Article Text

Abstract
Objectives To explore the role of Vδ2 T cells in the pathogenesis of rheumatoid arthritis (RA).
Methods Sixty-eight patients with RA, 21 patients with osteoarthritis and 21 healthy controls were enrolled in the study. All patients with RA fulfilled the 2010 American College of Rheumatology/European League Against Rheumatism criteria for RA. Peripheral Vδ2T population, chemokine receptor expression and proinflammatory cytokine secretion were quantified by flow cytometry. The infiltration of Vδ2 T cells within the synovium was examined by immunohistochemistry and flow cytometry. The effect of tumour necrosis factor (TNF)-α and interleukin (IL)-6 on Vδ2 T migration was determined by flow cytometry and transwell migration assay.
Results Peripheral Vδ2T cells, but not Vδ1 T cells, were significantly lower in patients with RA, which was negatively correlated with disease activity gauged by Disease Activity Score in 28 joints. Vδ2 T cells from RA accumulated in the synovium and produced high levels of proinflammatory cytokines including interferon-γ and IL-17. Phenotypically, Vδ2 T cells from RA showed elevated chemotaxis potential and expressed high levels of chemokine receptors CCR5 and CXCR3, which was driven by increased serum TNF-α through nuclear factor kappa B signalling. In vivo, TNF-α neutralising therapy dramatically downregulated CCR5 and CXCR3 on Vδ2 T cells and repopulated the peripheral Vδ2 T cells in patients with RA.
Conclusions High levels of TNF-α promoted CCR5 and CXCR3 expression in Vδ2 T cells from RA, which potentially infiltrated into the synovium and played crucial roles in the pathogenesis of RA. Targeting Vδ2 T cells might be a potential approach for RA.
- rheumatoid arthritis
- anti-TNF
- T cells
This is an Open Access article distributed in accordance with the Creative Commons Attribution Non Commercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: http://creativecommons.org/licenses/by-nc/4.0/
Statistics from Altmetric.com
Introduction
γδ T cells are a subset of T cell with distinctive T cell receptor (TCR), which is composed of one γ chain and one δ chain. γδ T cells mainly accumulate in mucosal tissues such as gut, and consist the minor population of peripheral lymphocytes (2%–5%).1 2 Two main subsets of γδ T cells have been defined, namely Vγ9/Vδ2 and Vδ1. Vγ9/Vδ2 T cells, the major population of peripheral blood γδ T cells, express TCR variable regions Vγ9 and Vδ2 and produce high levels of interferon (IFN)-γ and tumour necrosis factor (TNF)-α. They also participate in host defence against intracellular pathogens and haematological malignancies.3–6 Vδ1 T cells, which differ from Vγ9/Vδ2 T cells, mainly reside in the mucosal-associated lymphoid tissues and consist approximately 10%–30% of peripheral γδ T cells.7 Vδ1 cells have lower cytotoxicity compared with Vδ2 T cells, have regulatory potential8 and produce a broader set of cytokines, including interleukin (IL)-4 and IL-17.3 9
γδ T cells are implicated in many infectious diseases and tumours. In recent years, growing evidence has implicated γδ T cells in human autoimmune disorders such as diabetes, arthritis and multiple sclerosis. Vδ2-expressing circulatory γδ T cells significantly accumulated in the brains of patients with multiple sclerosis.10 In addition, γδ T cells were found to induce Ig secretion in B cell lines and induce autoantibody production in peripheral B cells from patients with systemic lupus erythematosus.11
Rheumatoid arthritis (RA) is a chronic inflammatory disease that causes severe joint destruction and deformity. RA is characterised with serum autoantibodies as well as extensive lymphocytes infiltration in the synovia, including T and B cells. CD4+ T cells play crucial roles in the pathogenesis of RA. However, accumulating evidence suggests that γδ T cells are also involved in RA.12 13 In collagen-induced arthritis, an experimental model of RA, preventive depletion of γδ T cells ameliorated the disease severity in DBA1/J mice.12 In human RA, synovial effusions (SF) and synovial membranes have been found to contain a high number of T cells bearing the γδ TCR.14 15 Their percentages in SFs were between twofold and fourfold higher compared with peripheral blood.
In light of this evidence, we investigated Vδ2 T cells in peripheral blood, SF and synovium from patients with RA and their contribution to the pathogenesis of RA.
Methods
Patients and controls
This study was approved by the Institutional Review Board of Peking Union Medical College Hospital. Written informed consent was obtained from each participating patient and healthy control (HC). Peripheral blood samples were collected from 15 patients with RA before and after treatment with TNF-α antagonist or IL-6 receptor antagonist.
Detection of phosphorylation
Peripheral blood mononuclear cells (PBMCs) were maintained for 24 hours in RPMI 1640 medium containing 0.1% serum. To activate cytokine-induced signalling, PBMCs were treated in RPMI 1640 containing TNF-α (100 ng/mL) for 5–30 min at 37°C. Then the cells were fixed and permeabilised according to the instructions of BD Phosflow Protocol. The treated and untreated cells were stained with antibody for 1 hour at room temperature. Stained cells were acquired by flow cytometry on a BD Accuri C6 flow cytometer (Becton Dickinson) and analysed using the FlowJo software (Tree Star).
Statistics
All data were analysed using SPSS V.17.0 software. One-way analysis of variance (ANOVA) with Tukey-Kramer post-hoc test was used to compare data displaying a normal distribution and homogeneity of variance. Two-way ANOVA was used to examine the influence of two different categorical independent variables. Student’s t-test was used to compare differences between two groups, and paired t-test was used to compare differences before and after treatment. Correlations were calculated using Spearman correlation analysis.
Other experimental procedures were included in the ‘online supplementary file’.
Supplementary file 1
Results
Peripheral blood Vδ2 T cells were lower in patients with RA and negatively correlated with disease activity
To systematically investigate the roles of γδ T cells in the pathogenesis of RA, we first compared the subpopulations of peripheral γδ T cells in RA, patients with osteoarthritis (OA) and HCs. The results showed a significant decrease of peripheral total γδ T cells in RA (3.45%±0.48% vs HC 8.35±1.00% vs OA 7.21%±0.77%; p<0.01) (figure 1A), which resulted from significant reduction of peripheral Vδ2 T cells (1.80%±0.32% vs HC 5.68±0.70% vs OA 4.75%±0.59%; p<0.01) but not Vδ1 T cells (figure 1B,C). In addition, the percentages of peripheral Vδ2 T cells of RA were negatively correlated with the levels of inflammatory markers, including C reactive protein, erythrocyte sedimentation rate as well as the Disease Activity Score in 28 joints (r=−0.6341, n=42, p<0.01; figure 1D). However, no correlation was observed between peripheral Vδ2 T cells and the titres of rheumatoid factor or anticyclic citrullinated peptide antibodies (figure 1D). Taken together, these results suggest peripheral Vδ2 T cells were closely related to RA, which suggested a role in the pathogenesis of RA.

Peripheral Vδ2 T cells were lower in patients with RA. Peripheral blood mononuclear cells obtained from patients with RA, patients with OA and HCs were stained with anti-CD3, anti-γδ TCR, anti-Vδ1 or anti-Vδ2 mAb followed by flow cytometry. The solid plots represent isotype controls, and the open plots represent indicated staining. The left panels show flow cytometry data of (A) γδ T cells, (B) Vδ1 T cells or (C) Vδ2 T cells. The right panels show bar graphs of the percentage of positively stained cells. Representative data of RA (n=30), HC (n=15) and OA (n=15) are shown. (D) The percentage of peripheral Vδ2 T cells in RA is negatively correlated with CRP, ESR and DAS28 (n=42). Results are expressed as mean±SEM. ns, no significance; **p<0.01 by one-way analysis of variance with Tukey-Kramer post-hoc test. Correlations are calculated using Spearman correlation analysis. Anti-CCP, anti-cyclic citrullinated peptide; CRP, C reactive protein; DAS 28, Disease Activity Score in 28 joints; ESR, erythrocyte sedimentation rate; HC, healthy control; OA, osteoarthritis; RA, rheumatoid arthritis; RF, rheumatoid factor; TCR, T cell receptor.
Vδ2 T cells accumulated in RA synovium and were proinflammatory
We then set out to investigate the mechanisms that led to the lower population of peripheral Vδ2 T cells in RA. We found that the proliferation rate of Vδ2 T cells in RA was comparable with that in OA or HC (RA 90.03±7.81% vs HC 82.53±14.97% vs OA 84.77%±6.51%; p>0.05) (online supplementary figure S1A). Also, the apoptosis rates of Vδ2 T cells in RA, OA and HC did not show any significant difference (RA 0.68±0.22% vs HC 0.88±0.56% vs OA 0.96%±0.37%; p>0.05) (online supplementary figure S1B). Therefore, the peripheral reduction of Vδ2 T cells in RA did not result from abnormal proliferation or apoptosis capacity.
Given the previous observation of accumulated γδ T cells in RA SF,16 we then examined the infiltration of Vδ2 T cells in the joints of RA. Consistently, we found a significantly higher percentage of Vδ2 T cells in RA SF compared with OA SF (5.29%±0.76% vs 1.25±0.44%; p<0.05 (figure 2A). In addition, we found a significantly higher infiltration of Vδ2 T cells in RA than in OA synovium when examining the cells from enzyme-digested fresh synovium (1.48%±0.19% vs 0.41±0.08%; p<0.05 (figure 2B), as well as immunohistochemical staining of the synovium (36.00%±3.60% vs 2.33±0.33%; p<0.05) (figure 2C). These findings suggested that peripheral Vδ2 T cells in RA potentially migrated and accumulated in the synovium.
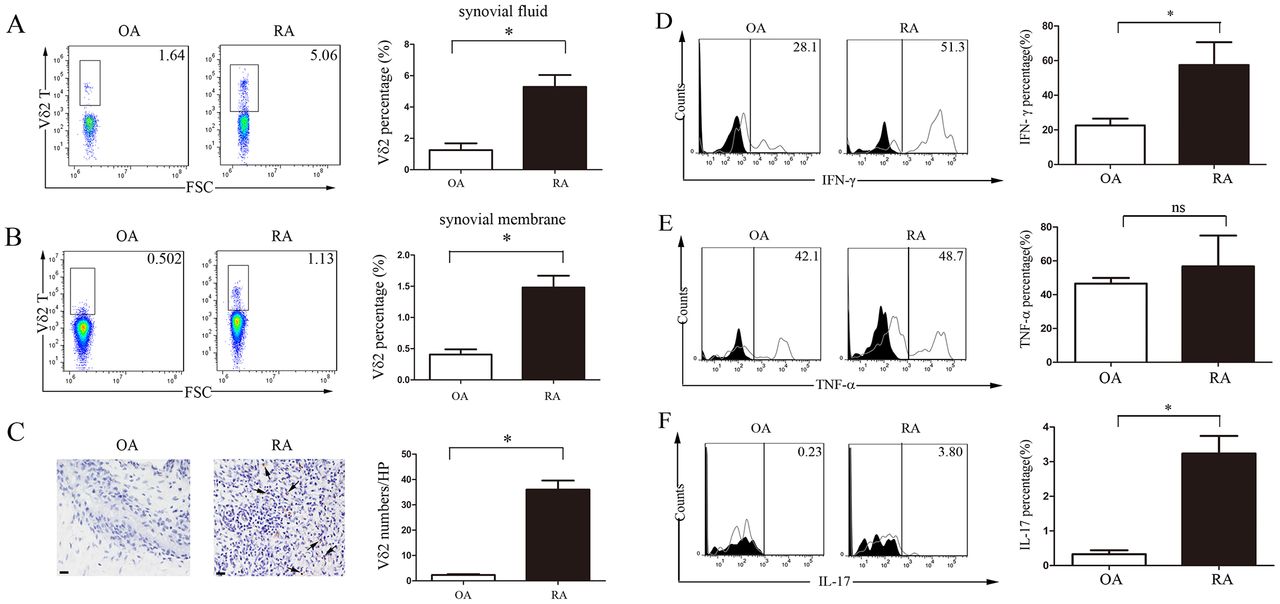
Vδ2 T cells accumulated at the affected joints of RA and secreted high levels of IFN-γ and IL-17. (A,B) The percentage of Vδ2 T cells in (A) SF and (B) enzyme-digested fresh synovium analysed by flow cytometry. Representative data of OA (n=4) and RA (n=4) are shown. (C) Infiltrations of Vδ2 T cells in the knee joint synovium of RA and OA were examined by immunohistochemical staining. Representative data of OA (n=3) and RA (n=3) are shown. Scale bars represent 50 µm. (D–F) Flow cytometry analyses of the intracellular staining of (D) IFN-γ, (E) TNF-α and (F) IL-17 in Vδ2 T cells from RA and OA synovium were performed. Data are representative of three independent experiments. The right panels show bar graphs of the percentage or the average number of positively stained cells. Results are expressed as mean±SEM. *p<0.05 by Student’s t-test. FSC, forward scattering; IFN-γ; interferon-γ; IL-17, interleukin-17; OA, osteoarthritis; RA, rheumatoid arthritis; SF, synovial effusion; TNF-α, tumour necrosis factor-α.
Similar to natural killer cells, Vδ2 T cells possess highly cytotoxic activity and produce proinflammatory cytokines, including IFN-γ and TNF-α (online supplementary figure S2D,E).3 4 We found that Vδ2 T cells from RA synovium produced higher levels of IFN-γ (57.43%±7.63% vs 22.60±2.26%; p<0.05) (figure 2D) and IL-17 (3.23%±0.30% vs 0.33±0.06%; p<0.05) (figure 2F) compared with Vδ2 T cells from OA synovium, although no significant difference in TNF-α production was observed in Vδ2 T cells between RA and OA (56.77%±10.51% vs 46.57±4.90%; p>0.05) (figure 2E). Similarly, peripheral Vδ2 T cells from RA synthesised more IFN-γ and IL-17 compared with those from OA and HC (online supplementary figure S2A–C). In addition, Vδ2 cells produced more TNF-α and IFN-γ than CD3+ T cells that were depleted of Vδ2+(CD3+Vδ2−) as well as total CD3+ T cells. Moreover, CD3+Vδ2− cells produced approximately 20%–35% less TNF-α and 25%–50% less IFN-γ than total CD3+ cells from both PBMC and synovial fluid of patients with RA (online supplementary figure S2D–E). These data suggest Vδ2 T cells from RA aberrantly secrete high levels of IFN-γ and IL-17, and potentially contribute to the pathogenesis of RA.
Chemotaxis of Vδ2 T cells to synovium was driven by CCR5 and CXCR3
To elucidate the mechanism of Vδ2 T cells accumulation in RA synovium, we performed in vitro transwell assay to examine the capacity of Vδ2 T cells chemotaxis in RA. As expected, we found that RA SF significantly promoted the recruitment of RA but not OA Vδ2 T cells (p<0.01, figure 3A). In contrast, neither RA nor OA SFs promoted the recruitment of RA Vδ1 T cells. These results suggest specific recruitment of Vδ2 T cells to the synovium in RA.

CCR5 and CXCR3 upregulation promoted Vδ2 T cell chemotaxis in RA. (A) Transwell migration assay: freshly isolated peripheral blood mononuclear cells from HC, OA and RA were loaded in the upper chamber, and SFs of OA, RA or medium were loaded in the lower chamber in the transwell invasion model. (B) The percentage and MFI of indicated chemokine receptors on Vδ1/Vδ2 T cells of RA. (C) Comparison of the proportion and MFI of indicated chemokine receptors in Vδ2 T cells from RA, HC and OA. (D) Vδ2 T cell migration assay with RA serum in the presence or absence of neutralising antibodies against CCR5, CXCR3 and CXCR6. (E) The concentration of known ligands of CCR5 and CXCR3 in SF of RA (n=22) and OA (n=10), and serum of RA (n=12) and OA (n=7). Data were pooled from three independent experiments (A,D) or five independent experiments (B,C). Results are expressed as mean±SEM. *p<0.05, **p<0.01 by one-way analysis of variance with Tukey-Kramer post-hoc test (A,C,D,E) and Student’s t-test (B). HC, healthy control; MFI, mean fluorescence intensity; OA, osteoarthritis; RA, rheumatoid arthritis; SF, synovial effusion.
The recruitment of leucocytes to target tissues is regulated by chemokines and corresponding receptors. We performed chemokine receptor expression profile screening, and found the expressions of CCR5, CXCR3 and CXCR6 on Vδ2T cells were significantly higher compared with those on Vδ1T cells (p<0.05, figure 3B). Moreover, the expressions of CCR5 and CXCR3 on RA Vδ2T cells were significantly higher than HC or OA Vδ2T cells (p<0.05, figure 3C). Furthermore, we applied neutralising antibody assay to verify the effect of these upregulated chemokine receptors on Vδ2 T cells recruitment, and we demonstrated that neutralising CCR5 and CXCR3 completely abrogated the migration capacity of RA Vδ2 T cells (figure 3D). Next, we examined the SF and serum levels of the known ligands of CCR5 (MIP-1α, MIP-1β, regulated on activation, normal T cell expressd and secreted (RANTES)) and CXCR3 (MIG, IP-10, I-TAC). We found all ligands except RANTES were significantly elevated in RA SF compared with OA SF and serum (p<0.01, figure 3E). Taken together, these data indicated that CCR5 and CXCR3 upregulation on Vδ2 T cells, in combination with high levels of their ligands in SF, work cooperatively to promote the recruitment of Vδ2 T cells to the affected joints in patients with RA.
TNF-α and IL-6 upregulated the expression of CCR5 and CXCR3 on Vδ2 T cells
To explore the triggering factors responsible for CCR5 and CXCR3 upregulation in RA, we cultured HC Vδ2 T cells with RA, OA and HC serum. Not surprisingly, we found the expressions of CCR5 and CXCR3 were significantly increased in the presence of RA serum (p<0.05, figure 4A), which were abrogated by administration of neutralising antibody against TNF-α, but not IL-17 (p<0.05, figure 4B). To support this finding, we demonstrated that TNF-α alone could significantly upregulate the expressions of CCR5 and CXCR3 on HC Vδ2 T cells (p<0.05, figure 4C). In addition, similar phenotypes were observed in IL-6-neutralised RA serum or IL-6-treated HC Vδ2 T cells (online supplementary figure S3). These results taken together suggest that TNF-α and IL-6 potentially play a role in the upregulation of CCR5 and CXCR3 expression on RA Vδ2 T cells.

TNF-α augmented the expression of CCR5 and CXCR3 on Vδ2 T cells. Flow cytometry analysis of (A) CCR5 and CXCR3 expression on Vδ2 T cells at indicated time points in the presence of HC, OA or RA serum; or (B) RA serum in combination with neutralising antibodies against TNF-α or IL-17 for 3 days; or (C) with medium in the presence or absence of TNF-α for indicated days. Data were pooled from three independent experiments. Results are expressed as mean±SEM. ns, no significance; *p<0.05, **p<0.01 by two-way ANOVA (A,C) or one-way ANOVA (B). ANOVA, analysis of variance; HC, healthy control; IL-17, interleukin-17; MFI, mean fluorescence intensity; OA, osteoarthritis; RA, rheumatoid arthritis; TNF-α, tumour necrosis factor-α.
NF-kB signalling pathway was involved in TNF-α-mediated upregulation of chemokine receptors in RA
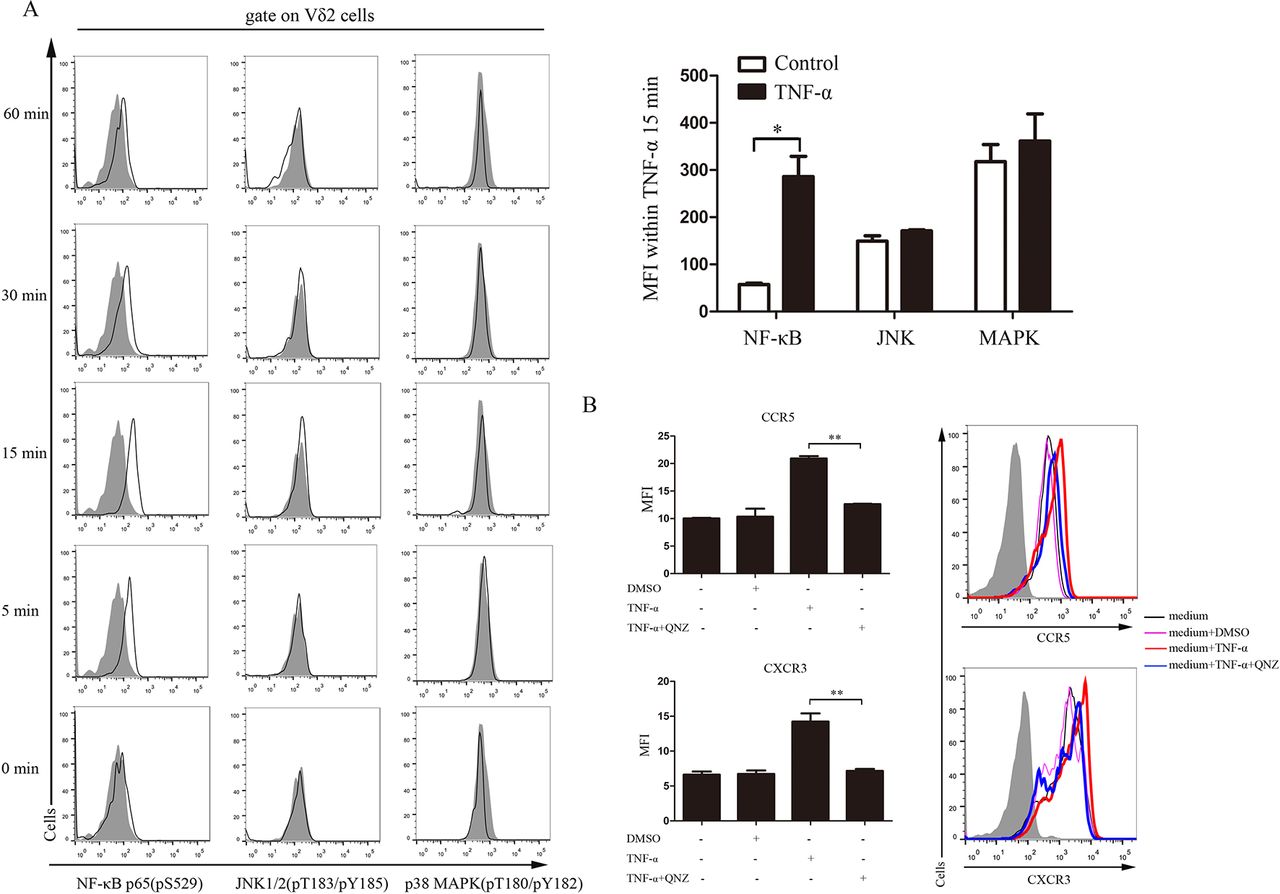
TNF-α is a multifunctional cytokine involved in apoptosis, cell survival, inflammation and a variety of immune responses via two distinct receptors17: TNFR-1 (p60) on all cells, and TNFR-2 (p80) that are mainly expressed on immune cells.18 The major signalling pathway of TNFR-associated factor 2 (TRAF-2) is activation of nuclear factor kappa B (NF-κB) via NF-κB-inducing kinase and inhibitor of nuclear factor kappa-B kinase (IKK) complex. In addition, TRAF-2 also phosphorylates mitogen-activated protein kinase (MAPK) and c-Jun N-terminal kinase (JNK) to activate c-Fos/c-Jun transcription factors.19 To find out which signalling pathway participates in the upregulation of chemokine receptors on Vδ2T cells, we detected phosphorylation of NF-κB p65 (pS529), JNK1/2 (pT183/pY185) and p38 MAPK (pT180/pY182) in TNF-α-treated Vδ2T cells by Phosflow at different time points. The results showed that NF-κB, but not JNK or MAPK, was phosphorylated on TNF-α stimulation (figure 5A). Furthermore, blocking NF-κB signalling with its specific inhibitor (QNZ) completely abrogated the upregulation of CCR5 and CXCR3 on Vδ2 T cells by TNF-α (figure 5B). Together, these findings indicate that NF-κB signalling was involved in the TNF-α regulated expression of CCR5 and CXCR3 on Vδ2 T cells.

NF-κB signalling pathway was involved in the expression of CCR5 and CXCR3 on Vδ2 T cells. (A) Vδ2 T cells were treated with TNF-α (100 ng/mL) for indicated time. The cells were permeabilised and stained with antibodies against NF-κB p65 (pS529), JNK1/2 (pT183/pY185) or p38 MAPK (pT180/pY182). The data represent one of three independent experiments. The right panels show bar graphs of MFI of Vδ2 T cells stimulated with TNF-α in 15 min. (B) Flow cytometric analysis of chemokine receptor expressions on TNF-α-stimulated HC Vδ2 T cells pretreated with QNZ (5 µM) or dimethyl sulfoxide (DMSO) for 1 hour. The solid plots represent isotype controls, and the open plots represent indicated staining. Results are expressed as mean±SEM. *p<0.05, **p<0.01 by paired t-test (A) and one-way analysis of variance with Tukey-Kramer post-hoc test (B). HC, healthy control; JNK, c-Jun N-terminal kinase; MAPK, mitogen-activated protein kinase; MFI, mean fluorescence intensity; NF-κB, nuclear factor kappa B; TNF-α, tumour necrosis factor-α.
TNF-α antagonist therapy restored Vδ2 T cells in patients with RA in vivo
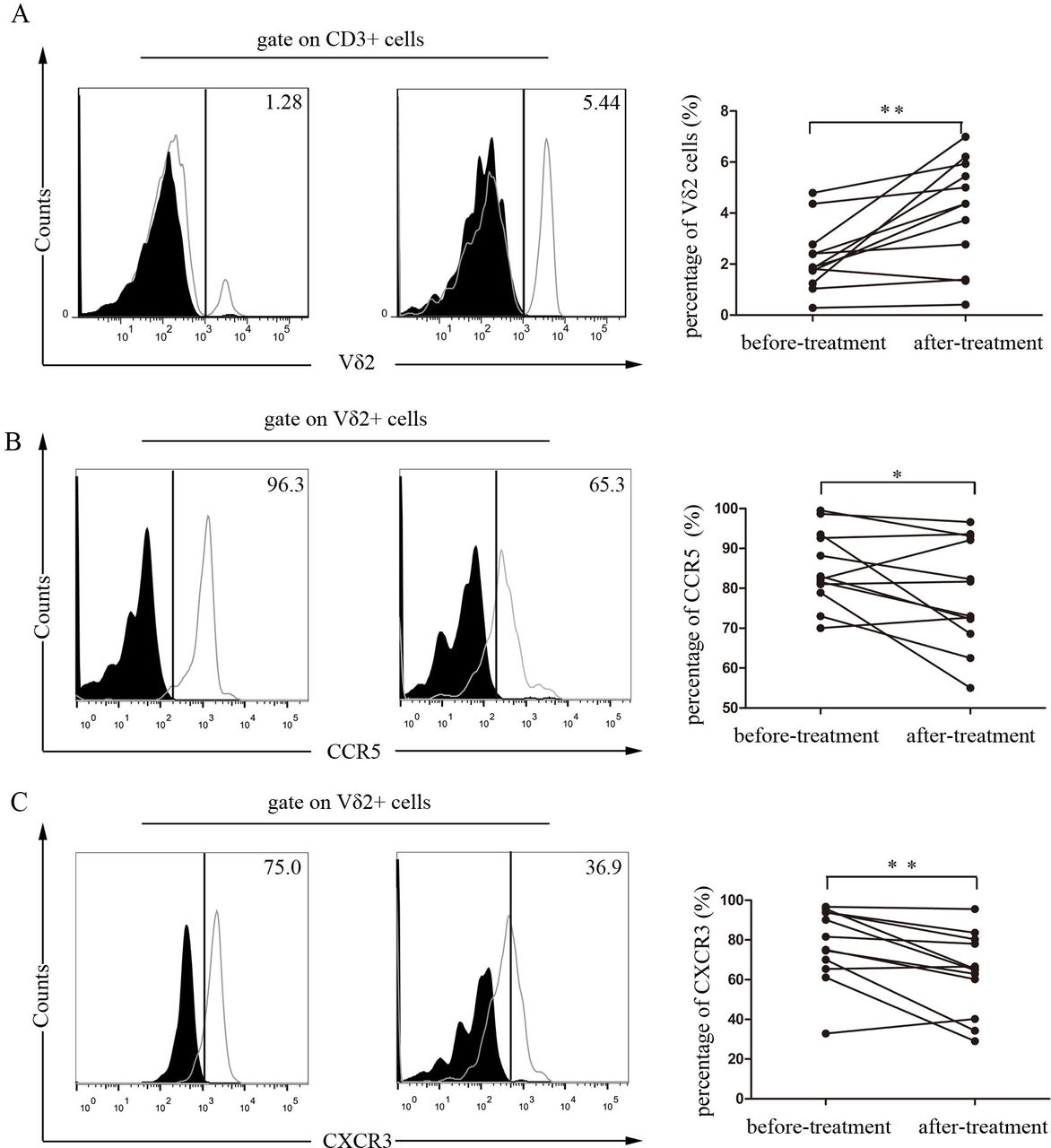
To investigate whether TNF-α regulated the chemotaxis of Vδ2 T cells in vivo, we examined the peripheral Vδ2 T cells from patients with RA treated with etanercept, a kind of TNF-α receptor fusion protein. We found that the percentage of Vδ2 T cells could be restored in patients with RA after treatment with etanercept (figure 6A). Furthermore, TNF-α antagonist treatment downregulated the expression of CCR5 and CXCR3 on Vδ2 T cells in patients with RA (figure 6B,C). We also detected the effect of IL-6 receptor antagonist on modulating peripheral Vδ2 T cell population and the expression of CCR5 and CXCR3. However, no significant effect was observed (online supplementary figure S4). Taken together, these findings further support the conclusion that TNF-α specifically regulates peripheral Vδ2 T cell trafficking to RA synovium by modulating the expressions of CCR5 and CXCR3 in vivo.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
TNF-α antagonist therapy repopulated peripheral Vδ2 T cells and downregulated CCR5 and CXCR3 expressions in patients with RA. Treatment-naïve patients with RA (n=12) were treated with etanercept in combination with methotrexate for 3 months. Flow cytometry was performed for the analysis of (A) the percentage of peripheral Vδ2 T cells and the expressions of (B) CCR5 and (C) CXCR3 on Vδ2 T cells before and after treatment. The solid plots represent isotype control, and the open plots represent Vδ2 T cells staining. *p<0.05, **p<0.01 by paired t-test. RA, rheumatoid arthritis; TNF-α, tumour necrosis factor-α.
Discussion
All the data in this study collectively suggested the hypothesis that in patients with RA, peripheral Vδ2 T cells potentially infiltrated into the synovium and secreted high levels of proinflammatory cytokines, which contributed to the pathogenesis of RA. Mechanistically, we further showed that elevated level of serum TNF-α in patients with RA induced high expressions of CCR5 and CXCR3 on Vδ2T cells, which promoted Vδ2 T chemotaxis, and NF-kB signalling pathway was involved in this process. More strikingly, anti-TNF-α therapy restored the peripheral Vδ2 T cells population as well as the expression of CCR5 and CXCR3 in patients with RA.
Previous reports show a higher proportion of γδT cells in RA SFs and synovium compared with peripheral blood.20 21 In our study, a significant lower percentage of peripheral Vδ2 T cells in patients with RA was noted, which might be caused by intensive accumulation into synovial tissues but not abnormal cell apoptosis or proliferation potential. Additionally, both Vδ2 cells from peripheral blood and from SF of patients with RA showed proinflammatory phenotype, which produced higher levels of IFN-γ and IL-17 compared with controls. Consistently, peripheral blood and SF from patients with RA contained heterogeneous γδT cells dominated with effector memory Vγ9/Vδ2 T cells producing inflammatory cytokines including IFN-γ and IL-17.22 Animal studies also show that γδT cells are the major source of IL-17 in joints and their increasing numbers are correlated with disease activity.16 23 24 In collagen-induced arthritis model, preventive depletion of γδ T cells significantly delayed the onset and severity of arthritis.12 24 Moreover, depletion of Vγ4+ cells, the counterpart of Vδ2 T cells in human and the major population of γδ T cells in mice, significantly attenuates arthritis severity, incidence of arthritis and anticollagen antibodies production. Taken together, our data suggest Vδ2 T cells are involved in the pathogenesis of RA, and targeting Vδ2 T cells might be a promising approach to treat RA.
We further explored the underlying mechanism of abnormal accumulation of Vδ2 T cells in synovial tissues. Chemokine receptors are expressed in various types of cells and interact with chemokines to promote ‘homing’ of cells to target tissues. By profiling chemokine receptors, we found CXCR3 and CCR5, the chemokine receptors preferentially expressed on IFN-γ-producing Th1 cells,25–28 were highly expressed on Vδ2 T cells and were essential for their migration to synovial tissues. Additionally, we confirmed that the corresponding agonistic ligands except RANTES were significantly elevated in SF of RA29 30 and promoted migration of circulating γδT cells.31 These chemokines are produced locally by synovial fibroblasts and TNF-α stimulation promotes their production.32 Collectively, these findings suggest upregulated CXCR3 and CCR5 in Vδ2 T cells potentially orchestrated with elevated chemokines to promote Vδ2 T migration.
Monocytes, macrophages and synovial fibroblasts produce high levels of cytokines including TNF-α and IL-1 on stimulation in RA.33 TNF-α, one of the major proinflammatory cytokines in RA, is a potent stimulator of synovial fibroblasts, osteoclasts and chondrocytes to release matrix metalloproteinases, which ultimately lead to joint destruction and bone degradation. In contrast, blocking TNF-α with its antagonist significantly reduces the production of matrix metalloproteinases. Additionally, neutralising TNF-α reduced production of other proinflammatory cytokines including IL-1, IL-6, IL-8 and gannulocyte-macrophage colony stimulating factor (GM-CSF).34–36 In our study, neutralising TNF-α in vivo dramatically restored the Vδ2 T population in patients with active RA. Mechanically, TNF-α regulates CCR5 and CXCR3 expression on Vδ2 T cells via NF-kB signalling, which is an important pathway of many inflammatory process. Additionally, p65(RelA), a member of NF-kB/Rel family, is a potent activator of the CCR5 promoter.32 37 Collectively, we suggest a novel alternative mechanism of action of TNF-α antagonist: anti-TNF-α therapy downregulates CCR5 and CXCR3 expression of Vδ2 T cells, and subsequently reduces Vδ2 T accumulation in synovial tissues and ameliorate arthritis.
Intriguingly, despite anti-IL-6 in RA serum abolished upregulation of CCR5 and CXCR3 in Vδ2 T in vitro and IL-6R antagonist therapy ameliorated RA disease in vivo, IL-6R antagonist did not rescue the Vδ2 T population in vivo. Elevated level of IL-6 after tocilizumab therapy38 39 might account for the attenuated repopulation of Vδ2 T cells.
The impact of Vδ2T cells on bone metabolism remains elusive. We showed Vδ2T cells of RA produced high levels of IFN-γ and IL-17, especially in synovial tissues. IL-17 and receptor activator for nuclear factor-kappa-B ligand (RANKL) are the major cytokines promoting osteoclast differentiation and activation, which ultimately lead to bone erosion. IL-17 stimulates osteoblasts to produce RANKL, which in turn induces differentiation of osteoclast progenitors into mature osteoclasts.40 Additionally, IL-17-secreting γδ T cells prime αβ T cells to produce IL-17 and enhance the function of Th17 cells.41 In contrast, the role of IFN-γ in osteoclast genesis is controversial. Although animal studies report that IFN-γ inhibits osteoclast formation,42 clinical studies have failed to demonstrate the efficacy of IFN-γ administration in bone loss prevention.43–46 The implication of elevated IFN-γ of Vδ2 T cells is yet to be further elucidated.
Moreover, Vδ2 T cells potentially contribute to RA pathogenesis in other ways. The majority of adult Vγ9/Vδ2 T cells express the CD45RO memory phenotype2 with memory CD45RA-CD27+ subset and effector CD45RA-CD27− subset.47 The effector memory Vγ9/Vδ2 T cells exhibit phenotypic characteristics of specific antigen-presenting cells, including high human leukocyte antigen DR (HLA-DR) and CD80/86 expression, which promotes B cell activation and polarises adaptive immunity towards a Th1 immune response in patients with RA.22 Both circulating Vδ2 cells and residential Vδ2T cells produce a variety of cytokines and chemokines including MIP, RANTES and IL-8,48 49 inducing macrophage aggregation and activated T lymphocytes migration,50 which may also be involved in the pathogenesis of RA.
In summary, we demonstrate that Vδ2T cells were lower in peripheral blood and accumulated in the RA joint and secreted increasing amounts of proinflammatory cytokines that are involved in the pathogenesis of RA, which resulted from its upregulation of CCR5 and CXCR3 induced by TNF-α via NF-kB signalling pathway. Elucidation of the roles of Vδ2 T cells in RA advances our knowledge in understanding the complex pathogenetic mechanism of RA, and provides an alternative mechanism of biological agents to develop new promising biomarker of therapy, and exploring potential Vδ2 cell-targeted therapy.
Acknowledgments
We thank the health professional staff from the Department of Rheumatology and Clinical Immunology, Peking Union Medical College Hospital and the patients for their participation in this study. We preferentially thank Chen Zhou, Sai-Nan Bian and other healthy controls for donation of blood. We thank Lin-Fang Huang and Lin-Feng Lian for their advice and technical assistance. We thank Dr Austin Cape for careful reading and feedback.
References
Footnotes
W-XM, S-SY and HC contributed equally.
Handling editor Tore K Kvien
Contributors W-XM, S-SY and HC contributed equally to this manuscript. XZ and WH conceptualised and designed the project and supervised the project. W-XM, S-SY and HC performed all the experiments and wrote the manuscript with contributions from all authors. J-MZ revised the manuscript. CZ, L-FH, L-DZ, Y-YF, H-XY and WZ participated in the sample collection and clinical analysis. All authors read and approved the manuscript.
Funding This study was supported by grants from the National Natural Science Foundation of China (81325019, 81630044, 81273312, 81601432, 81550023), the National Science Fund for Distinguished Young Scholars of China (813250046), Beijing Municipal Natural Science Foundation (7141008), National Key Research and Development Program: ‘Precise Medical Research’ (2016YFC0903900), CAMS Innovation Fund for Medical Sciences (2016-12M-1-003, 2016-12M-1-008,2017-12M-1-007), and ‘Stem Cell and Translational Research’ (2016YFA0101001).
Competing interests None declared.
Patient consent Obtained.
Ethics approval Institutional Review Board of Peking Union Medical College Hospital.
Provenance and peer review Not commissioned; externally peer reviewed.