants were not associated with CVD in the general population suggests that there exist at least partly different underlying mechanisms behind CVD in SLE and the general population. The clinical picture for patients with and without the IL19 risk allele did not differ regarding the ACR criteria18 or the SDI.24 Thus, the higher frequency of strokes and MIs in the group with the risk allele is not caused by a more severe SLE disease. Furthermore, when adjusting for known CVD risk factors, the IL19 risk allele association remained. Thus, our results suggest that the IL19 risk allele is an independent risk factor for stroke and MI in SLE patients with European decent. The observation that the IL19 risk allele also was associated with stroke/MI in patients with RA, but not in the general population, furthermore suggest that the gene variants identified could be confined to patients with an inflammatory rheumatic disease, or at least SLE and RA. However, this assumption needs to be confirmed in studies with patients with other diagnoses.
The IL19 risk locus is a putative binding site for several TFs and is located in a region of open chromatin with predicted enhancer function in LCLs and CD4 and CD8-positive T cells.32 33 In our EMSA analyses, we observed differential binding to the IL19 risk allele of nuclear extract from PBMCs stimulated with IFN-α and activated Jurkat cells. Because extract from unstimulated cells did not show differential binding to the site, cell activation or high levels of IFN-α as observed in SLE seems to be a requirement for the expression of the relevant protein. Publicly available data34 identified IKAROS family zinc finger 3 (IKZF3) as one possible TF with differential binding to the two alleles. IKZF3 is important for regulation of B cell differentiation, and recently a SNP in the promotor region of IKZF3 was shown to be associated with SLE.35 Interestingly, expression of IKZF3 is regulated by IRF8,36 a TF involved in IFN signalling and associated with CVD in SLE.11 37 In conclusion, our EMSA analyses suggest that the IL19 risk locus functions as a cell activation-dependent regulatory region and that the IL19 risk allele abrogates the binding of a TF induced by IFN-α or T cell activation.
To examine if the IL19 risk allele affects gene expression, we used data from the GTEx project25 but found no difference in RNA expression. Next, we went on to analyse protein expression of genes located in the same topologically associated domain38 and measured IL19 and IL10 levels in the patients with SLE. Similar to previously identified SNPs with long-range functional connections,39 we found that the intronic IL19 risk allele affects expression of the IL10 gene, located at a distance of 30 kb from the IL19 locus. As IL10 is known to stimulate B cells and antibody production,40 we measured levels of aPL and indeed found elevated levels of both aCL and aβ2-GPI as well as LA in patients with the IL19 risk allele. As presence of aPLs is a known risk factor for CVD,10 41 especially stroke,12 these results suggest that the IL19 risk locus exerts a regulatory effect on IL10 expression, enhancing production of prothrombotic aPL by B cells. However, as both aPL and the IL19 risk allele remained significantly associated with stroke/MI in the multivariable regression analysis other mechanisms are also possible. Recently, it was reported that IL10 enhances IFN-α-mediated endothelial progenitor cell (EPC) dysfunction and that levels of IL10 correlated with EPC function in SLE but not in healthy controls.42 Thus, there are several mechanisms whereby IL10 could exert a negative effect on the circulatory system in SLE with implications for cardiovascular pathology.
The SRP54-AS1 locus was found to be associated with stroke/TIA in SLE. This locus was not associated with a more severe SLE disease, aPLs or subclinical atherosclerosis suggesting a novel mechanism of action. Published ChIP-Seq data indicate that the SRP54-AS1 locus is located in a region with enhancer/promoter function in CD4, CD8, CD19 and CD14 positive cells with several TFs potentially binding at the locus, including STAT1. In fact, we demonstrated that STAT1 display a lower affinity to the alternative allele. This is interesting given the enhanced expression of STAT1 observed in PBMCs of patients with SLE43 and its role in intracellular signalling following binding of IFN-α to the type I IFN receptor.44 According to the GTEx database, FAM177A1, SRP54, PPP2R3C and SRP54-AS1 are differentially expressed in individuals with the SRP54-AS1 risk allele. Noteworthy, FAM177A1 is differentially expressed in the artery wall and has previously been associated with neurogenetic disorders.45 Because FAM177A1 has unknown functions, further studies are needed to determine possible mechanisms connecting FAM177A1 and stroke/TIA in SLE.
The strength of the present study is the large number of patients with SLE and RA investigated, the detailed information regarding CVD events available and that cytokine levels and aPLs were measured in a large number of patients. Furthermore, the Immunochip has approximately 48 000 independent SNPs when LD is considered, and a conservative p value threshold for significance of meta-analysis would be 1×10−6.16 When including both the patients with SLE and RA in the meta-analysis, the statistical analysis reached this level of significance. Limitations include that the patients in the replication cohort were younger at follow-up resulting in fewer CVD events compared with the discovery cohort and that aPL and stroke data were missing for 20% of patients in the discovery cohort. In addition, we did not have data regarding lipid levels for the patients but identified risk genes have no reported function in lipid metabolism.46
In summary, we have identified two potential risk genes of importance for development of CVD in SLE and for IL19 possibly also in RA. SLE patients with the IL19 risk allele, associated with stroke/MI, more often had high IL10 and elevated levels of aPL. The SRP54-AS1 risk allele, associated with stroke/TIA in SLE, was found to likely affect expression of a number of genes. Strikingly, both identified loci seem to affect gene regulation, rather than protein structure. We could also show that the IL19 gene variant is associated with stroke/MI in both SLE and RA but not in the general population, which suggests the existence of shared mechanisms in the development of CVD in patients with inflammatory rheumatic diseases and this assumption deserves further studies.
Acknowledgments
We would like to thank the patients and healthy volunteers for participating, Lisbeth Fuxler, Rezvan Kiani and Karolina Tandre for technical assistance and collecting blood samples. Genotyping of the discovery and RA cohort was performed by the SNP&SEQ Technology Platform, Uppsala.
References
Footnotes
Handling editor Josef S Smolen
Contributors DL, LR, JKS and AA designed the study. JD, SE, M-LE and DL performed the experiments. KJ-U performed the carotid ultrasound examinations. DL, ES, CB, KET, MF, IG, CS, AAB, SR-D, LAC and LR collected the data. AA, KET, LÄ and DL performed the statistical analysis. DL, AA, KET, JKS, LR, ES, JD, A-CS, LAC, SR-D and M-LE analysed the data. DL, JKS, JD and LR wrote the manuscript. All authors approved the final version of the manuscript.
Funding This project was funded by the Swedish Research Council for Medicine and Health (D0283001, A0258801, E0226301 and E0395401), Knut and Alice Wallenberg Foundation (2011.0073), AstraZeneca-Science for Life Laboratory Research Collaborationgrant (DISSECT), Swedish Society of Medicine and Ingegerd Johansson donation, Swedish Rheumatism Foundation, King Gustaf V’s 80-year Foundation, Torsten Söderberg Foundation, Uppsala County Council and Uppsala University Hospital (ALF), Stockholm County Council (ALF), Uppsala University, Swedish Heart-Lung foundation, Selander Foundation, Agnes and Mac Rudberg Foundation, Gustaf Prim Foundation and COMBINE. Work related to the UCSF cohort was supported by NIH grants UL1-TR-00004, P60-AR-053308 and R01-AR-44804.
Competing interests None declared.
Patient consent Obtained.
Ethics approval The regional ethics committee in Uppsala, Sweden and the local ethics committees.
Provenance and peer review Not commissioned; externally peer reviewed.