Article Text

Abstract
Objectives Systemic sclerosis (SSc) is characterised by tissue fibrosis and vasculopathy with defective angiogenesis. Transforming growth factor beta (TGF-β) plays a major role in tissue fibrosis, including downregulation of caveolin-1 (Cav-1); however, its role in defective angiogenesis is less clear. Pigment epithelium-derived factor (PEDF), a major antiangiogenic factor, is abundantly secreted by SSc fibroblasts. Here, we investigated the effect of TGF-β and Cav-1 on PEDF expression and the role of PEDF in the ability of SSc fibroblasts to modulate angiogenesis.
Methods PEDF and Cav-1 expression in fibroblasts and endothelial cells were evaluated by means of immunohistochemistry on human and mouse skin biopsies. PEDF and Cav-1 were silenced in cultured SSc and control fibroblasts using lentiviral short-hairpin RNAs. Organotypic fibroblast–endothelial cell co-cultures and matrigel assays were employed to assess angiogenesis.
Results PEDF is highly expressed in myofibroblasts and reticular fibroblasts with low Cav-1 expression in SSc skin biopsies, and it is induced by TGF-β in vitro. SSc fibroblasts suppress angiogenesis in an organotypic model. This model is reproduced by silencing Cav-1 in normal dermal fibroblasts. Conversely, silencing PEDF in SSc fibroblasts rescues their antiangiogenic phenotype. Consistently, transgenic mice with TGF-β receptor hyperactivation show lower Cav-1 and higher PEDF expression levels in skin biopsies accompanied by reduced blood vessel density.
Conclusions Our data reveal a new pathway by which TGF-β suppresses angiogenesis in SSc, through decreased fibroblast Cav-1 expression and subsequent PEDF secretion. This pathway may present a promising target for new therapeutic interventions in SSc.
- systemic sclerosis
- fibroblasts
- autoimmune diseases
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Introduction
Systemic sclerosis (SSc) or scleroderma is an autoimmune connective tissue disease characterised by accumulation of extracellular matrix (ECM) proteins within the affected tissues and a widespread vasculopathy comprising both defective angiogenesis and fibroproliferative vasculopathy.1–4 It is well established that transforming growth factor beta (TGF-β) plays a key role in accumulation of collagen and ECM proteins and downregulation of caveolin-1 (reviewed in Del Galdo et al 5and Lafyatis6). Nevertheless, the pathogenesis of sustained microangiopathy and defective angiogenesis, and their causal links with tissue fibrosis are less clearly understood. Recent studies employing a mouse strain with a ligand-dependent upregulation of TGF-β signalling in tissue fibroblasts (TBRIIΔk-fib) demonstrate that increased TGF-β signalling favours the onset of fibroproliferative vasculopathy typical of SSc following minimal endothelial cell injury,7 suggesting that TGF-β activation may also play a role in the pathogenesis of vasculopathy in SSc.
Proteomic studies have previously identified pigment epithelial-derived factor (PEDF) as one of the most abundant secreted proteins by SSc skin fibroblasts compared with healthy controls.8 PEDF is a 46 kDa secreted glycoprotein that belongs to the serpin superfamily but has no protease inhibitory function. Despite the lack of protease activity, PEDF exerts diverse physiological functions including antiangiogenesis,9 antivasopermeability10 and neurotrophic activities.11 12 PEDF is expressed abundantly in pigmented epithelium of the cornea where it plays a crucial role in the suppression of angiogenesis.9 13–16
Previous studies have shown that PEDF is highly expressed in idiopathic pulmonary fibrosis and is inducible by TGF-β in cultured human lung fibroblasts, suggesting a potential link with the pathogenesis of tissue fibrosis.17 Here, we set to determine PEDF expression in SSc skin biopsies and in TBRIIΔk-fib mice in vivo and to determine the role of PEDF expression in the antiangiogenic function of fibroblasts in vitro.
Materials and methods
Patient samples and patient skin biopsies
Skin biopsies from nine patients with early diffuse cutaneous systemic sclerosis (dcSSc)18 and nine healthy controls were obtained at the SSc clinic within the Leeds Institute of Rheumatic and Musculoskeletal Medicine (UK) and the Rheumatology Unit in L’Aquila (Italy). Biopsies were taken with full informed consent as approved by National Research Ethics Service (NRES) Committee (REC 10/H1306/88) and the local ethical committee in University of L’Aquila, and processed as described in detail in online supplementary methods.
Supplementary file 1
Cell culture
Dermal fibroblasts (FBs) from patients with dcSSc and controls were isolated as previously described19 and detailed in online supplementary methods. Human dermal microvascular endothelial cells (MVECs) were purchased from PromoCell, UK, cultured in Endothelial Cell Growth Medium BulletKit (Lonza, Slough, UK) and used at passages 2–4. Human umbilical vein endothelial cells (HUVECs) were obtained from TCS Cellworks, cultured in fully supplemented human large vessel endothelial cell medium (TCS Cellworks, UK) and used at passages 3–4. Human epidermal melanocytes (HEMs) were purchased from ScienceCell, cultured in melanocyte medium supplemented with melanocyte growth supplement (TCS Cellworks) and used at passages 3–4. All cells were kept at 37°C in a humidified atmosphere of 5% CO2.
hTERT immortalisation and transduction with lentiviral short-hairpin RNA
For immortalisation of primary FBs, pBabe human telomerase reverse transcriptase (hTERT) puromycin retrovirus was employed following standard protocols (described in detail in online supplementary methods). Cav-1 and PEDF expression were silenced by transduction with shRNAmir GIPZ lentiviruses (Open Biosystems, Surrey, UK) following manufacturer’s instructions (online supplementary methods).
Culture treatments
To evaluate the effect of TGF-β on PEDF and COLA1, cells were grown to confluence in six-well culture plates in dulbecco modified eagle medium (DMEM) 10% fetal calf serum (FCS), serum starved in DMEM 0.5% FCS for 24 hours and stimulated in the presence of 10 ng/mL recombinant human (rh) TGF-β1 (Sigma, USA) for 48 hours. Ascorbic acid (40 µg/mL) was used to optimise collagen production.20
RNA isolation and RT-PCR analysis
Total RNA was isolated using the RNeasy Mini Kit (Qiagen, USA) according to the manufacturer’s instructions. One microgram of total RNA from each sample was retrotranscribed to first-strand cDNA using the SuperScript III One Step RT-PCR system (Invitrogen, UK). Quantitative RT-PCR was performed in triplicates using SYBR Green RT-PCR Master Mix Kit and the ABI PRISM 7500 Fast Real Time PCR System (Applied Biosystems) with the following primers: PEDF 5′-TGTCTCCAACTTCGGCTATG-3′ (forward) and 5′-AGTAGAGAGCCCGGTGAATG-3′ (reverse), Cav-1 5′-CGACCCTAAACACCTCAACGA-3′ (forward) and 5′-TCCCTTCTGGTTCTGTCA-3′ (reverse). Quantification was performed using the comparative CT (cycle-threshold) method employing ribosomal 18S as a housekeeping gene.
Protein quantification and immunodepletion
Secreted PEDF and Collagen I were detected by immunoblotting in cultured cells and supernatants and by ELISA in patient sera as described in online supplementary methods. Immunodepletion for PEDF was conducted using monoclonal mouse anti-PEDF (Chemicon-Millipore, Clone 10F12.2). Full experimental procedures are described in detail in online supplementary methods.
Angiogenesis assays
The organotypic co-culture assay21–23 was performed with MVECs or HUVEC and either primary or hTERT immortalised fibroblasts stably infected with lentivirus coding for either non-silencing control, Cav-1 or PEDF short-hairpin (sh) RNAs as described in detail in online supplementary methods. Number of tubules and total tubule length were analysed using the Angiosys software (TCS Cellworks).
Reduced growth factor matrigel (VWR, UK) was used to perform angiogenesis matrigel assays in HUVEC employing supernatants from cultured FBs (online supplementary methods).
FACS analysis and proliferation assay
For PEDF detection using FACS analysis, fibroblasts were incubated in the presence of a protein transport inhibitor GolgiPlug (BD Biosciences) for 12 hours according to manufacturer’s instructions. The cells were then stained on ice with rabbit polyclonal anti-PEDF-PECy5.5 (Bioss) following fixation and permeabilisation, and analysed against the corresponding isotype control using BD FACSDiva software V.6.0 BDTM LSR II flow cytometer.
For determination of HUVEC proliferation, cells were labelled with the carbocyfluorescein succinimidyl ester dye analogue, CellTrace Violet (Invitrogen). Prior to co-culture experiments, in order to track cell division following co-culture and for accurate gating, CD90-PEvio770 and CD31-APC (Miltenyi biotec) were used to exclude potential contamination of HUVEC with co-cultured fibroblasts; 7-aminoactinomycin D was used as a viability marker. Cell division frequency and proliferation indices from list mode data were determined using proliferation wizard of ModFit software V.3.2 (Verity Software House, Topsham, ME, USA).
Immunohistochemistry
Immunohistochemistry (IHC) analysis of human skin biopsies was performed on 3 µm paraffin sections using mouse monoclonal anti-PEDF (Clone 10F12.2; Millipore, UK), rabbit polyclonal anti-CD31, rabbit polyclonal anti-α-smooth muscle actin (SMA) (Abcam, UK) and a rabbit polyclonal anti-Cav-1 antibody (Santa Cruz, UK). Detailed procedure of the two-step staining is described in the online supplementary methods. The number of positive cells was counted by two pathologists, blinded to tissue source and expressed as the mean of two observations for each sample. For mouse skin biopsies, we employed rabbit polyclonal anti-PEDF (Aviva Systems Biology; Insight Biotech, UK), rabbit polyclonal anti-CD31 (Santa Cruz) and rabbit polyclonal anti-Cav-1 (Santa Cruz) antibodies. All sections were imaged using an Axioplan Zeiss light microscope equipped with an AxioCam digital camera.
TβRIIΔk-fib animal model
TβRIIΔk-fib transgenic mice were provided by Professor C. Denton of UCL Medical School Centre for Rheumatology and Connective Tissue Diseases, London, UK. The generation of TβRIIΔk-fib transgenic (TG) mice has been described previously.7 Mice were genotyped by PCR using LacZ primers and an internal control. Experiments were performed on three transgenic mice aged 6 weeks and compared with sex-matched littermate controls. Animals were housed in a conventional clean facility, with access to food and water ad libitum. Strict adherence to institutional guidelines was practised, under full local ethics committee and Home Office approval.
Results
PEDF expression is increased in SSc patient biopsies compared with healthy donors
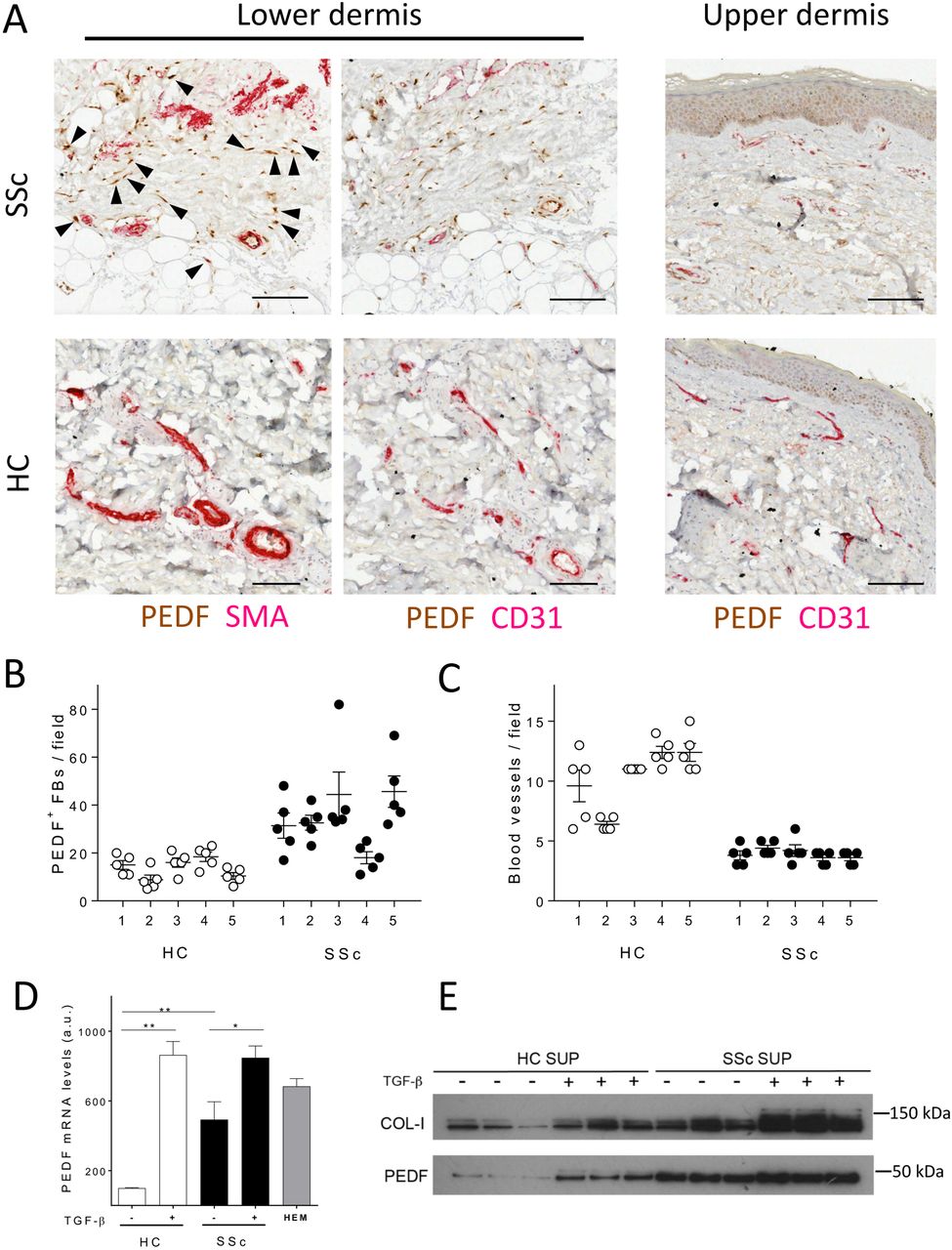
In healthy skin, PEDF is expressed mainly in the germinal layer of the epidermis (figure 1A). In contrast, in early diffuse SSc, we observed a strong staining for PEDF within the lower reticular dermis, both in fibroblasts and around blood vessels (figure 1A). Double IHC studies revealed that strong positivity of PEDF correlated with strong positivity of α-SMA (figure 1A and online supplementary figure 1A, B), indicating increased number of myofibroblasts positive for PEDF in SSc biopsies. Interestingly, we also observed positivity for PEDF in vascular endothelial cells identified by CD31 staining as well as in perivascular cells (figure 1A). Quantification of PEDF-positive dermal fibroblasts within the reticular connective tissue showed significantly higher PEDF positivity in SSc fibroblasts compared with healthy control fibroblasts (figure 1B). This was accompanied by the already described decreased abundance of blood vessels in the SSc biopsies compared with healthy controls (figure 1C). Consistent with these findings, SSc skin biopsies showed 5.5-fold increase in PEDF mRNA expression as assessed by qRT-PCR (P<0.01, n=5, online supplementary figure 1D). Following this observation, we have measured PEDF concentration in 38 patients with dcSSc and 34 healthy controls from our observational cohort but found no statistically significant difference in serum levels of PEDF (P=0.87).
Supplementary file 2
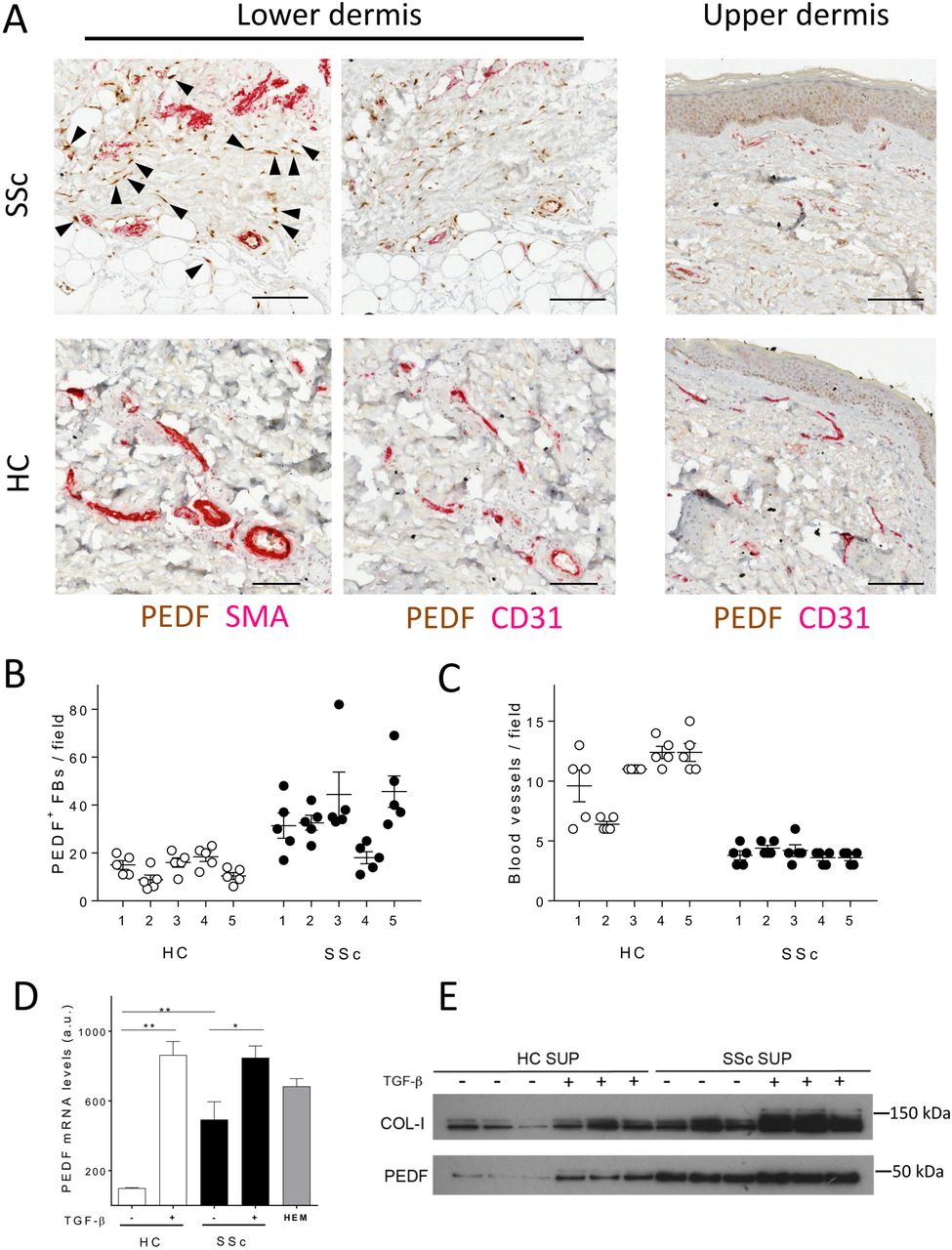
Pigment epithelium-derived factor (PEDF) expression is increased in systemic sclerosis (SSc) skin in vivo and it is inducible by transforming growth factor beta (TGF-β) in vitro. (A) Images depict representative forearm skin biopsies from healthy controls (HC) and patients with SSc (SSc) double stained for PEDF and α-smooth muscle actin (SMA), or PEDF and CD31 as indicated. Arrowheads point to spindle-shaped PEDF and SMA-positive cells within the dermis. For a larger field of stained skin section, see online supplementary figure 1A, B. Scale bars, 100 µm. (B) Dot blots show counts of PEDF-positive fibroblasts in HC and SSc samples; bars represent mean values±SEM (n=25 microscopic fields per biopsy from five different biopsies). (C) Dot blots show blood vessel counts in HC and SSc samples; bars represent average values±SEM (n=25 microscopic fields per biopsy from five different biopsies). Data on additional patient samples are shown in online supplementary table 1. (D) Histogram shows PEDF mRNA levels by RT-PCR in isolated HC fibroblasts (HC-FBs) and SSc fibroblasts (SSc-FBs); bars represent mean values±SEM (n=9 samples from each HC and SSc). Where indicated, cells were treated with 10 ng/mL TGF-β for 24 hours; HEM, human epithelial melanocytes positive control. *P<0.05, **P<0.01 by unpaired t-test. (E) Western blot shows PEDF in supernatants (SUP) collected from HC-FBs and SSc-FBs cultured in the presence or absence of TGF-β (10 ng/mL for 48 hours).
Supplementary file 3
PEDF expression in SSc and healthy fibroblasts is induced by TGF-β
To determine whether the increase in PEDF expression resulted from upregulation in gene expression, we quantified PEDF mRNA levels by RT-PCR analysis in primary dermal FBs. FBs from patients with SSc expressed approximately 5-fold higher PEDF mRNA levels compared with HC-FBs (figure 1D). Concordantly, there was over 2-fold higher levels of intracellular PEDF protein in SSc-FBs compared with HC-FBs, as assessed by FACS mean fluorescence intensity for PEDF (online supplementary figure 1C). In order to establish whether the upregulation of PEDF expression in SSc-FBs resulted from increased TGF-β signalling, we performed RT-PCR on healthy control and patient isolated fibroblasts following 48 hours of stimulation with hrTGF-β. hrTGF-β induced PEDF expression in HC-FBs by 9-fold compared with 1.6-fold increase in SSc fibroblasts (figure 1D). The levels of PEDF mRNA expression following TGF-β stimulation were comparable in HC and SSc fibroblasts. Concordantly, mean fluorescent intensity by FACS for intracellular PEDF was also increased by 2.4-fold following TGF-β stimulation (online supplementary figure 1C). Western blot analysis of supernatants from fibroblast cultures confirmed that rhTGF-β treatment increased the levels of secreted PEDF, and this was associated with an increased secretion of collagen 1 (figure 1E) as shown previously.24
SSc fibroblasts suppress angiogenesis in a PEDF-dependent manner
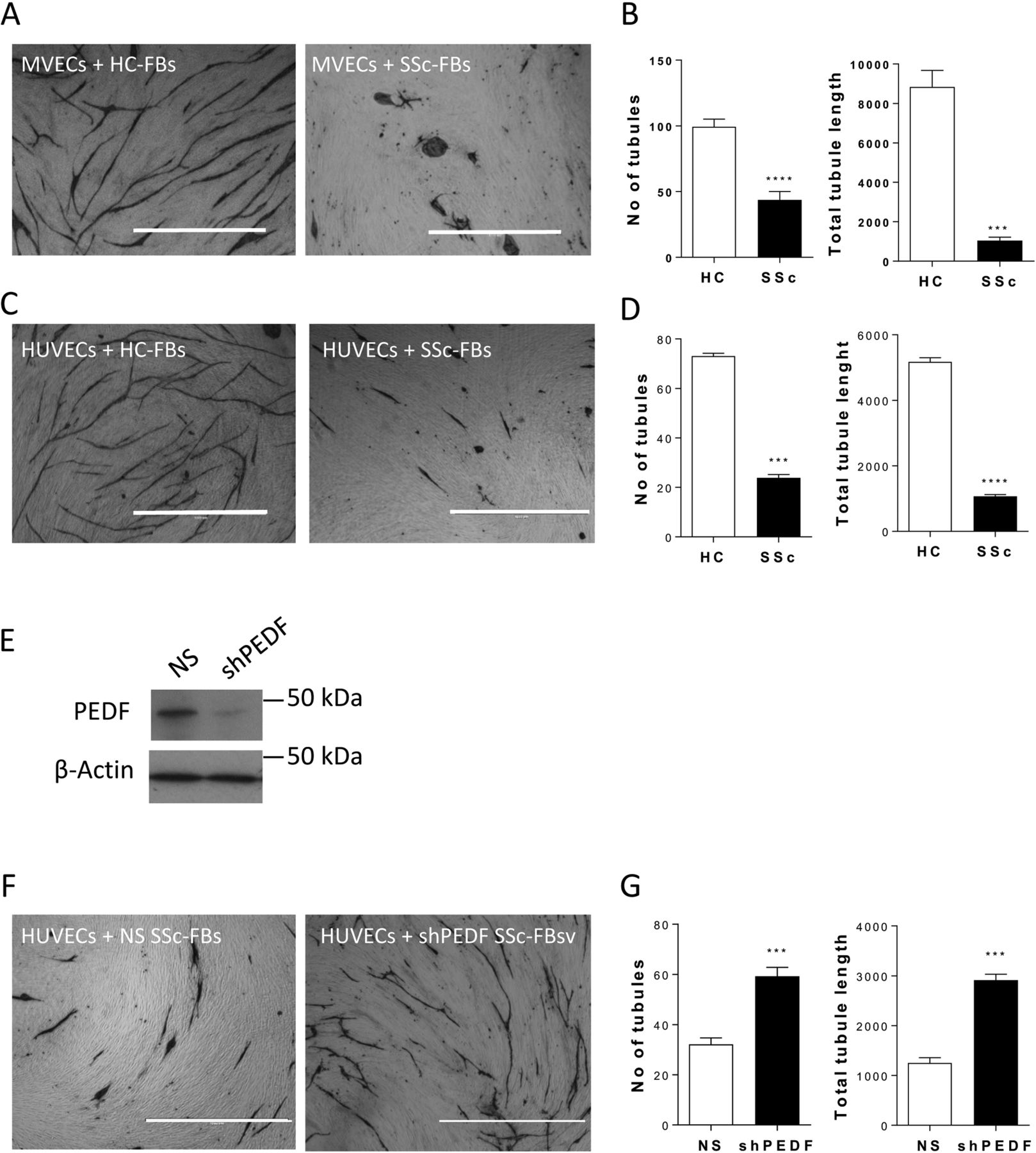
To determine whether the observed increased expression of PEDF by dermal fibroblasts in SSc skin biopsies could contribute to defective angiogenesis, we performed endothelial-fibroblast organotypic angiogenesis in vitro assays in which endothelial cells form tubules highly reminiscent of capillaries formed during angiogenesis in vivo, embedded in natural matrix produced by the fibroblasts.21–23 Use of rhPEDF in this assay confirmed in vitro the known antiangiogenic effect of PEDF (online supplementary figure 2). In the same model, co-culture of MVECs with SSc-FBS showed a 60% decrease in the number of tubules and 90% decrease in total tubule length compared with HC-FBs (figure 2A, B). Similar results were obtained when HUVECs were co-cultured with HC-FBs or SSc-FBs (figure 2C, D), hence HUVECs were employed in subsequent assays. Knockdown of PEDF in SSc-FBs by means of lentiviral shRNA (figure 2E), followed by co-culture with ECs, rescued the number of tubules and total tubule length by 1.9-fold and 2.5-fold, respectively, compared with SScFBs infected with control lentivirus harbouring non-silencing shRNA (figure 2F, G).
Supplementary file 4

Suppression of angiogenesis in an organotypic co-culture assay by systemic sclerosis (SSc) fibroblasts is reversed by pigment epithelium-derived factor (PEDF) knockdown. (A, C) Images show representative microscopic fields from co-culture assays of human dermal microvascular endothelial cells (MVECs) (A) or human umbilical vein endothelial cells (HUVECs) (C) seeded onto confluent fibroblasts (FBs), healthy control (HC-FBs) or SSc (SSc-FBs), stained for the endothelial marker CD31 (fibroblasts are seen unstained in the background). Note that HUVECs reproduce the behaviour of MVECs in the organotypic assays. (B, D) Histograms show the number of tubules and total tubule length quantified using Angiosys software, represented as mean±SEM (n=12 microscopic fields at ×4 magnification from triplicate wells). (E) Representative western blot showing intracellular PEDF levels in SSc fibroblasts treated with GolgiPlug, non-silencing control (NS) or with PEDF depletion (shPEDF) by means of lentiviral short-hairpin RNA (sh). (F) Images show representative microscopic fields from co-culture assays of HUVECs seeded onto confluent SSc fibroblasts (SSc-FBs), non-silencing control (NS) or with PEDF depletion (shPEDF). (G) Quantification of the number of tubules and total tubule length represented as mean±SEM (n=12 microscopic fields at ×4 magnification from triplicate wells). **P<0.01, ***P<0.001 by unpaired t-test. Scale bars, 100 µm.
Downregulation of caveolin-1 induces PEDF expression in dermal fibroblasts
TGF-β downregulates caveolin-1 expression in vitro, both at the RNA and protein level,25 26 and this has been associated with myofibroblasts’ profibrotic activation.27 28 Double IHC studies showed an abundance of caveolin-1-positive cells largely negative for PEDF in HC skin, whereas in SSc skin biopsies, the majority of fibroblasts were PEDF positive and caveolin-1 negative (figure 3A, B). Moreover, there was an overall reduction of caveolin-1 positivity in SSc skin biopsies (figure 3A–C) as previously shown.18 These results suggested an inverse correlation between PEDF and caveolin-1 expression. The increased secretion of PEDF and reduced expression of caveolin-1 was conserved in vitro. SSc fibroblasts showed increased PEDF protein levels (online supplementary figure 1) and decreased expression of caveolin-1 (figure 3D). Interestingly, the relative expression of caveolin-1 in subcultured fibroblasts (SSc 1–5) followed the same trend observed by IHC in patients 1–5 (online supplementary table 1). Stable silencing of caveolin-1 by lentiviral delivery of caveolin-1 shRNA showed 85% caveolin-1 knockdown compared with control (figure 4A). Fibroblasts with caveolin-1 knockdown displayed over 2-fold upregulation of PEDF expression compared with controls (figure 4B). Accordingly, we observed an increased secretion of PEDF in the supernatants harvested from fibroblasts with caveolin-1 knockdown compared with control (figure 4C). These results show that decreased caveolin-1 expression in dermal fibroblasts induces the expression and secretion of PEDF in vitro and that the inverse correlation of expression is observable in vivo.

Decreased caveolin-1 tissue expression in systemic sclerosis (SSc) is conserved in vitro and correlates with high pigment epithelium-derived factor (PEDF) expression. (A) Images show representative forearm skin biopsies from healthy controls (HC) and patients with SSc double stained for PEDF and caveolin-1 (Cav-1). Arrowheads point to Cav-1-positive cells; note that Cav-1-positive cells show no PEDF positivity. Scale bars, 50 µm. (B) Dot plots show quantification of PEDF-positive and Cav-1-negative fibroblasts (FBs) from HC and patients with SSc. (C) Dot plots show quantification of Cav-1-positive FBs in HC and patients with SSc. Data on additional patient samples are shown in online supplementary table 1. (D) Western blots of five HC and SSc FB cultures for Cav-1 and glyceraldehyde 3-phosphate dehydrogenase (GAPDH). Dermal fibroblasts were subcultured from the same biopsies analysed by immunohistochemistry and loaded in the same order as shown in panels (B) and (C) and summarised in online supplementary table 1.

Caveolin-1 (Cav-1) knockdown stimulates pigment epithelium-derived factor (PEDF) expression in dermal fibroblasts and suppresses tubulogenesis without affecting endothelial cell proliferation. (A–C) Cav-1 knockdown stimulates PEDF expression. (A) Histogram depicts caveolin-1 knockdown (shCav-1) in dermal fibroblasts by means of lentiviral short-hairpin RNA (sh), as percentage non-silencing control (NS) by RT-PCR. Bars represent mean values±SD (n=3 independent experiments carried out in triplicates). ***P<0.001 by unpaired t-test. (B) Histogram depicts PEDF mRNA by RT-PCR with caveolin-1 knockdown (shCav-1) compared with non-silencing control (NS). Bars represent mean values±SD (n=3 independent experiments carried out in triplicates). **P<0.01 by unpaired t-test. (C) Representative western blot shows levels of PEDF in supernatants (SUP) from fibroblasts with shCav-1 compared with NS control. (D) Images show representative microscopic fields from co-culture assays of human umbilical vein endothelial cells (HUVECs) seeded onto confluent systemic sclerosis fibroblasts (SSc-FBs), non-silencing control (NS) or with PEDF depletion (shPEDF) stained by CD31. Scale bars, 100 µm. Note the decreased tubule formation with caveolin-1 knockdown in fibroblasts. (E) Histograms show number of tubules and total tubule length in (D) quantified using Angiosys software, represented as mean±SEM (n=12 microscopic fields from three different experiments). (F) Representative images from HUVEC matrigel assays with cultures treated with supernatants from dermal fibroblasts, non-silencing control (NS) or with caveolin-1 knockdown (shCav-1). Note the HUVEC monolayer organisation and reduced number of loops in shCav-1 supernatant-treated cultures compared with control. (G) Quantification of number of loops in (F) represented as mean±SD (n=12 HPF from three different experiments). (H) Histograms show cell generation of HUVEC preloaded with CellTrace Violet co-cultured with caveolin-1 knockdown (shCav-1) fibroblasts or non-silencing control fibroblasts (NS); PI, relative proliferation indexes. Dye dilution analysis performed using Modfit proliferation algorithm. (I) Column chart showing quantification of HUVEC generations while in co-culture with caveolin-1 knockdown (shCav-1) fibroblasts, or non-silencing control fibroblasts (NS); chart indicates some increase in proliferation at G5/G6 in caveolin-1 knockdown fibroblast co-cultures (shCav-1) compared with non-silencing control fibroblasts (NS). **P<0.01; ***P<0.001 by unpaired t-test. HPF, high power fields.
Caveolin-1 downregulation in fibroblasts inhibits angiogenesis without affecting endothelial viability and proliferation
Fibroblasts with lentiviral shRNA driven caveolin-1 knockdown suppressed tubule formation in the organotypic angiogenesis assay, with a 62.5% decrease in tubule length and 58.2% decrease in total number of tubules (P<0.001 for both) (figure 4D) compared with controls. Accordingly, matrigel assays using supernatants harvested from the same cells showed nearly 50% reduction in tube formation (P<0.01) (figure 4F, G). Therefore, the inhibitory effect on tubulogenesis of caveolin-1 knockdown in fibroblasts is transferable by tissue culture supernatants, supporting the notion that it is mediated via secreted factors such as PEDF. Consistently, immunodepletion of PEDF from fibroblast supernatants, through treatment with a blocking antibody (online supplementary figure 3A), rescued tubule morphogenesis significantly in the matrigel assay (online supplementary figure 3B, C).
Supplementary file 5
Viability of endothelial cells (CD31pos/CD90neg) in the organotypic angiogenesis assay was comparable between the two experimental conditions (online supplementary figure 3D). Similarly, the proliferation index by the dye dilution method of endothelial cells was comparable in the two conditions (figure 4H, I). These data indicate that the impairment in angiogenesis, mediated by fibroblasts with caveolin-1 knockdown, does not result from decreased endothelial cell viability or proliferation.
TGF-β signalling suppresses caveolin-1 expression and stimulates PEDF expression in vivo
To investigate whether the findings from the patient sample analysis and culture systems hold in an in vivo model of overactivation of TGF-β signalling, we employed mice with controlled overexpression of TGF-β receptors in fibroblasts driven by the col1a1 promoter (TβRIIΔk-fib). Analysis of sections from mouse skin biopsies showed reduced caveolin-1 expression in TβRIIΔk-fib transgenic mice compared with wild-type littermate controls (figure 5A). Conversely, expression of PEDF was increased in the skin of the transgenic mice (figure 5B). Importantly, increased PEDF and reduced caveolin-1 expression in TβRIIΔk-fib transgenic mice was associated with a reduction in the number of capillaries as assessed by IHC for CD31 (figure 5C).

Skin immunohistochemistry of transgenic mice (TβRIIΔk-fib) and wild-type (WT) mice. Images show representative mouse skin biopsies stained for caveolin-1 (Cav-1) (A), pigment epithelium-derived factor (PEDF) (B) or CD31 (C) from transgenic TβRIIΔk-fib mice and wild-type littermate controls. Note the decreased expression of caveolin-1 and increased expression of PEDF in TβRIIΔk-fib biopsies. Scale bar, 50 µm, original magnification ×20. Plots show quantification of caveolin-1-positive (A) and PEDF-positive (B) fibroblasts (FBs), and blood vessel counts (C). Bars represent average values±SD (n=9 biopsies from three mice per genotype). *P<0.05; **P<0.01; ***P<0.001 by unpaired t-test.
Discussion
Here, we demonstrate for the first time that dermal fibroblasts in SSc play a direct role in the impairment of angiogenesis via secretion of PEDF. The increased PEDF expression in dcSSc skin biopsies validates the proteomics data of dcSSc fibroblast secretome,8 and it is consistent with the increased PEDF expression observed in IPF by Cosgrove et al.17 The PEDF-positive cells were both tissue fibroblasts (for their classic spindle-shape morphology) and myofibroblasts (SMA positive). In addition, we observed high PEDF expression in cells with perivascular localisation, suggesting that in vivo, multiple cell types may contribute to aberrant secretion of PEDF in SSc. Nevertheless, we did not find an increased PEDF serum concentration in patients with dcSSc versus healthy controls, consistent with the known paracrine mode of action of PEDF.
Interestingly, PEDF was found significantly more abundant in the lower dermis of patients with dcSSc, suggesting that reticular fibroblasts may play an important role in the initiation and progression of impaired angiogenesis.
Here, we show for the first time that TGF-β signalling in fibroblasts suppresses angiogenesis through secretion of PEDF, and that this pathway remains active in fibroblasts explanted from SSc skin. This is consistent with a wealth of data in the literature indicating that SSc fibroblasts maintain in vitro hallmarks of TGF-β signalling activation with passaging, including increased collagen production, phosphorylation of SMAD, Jnk and ERK as well as increased expression of α-SMA.
Another big set of evidence indicates that PEDF mediates its antiangiogenic effects through multiple mechanisms including suppression of migration via p38 signalling, induction of apoptosis through MEK5/Erk5 signalling to peroxisome proliferator-activated receptor gamma and NF-κB,9 29 30 and antagonism of vascular endothelial growth factor (VEGF) signalling via γ-secretase cleavage of VEGF receptors.13 31
Therefore, our data link two widely demonstrated molecular pathways and suggest an explanation of TGF-β-induced vasculopathy in SSc.
Intriguingly, PEDF has been found to have an antifibrotic effect in a chemically induced model of liver fibrosis.32 33 Although we cannot exclude that the TGF-β-induced expression of PEDF observed both in SSc8 34 and idiopathic pulmonary fibrosis17 could represent an attempt to negatively feedback the fibrotic process, here we show that PEDF expression can ultimately contribute to the defective angiogenesis in a paracrine manner during SSc.
Previously, we and others reported that SSc skin and lung biopsies show a decreased expression of caveolin-1 when compared with healthy controls, which is associated with tissue fibrosis.18 25–28
Caveolin-1 plays a bidirectional role in endothelial cells during angiogenesis (reviewed in Sowa35) by promoting36 37 or inhibiting38 blood vessel formation; however, the indirect effects on angiogenesis of low caveolin-1 levels in tissue fibroblasts were never studied before. The current study shows that similar to previous data on α-SMA and collagen, also PEDF increased expression in vivo is conserved in subcultured dermal fibroblasts consistent with the known sustained TGF-β signalling of SSc fibroblasts in vitro. Most importantly, we show that the inverse correlation in the expression of caveolin-1 and PEDF is associated with the decreased number of capillaries observed in SSc. Further, we show a causative link between caveolin-1 decreased expression and PEDF secretion, which in turn suppresses angiogenesis in vitro, without affecting cell viability or proliferation. Altogether, the rescue experiments we have performed using PEDF knockdown in SSc fibroblasts and PEDF blocking antibody in SSc supernatants (figure 2F, G and online supplementary figure 3) clearly show the importance of PEDF in the antiangiogenic phenotype of these cells.
Further, here we show that the inverse relationship between caveolin-1 and PEDF expression observed in the patient samples is recapitulated in the TβRIIΔk-fib transgenic mice with overactivation of TGF-β signalling in tissue fibroblasts.7 Consistent with this observation and the antiangiogenic function of PEDF, we also noted a reduced capillary density in the TβRIIΔk-fib mouse skin biopsies. Functional experiments will be necessary to determine whether this transgenic line can be used as preclinical model of TGF-β-induced vasculopathy and targeting of this signalling axis in SSc.
Overall, the findings in this study strongly support the notion that TGF-β is involved in the pathogenesis of vasculopathy in SSc, and establish a causative link between caveolin-1 downregulation in tissue fibroblasts, PEDF expression and defective angiogenesis (figure 6). While this study focuses on dcSSc, in follow-up studies it would be interesting to investigate the relationship between PEDF, caveolin-1 and angiogenesis in limited cutaneous SSc (lcSSc). It is worth noting that lcSSc skin biopsies do not show the hallmarks of TGF-β activation seen in dcSSc,39 and therefore it would be very interesting to investigate whether the vasculopathy observed in lcSSc is driven by a non-TGF-β-related pathway.

{kind=link}
![[SP4.jpg]](https://ard.bmj.com/content/annrheumdis/77/3/431/DC4/embed/inline-supplementary-material-4.jpg?download=true){kind=link}
{kind=link}
{kind=link}
{kind=link}
![[SP5.jpg]](https://ard.bmj.com/content/annrheumdis/77/3/431/DC5/embed/inline-supplementary-material-5.jpg?download=true){kind=link}
{kind=link}
{kind=link}
Mechanistic model depicting the relationship between caveolin-1 (Cav-1) and pigment epithelium-derived factor (PEDF). Transforming growth factor beta (TGF-β) signalling strength is autoregulated by Cav-1-dependent TGF-β receptor internalisation.40 Caveolin-1 downregulation and potentiation of TGF-β signalling promotes PEDF transcription, expression and secretion by dermal fibroblasts, suppressing angiogenesis in systemic sclerosis (SSc). Exposure to high levels of TGF-β and receptor overactivation in SSc sustains caveolin-1 downregulation at the transcriptional level,27 thus promoting further PEDF expression and impairment of angiogenesis.
References
Footnotes
VL and JE contributed equally.
Handling editor Tore K Kvien
Contributors VL, JEC, YME-S, MS, GG, ECD-S and FE contributed to the experimental data collection and analysis. GA and PC contributed to the identification and collection of patient clinical data and biosamples. PE, CPD, RG and GM contributed to data revision, manuscript revision and discussion. VL and JEC drafted the manuscript. FDG ideated the experimental plan and supervised all aspects of research. FDG and GM critically reviewed data and their analysis.
Funding This study was partially funded by NIHR CDF to FDG and EULAR ODP grant to FDG.
Competing interests None declared.
Ethics approval The study was approved by the NHS REC 10/H1306/88 and Institutional Review Board of University of L’Aquila.
Provenance and peer review Not commissioned; externally peer reviewed.
Data sharing statement There are no additional or unpublished data that need to be shared as part of this study.