Article Text

Abstract
Objective: Thymic function declines exponentially with age. Impaired thymic function has been associated with autoimmune disease in adults but has never been formally assessed in childhood autoimmunity. Therefore, thymic function in children with the autoimmune disease juvenile idiopathic arthritis (JIA) was determined.
Methods: Thymic function was measured in 70 children and young adults with JIA (age range 2.1–30.8 (median 10.4)) and 110 healthy age-matched controls using four independent assays. T cell receptor excision circles (WBLogTREC/ml) and the proportion of CD4+ CD45RA+CD31+ T cells (representing recent thymic emigrants; %RTEs) were quantified and intrathymic proliferation measured by calculating the αTREC/ΣβTREC ratio. Lastly, regulatory T cells (TReg) of thymic origin (CD4+FOXP3+) were quantified in peripheral blood to assess the ability of the thymus in JIA to generate this T cell subset.
Results: Thymic function was equivalent by all four parameters in JIA when compared with the control population. Furthermore, there was no consistent effect of JIA subtype on thymic function, although intrathymic proliferation was higher in the small rheumatoid factor (RF)+ polyarticular group. There were no significant effects of disease-modifying antirheumatic drugs (DMARDs) or oral corticosteroids on thymic function, although those with the worst prognostic ILAR (International League of Associations for Rheumatology) subtypes were also those most likely to be on a DMARD.
Conclusions: It is demonstrated that children and young adults with JIA, unlike adults with autoimmune diseases, have thymic function that is comparable with that of healthy controls. The varied pathologies represented by the term “JIA” suggest this observation may not be disease specific and raises interesting questions about the aetiology of thymic impairment in adult autoimmunity.
Statistics from Altmetric.com
The thymus plays a critical role in the development of normal immune tolerance.1 As well as negatively selecting potentially autoreactive T cells, it also positively selects the naturally occurring regulatory T cell subset (TReg) (CD4+FOXP3+).2 Cross-sectional studies have demonstrated impaired thymic function in several adult autoimmune diseases.3–6 It remains unclear whether the thymic defect is primary or secondary, although single gene defects affecting thymic integrity can predispose to autoimmunity.7 8 In contrast to adults, thymic function has not been formally quantified in childhood autoimmunity such as juvenile idiopathic arthritis (JIA).
The normal thymus is largest in childhood and is well known to decline in size and function with increasing age.9 Whilst not normally of apparent consequence, this decline becomes important under circumstances where the T cell pool is depleted, such as in HIV-AIDS or during lympho-ablative treatments such as bone marrow transplant or stem cell transplantation (SCT). This (normal) decline in thymic function with ageing is thought to explain the slower and often less complete T cell reconstitution following SCT in adults when compared with that in children.10 11
JIA describes a heterogeneous group of diseases12 ranging in severity from severe polyarticular disease sometimes associated with systemic features, to more benign oligoarticular disease. The more severe JIA subtypes have a poor prognosis, with lack of response to intense immunosuppression in some cases.13 14 Autologous SCT has recently been shown to induce prolonged, drug-free remission in a proportion of such refractory patients.15
Measuring thymic function in humans is complicated by the lack of a specific phenotype for recent thymic emigrants (RTEs), although the surface profile CD4+CD45RA+CD31+ has been proposed as a potential marker.16 Rearrangement of the T cell receptor α chain (TCRα) gene results in the formation of an episome of DNA called a “T cell receptor excision circle” (TREC).17 TRECs are present only in thymically derived T cells and do not divide with cellular mitosis. When quantified per millilitre of whole blood, TRECs are widely accepted to be the optimal measurement of thymic function.18
In the current study we compare thymic function in children with JIA and healthy controls. By measuring TRECs, the proportion of CD4+CD45RA+CD31+ T cells and TReg in peripheral blood, and intrathymic T cell development using a novel assay, we demonstrate that thymic function is not compromised in JIA.
PATIENTS AND METHODS
Study subjects
Following ethical approval and informed consent from the child and/or a parent or guardian, 9 ml blood samples were obtained from healthy control children undergoing simple surgical procedures. Samples from young adults were donated by healthy volunteers. Study subjects were consecutive attendees at the Newcastle Regional Paediatric Rheumatology Service in Newcastle. All were given a diagnosis of JIA classified according to the ILAR (International League of Associations for Rheumatology) classification by a consultant Paediatric Rheumatologist.12 Samples from older patients were obtained at adolescent and young adult rheumatology clinics. Blood was collected into Vacuette EDTA K3 tubes (Greiner Bio-one, Kremsmünster, Austria).
DNA extraction
DNA was extracted from 300 μl of whole blood using the Wizard Genomic DNA extraction kit (Promega, Madison, Wisconsin, USA) and stored at 4°C. Sample purity and quantity was determined by spectrophotometry (Nanodrop ND-100, Wilmington, Delaware, USA).
TREC quantification
We have developed a method quantifying TRECs/ml in DNA extracted from 300 μl of whole blood19. Briefly, WBLogTREC/ml is determined from a simultaneously amplified standard curve (range 107–101 TREC molecules) using quantitative real-time PCR (RQ-PCR), an ABI Prism 7900HT Sequence Detector System and SDS2.2 software (Applied Biosystems, Warrington, UK).
Reactions (25 μl) contained primers CACATCCCTTTCAACCATGCT and GCCAGCTGCAGGGTTTAGG both at 700 nM, 150 nM Taqman hydrolysis probe (6-FAM-ACACCTCTGGTTTTTGTAAAGGTGCCCACT-TAMRA), 12.5 μl of JumpStart Taq ReadyMix (Sigma, Poole, UK) and 200 ng of DNA. Thermal cycling conditions were 50°C for 2 min then 95°C for 10 min, then 40 cycles of 95°C for 15 s and 60°C for 1 min. Experimental samples were run in duplicate and averaged.
Quantification of intrathymic proliferation
Intrathymic proliferation was measured using a modified version (to allow for use with SYBR Green) of the method published by Dion et al.20 Ten TCRβ chain Dβ→Jβ rearrangements (βTRECs) and the TCRα chain δrec→ψJα rearrangement (αTRECs) were quantified in separate, nested PCRs. A duplex first round reaction with primers for individual βTRECs and genomic CD3 was followed by individual RQ-PCRs. The CD3 copy number multiplied by 0.5 (since each cell contains two copies) allowed the βTREC frequency to be reported per 105 cells.
First-round 25 μl PCRs contained 1× High Fidelity PCR buffer (Invitrogen, Paisley, UK), 0.2 mM each of dNTPs (Invitrogen), 1.5 mM MgCl2 (Invitrogen), 1 U of Platinum Taq High Fidelity (Invitrogen), 500 nM of both primers (Sigma Genosys, UK) and 150 ng of target DNA. Initial denaturation was at 95°C for 10 min followed by 22 cycles of 95°C for 30 s and 72°C for 30 s. Second-round, 25 μl reactions contained 2× SybrGreen PCR Master Mix (TAKARA, Berkshire, UK), 400 nM of each primer (Sigma Genosys) and 10 μl of template. Thermal cycling conditions were 95°C for 10 s followed by 40 cycles of 95°C for 5 s and 60°C for 60 s.
Surface staining of peripheral blood mononuclear cells (PBMCs) for quantification of recent thymic emigrants
A 70 μl aliquot of whole blood was incubated in the dark at room temperature for 15 min, with combinations of antibodies to surface antigens: CD31–PE (phycoerythrin), CD45RA–FITC (fluorescein isothiocyanate; Serotec, Oxford, UK) and CD4–PE–Cy5 (Beckton Dickinson, Oxford, UK). Stained samples were incubated with 1× FACS Lyse Solution (Beckton Dickinson) for 10 min and washed twice.
Isolation of PBMCs from whole blood and intracellular cell staining
PBMCs were isolated according to a standard, sucrose density gradient (Lymphoprep) protocol and cryopreserved in 10% dimethylsulfoxide (DMSO) in fetal calf serum (FCS) at −80°C until use. Thawed cells were surface stained with CD4+–FITC and then FOXP3–APC (allophycocyanin) antibody (eBiosciences, Wembley, UK) according to the manufacturer’s instructions. Flow cytometry was performed using a FACScan and data were analysed using FloJOv6.1.2. Sample identities were blinded to the assessor.
Statistical analysis
Statistical analyses were performed using SPSS 11th edition software. Values for WBLogTREC/ml were initially log transformed to allow parameteric testing. Population distributions were tested for normality with a one-sample Kolmogorov–Smirnov test. Correlations for normally distributed data were tested for significance using Pearson’s correlation coefficient. Differences between groups were assessed by analysis of covariance (ANCOVA) where other covariates were present, unless otherwise stated. Bonferroni corrections were applied to correct for multiple comparisons. Results were considered significant when p⩽0.05.
RESULTS
Thymic function declines with age throughout childhood and does not differ between patients with JIA and healthy controls
WBLogTREC/ml was quantified in 70 patients with JIA and 110 healthy controls (HCs). Table 1 describes the cohort. One additional patient had DiGeorge syndrome (chromosome 22q11 deletion-associated syndrome with thymic hypoplasia/aplasia) and a co-existing inflammatory arthritis. Figure 1A shows that there was no significant difference in thymic function between the groups after adjusting for age and gender (see below). Negative correlations with age were detected for both groups (p<0.01). The patient with DiGeorge syndrome had markedly deficient thymic function as evidenced by a 1.5Log deficit in WBLogTREC/ml when compared with age-matched controls. There was no difference in thymic function when ILAR JIA subtypes were compared (fig 2A).

Thymic function is not significantly different in children with juvenile idiopathic arthritis (JIA) (grey diamonds, dashed regression line) and healthy controls (HCs) (filled squares, solid regression line) when quantified using WBLogTREC/ml (T cell receptor excision circles) (A) and by the proportion of T cells with the phenotype CD4+CD31+CD45RA+ (%RTE (recent thymic emigrants)) (B). WBLogTREC/ml was significantly negatively correlated with age (r = −0.762; p<0.001 for controls and r = 0.625; p<0.001 in JIA) as was %RTE (r = −0.656; p<0.001 and r = 0.491; p<0.001 respectively). In a single patient with co-existent DiGeorge syndrome (grey triangle with dashed circle surround), TREC levels were markedly reduced, but %RTE was age appropriate. In both HCs (C) and the JIA group (D), females (grey diamonds, dashed regression lines) have greater thymic function than males (filled squares, solid regression line) using WBLogTREC/ml (p = 0.002 and p = 0.003 respectively).

Thymic function does not significantly differ between International League of Associations for Rheumatology juvenile idiopathic arthritis (JIA) subtypes when measured using either WBLogTREC/ml (T cell receptor excision circles) (A) or %RTE (recent thymic emigrants) (p = NS) (B). Bars represent age-adjusted group means (SEM). Groups were compared using analysis of covariance with age as a covariate and multiple comparisons corrected for by performing a Bonferroni correction. PolyRF+ and PolyRF–, polyarticular rheumatoid factor positive and negative, respectively; PsA, psoriatic; Enth, enthesitis associated; Ext oligo, extended oligoarticular; Persis oligo, persistent oligoarticular).
The proportion of CD4+CD45RA+CD31+ declines with age in HCs and patients with JIA
CD4+CD45RA+CD31+ T cells are TREC rich and have been proposed as the phenotype of RTEs in humans.16 The proportion of T cells with this phenotype (%RTEs) declined with age in patients with JIA (n = 50) and HCs (n = 77) (fig 1B), but there was no significant difference in age-adjusted mean %RTEs.
Since both WBLogTREC/ml and %CD4+CD45RA+CD31+ T
cells have been proposed as markers of thymic function, there should be a strong positive correlation between them, independently of disease status. Although the relationship was strong in HCs (r = 0.71; p<0.01), it was less robust in patients with JIA (0.45; p<0.01). There was no effect of ILAR subtype on %RTEs (fig 2B).
In our patient with DiGeorge syndrome, the %RTEs was normal despite a significantly reduced WBLogTREC/ml value (fig 1B), suggesting that CD4+CD45RA+CD31+ T cells may be capable of self-renewal in the absence of thymic replenishment.
Gender differences in thymic function are preserved in children with JIA
In both mice and adult humans, females have a larger thymic output (as determined by TREC quantification and histological analysis of thymic volume) than age-matched males.21 22 Using the two assays described above, we confirmed higher age-adjusted thymic output in females, in both HCs and patients with JIA (fig 1C,D). The age-adjusted mean WBLogTREC/ml value for HC females (n = 51) was 5.039 and for males (n = 59) 4.985 (p = 0.002); in the JIA group, the comparable figures were 5.004 (n = 46) and 4.826 (n = 24), (p = 0.003).
Using %RTEs, a gender difference was again detected in HCs (age-adjusted mean %RTEs in females (n = 36) 52.9%, males (n = 41) 45.6% (p = 0.001) but not in the JIA group (data not shown).
Effect of immunosuppressant therapy on thymic function in JIA
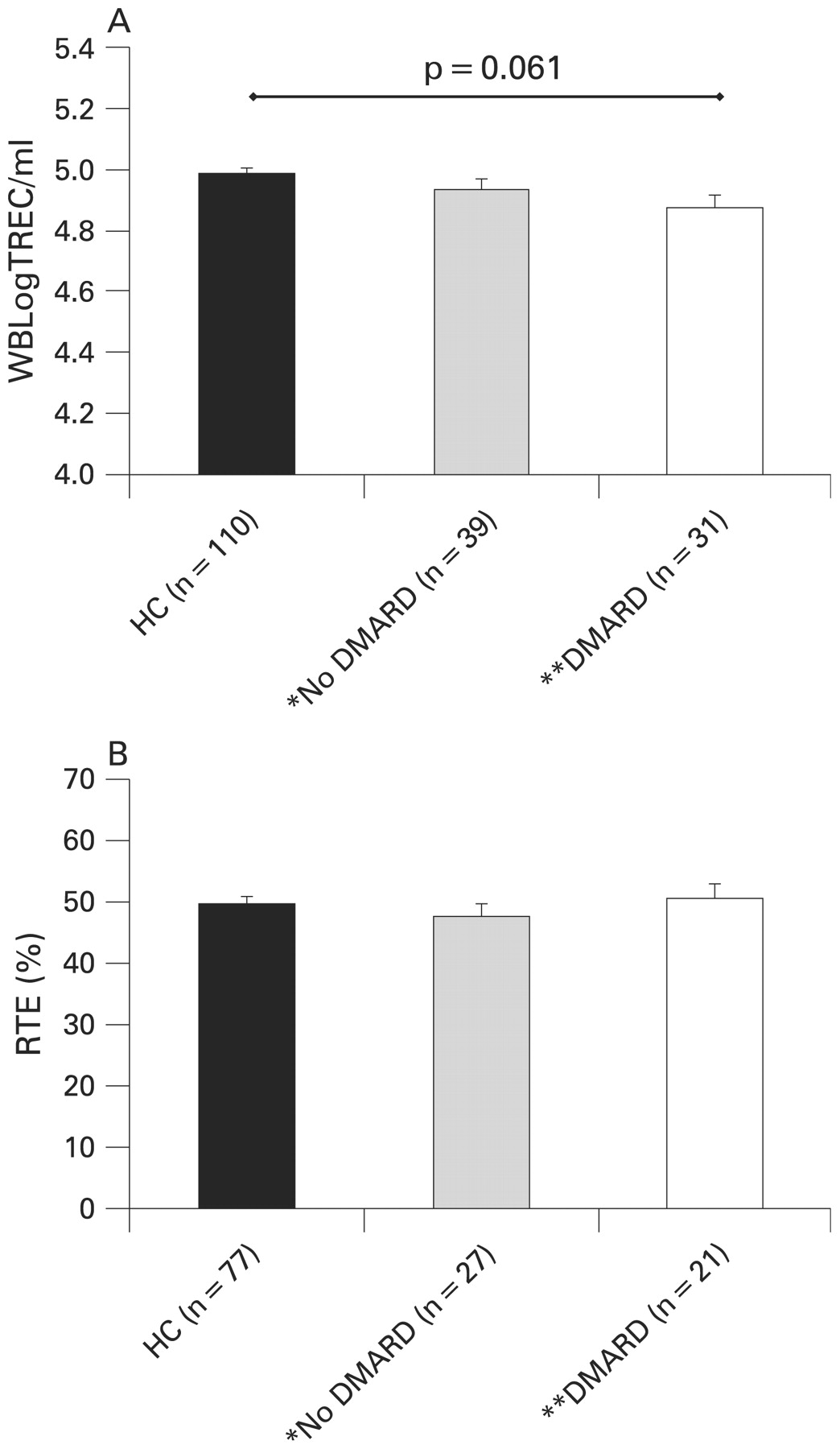
Corticosteroids and some immunosuppressive treatments have been considered to inhibit thymic function,23–25 and we therefore sought an effect in JIA. It should be noted that the majority of children taking DMARDs or oral corticosteroids had more severe disease, falling into the systemic-onset, polyarticular and extended oligoarticular subtypes (see table 1). However, when patients receiving these drugs were compared with those not receiving them and with healthy controls using WBLogTREC/ml, no evidence of a significant effect of DMARDs (when considered collectively (n = 31)) (4.874 vs 4.933 vs 4.985) (fig 3A) or corticosteroids (± DMARD) (n = 11) (4.818 vs 4.924 vs 4.985) was found (data not shown). There was, however, a trend toward lower WBLogTREC/ml values in patients with JIA on a DMARD (p = 0.061) and those taking steroids (± DMARD) (p = 0.07) when compared with HCs. There was no effect of taking methotrexate (MTX) on WBLogTREC/ml values (4.887) when compared with those not on MTX (4.917) and with HCs (4.985) (p = NS) although some patients not on MTX were taking an alternative DMARD (primarily antitumour necrosis factor α (TNFα)) (n = 8). Other treatment groups contained too few patients to exclude an independent effect. In contrast, after adjustment for age and gender, %RTEs was significantly higher in patients on oral corticosteroids (n = 8; 58.3%) than in those not on oral corticosteroids (n = 40; 47.0%; p = 0.013), and the comparison with HCs (n = 77) approached significance (49.8%; p = 0.067). There was no difference between those not on oral corticosteroids and HCs. There was no significant effect of DMARD use (fig 3B) or MTX specifically on %RTEs.

There was no significant effect on thymic function when measured using WBLogTREC/ml (T cell receptor excision circles) (A) or %RTEs (recent thymic emigrants) (B) of being on a disease-modifying antirheumatic drug (DMARD). Bars represent age- and gender-adjusted group means (SEM). Groups were compared by analysis of covariance with a Bonferroni correction to adjust for multiple comparisons. Patients not on a DMARD were more likely to have persistent oligoarticular disease than those taking one; see table 1. There was a trend toward lower WBLogTREC/ml values in those taking a DMARD (p = 0.061) when compared with healthy controls (HCs). *Includes those taking only non-steroidal anti-inflammatory drugs or relying only on local joint injections for disease control. **DMARDs include antitumour necrosis factor α agents and interleukin 1 receptor antagonist (IL1ra).
Intrathymic proliferation in JIA
When the TCRα chain gene rearranges, the locus encoding the TCRδ chain is excised in a conserved rearrangement, resulting in the formation of a TREC that can be identified in approximately 70% of emerging thymocytes. This rearrangement is the basis of most standard TREC assays, including our own WBLogTREC/ml assay.17 When the TCRβ chain gene is rearranged, no single rearrangement occurs, but there are only 13 potential Dβ→Jβ gene rearrangements.26 Dion et al devised an assay that quantifies 10 of these 13 TREC-generating rearrangements, the sum of which provides an estimate of the frequency of T cells in peripheral blood that have rearranged their TCRβ chain gene (ΣβTREC).20 TCRα chain rearrangement occurs after TCRβ chain rearrangement. Between these events a phase of thymocyte proliferation expands thymocytes with a rearranged TCRβ chain, providing a substrate for the subsequent generation of TCR diversity through α chain/β chain pairing. The ratio between the numbers of T cells containing detectable TCRβ and TCRα chain rearrangements (αTREC:ΣβTREC) therefore gives an indication of the number of divisions, which is a measure of intrathymic proliferation and overall thymic function.
We determined intrathymic function in patients with JIA and HCs. No difference was found when 19 children with JIA were compared with 19 HCs, and there was also no detectable effect of gender (data not shown). There were insufficient samples to seek an effect of corticosteroids or DMARDs. The effect of ILAR subclassification was assessed. There were no significant differences between the groups after post hoc testing, with the exception of all comparisons with the polyarticular RF+ group (n = 3) which (after the two groups where n = 1 were excluded) achieved statistical significance (fig 4A). The true significance of this result is difficult to assess because of small patient numbers, and was lost when we combined together the poor prognosis subtypes of systemic onset, polyarticular and extended oligoarticular JIA (“non-oligoarticular”, n = 9) and compared with persistent oligoarticular JIA (“oligoarticular” n = 11), a classification previously used27 (fig 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) The ratio between the numbers of T cells containing detectable T cell receptor β (TCRβ) and TCRα chain rearrangements (αTREC:ΣβTREC) (y-axis) was compared between International League of Associations for Rheumatology subtypes using analysis of covariance with Bonferroni correction for multiple comparisons, with age and gender as covariates because of their effect on the αTREC (T cell receptor excision circle) value. When the two subsets where n = 1 were excluded, the comparisons between the polyarticular rheumatoid factor (RF)+ group and the reminder were statistically significant (p<0.05)—although caution should be applied to the interpretation of this result given the small group numbers. Sys, systemic (n = 3); PRF+ and PRF−, polyarticular RF+/polyarticular RF− (n = 3 both groups); PsA, psoriatic (n = 1); ExO, extended oligoarticular (n = 1); PerO, persistent oligoarticular (n = 8). (B) When grouped into persistent oligoarticular disease and non-oligoarticular disease (see text), there were no significant differences between the groups. Boxes represent the interquartile ranges (IQR) and bars the group medians. Whiskers give the data range, with “•” representing extreme values (>3 IQR from the box end). HC, healthy controls.
In the patient with DiGeorge syndrome, the αTREC:ΣβTREC ratio was 57 in comparison with the HC group median (352) and the JIA group median (628), reflecting the impaired structural environment of the DiGeorge thymus and its reduced capacity for thymocyte expansion. The ΣβTREC value in this patient was comparable with JIA values 1.58 vs 5.38 (range 0.53–17.41), whereas the αTREC value was significantly reduced, 91 vs 3002 (range 851–5065). This implies that thymocyte proliferation is impaired in DiGeorge syndrome in comparison with HCs and children with JIA.
Thymus-derived TReg are more frequent in persistent oligoarticular JIA
Lastly, we quantified the proportion of CD4+ T cells with the regulatory phenotype CD4+FOXP3+ (TReg). There was no detectable difference overall between patients with JIA and HCs. When individual ILAR subtypes were compared, there was a weak but significant difference between the patients with systemic and persistent oligoarticular disease (p = 0.031) in a post hoc analysis, although sample sizes were again small (table 2). To strengthen our analysis by increasing group numbers, patients with “non-oligoarticular” disease (as above) were compared with patients with persistent oligoarticular disease. We found a significant increase in the proportion of TReg in the oligoarticular group (8.19% vs 6.481%; p = 0.044, Tukey HSD post hoc analysis), consistent with other reports. Unlike those reports, however, there was no difference from HCs. There was no effect of gender, age or DMARD use, although MTX use was more common in non-oligoarticular JIA (see table 1), potentially confounding the difference between disease subtypes.
DISCUSSION
Our data demonstrate that, in children with JIA, thymic function is comparable with that of healthy control children. This conclusion is strengthened by our analysis of multiple aspects of thymic function, including intrathymic proliferation and “downstream” effects on RTEs and TReg. Consistent with previous reports, we have demonstrated greater thymic function in females28 and an age-associated decline in healthy controls,29 findings that persist in JIA. In a single patient with inflammatory arthritis and co-existent DiGeorge syndrome, we showed a dramatic reduction in thymic function, consistent with thymic hypo/aplasia and similar to the effect of partial thymectomy during paediatric cardiac surgery.30 Although we have previously confirmed that consecutive recruitment of patients from our clinic population provides a group of patients with a typical case mix31 (table 1), the short recruitment period (4 months) may have biased our population toward those with more severe disease (since they are more likely to attend hospital).
When data were analysed according to ILAR disease subtype, a minority of comparisons reached statistical significance or demonstrated a trend in that direction. For example, there was a trend toward reduced thymic function for those on a DMARD (although since disease group and DMARD use co-segregate, this analysis is confounded by ILAR subtype), and steroid use was associated with an increase in the %RTEs. The αTREC:ΣβTREC ratio was increased in the ILAR polyarticular RF+ group, whilst the persistent oligoarticular group had a significantly higher proportion of TReg than the ILAR systemic disease group, consistent with previous reports. These isolated observations should be interpreted with caution due to the small group numbers, and they require further analysis in a larger study in order to determine their true significance.
T cells with the phenotype CD4+CD45RA+CD31+ have significantly higher TREC content per cell than unselected CD4+ T cells,16 confirming that this subset is enriched for true thymic emigrants. Furthermore, CD4+CD45RA+CD31+ T cell telomeres are longer and telomerase activity higher than in CD4+CD45RO+CD31+ T cells, suggesting a limited replicative history.32 This phenotype has been used to monitor thymic function longitudinally33–35 and, although interindividual variation is high, values are stable longitudinally35 (ARL and JDI, unpublished results). Thus, it was interesting to note that, despite an ∼1.5Log deficit in WBLogTREC/ml in a patient with DiGeorge syndrome, the proportion of CD4+CD45RA+CD31+ T cells was age appropriate. Furthermore, WBLogTREC/ml and %RTEs did not always correlate well, particularly in disease (lending support to the recent observation that CD4+CD45RA+CD31+ T cells can in part be maintained by peripheral homeostasis36), and thus may represent a less useful measure of thymic function in this setting. Naïve T cells proliferate excessively in patients with rheumatoid arthritis (RA), possibly under cytokine stimulation.37 A similar phenomenon in JIA would weaken the correlation between CD4+CD45RA+CD31+ T cells and WBLogTREC/ml, as we have shown here.
Regardless of its indication, adults fare less well than children following autologous SCT (ASCT).38 In part this reflects an inverse relationship between age and CD4+ T cell reconstitution39 40 but, in autoimmunity, disease-related thymic compromise may exacerbate the distinction. For example, patients with RA remain lymphodepleted for years following ASCT,41 but disease usually relapses rapidly despite persisting lymphopenia. In contrast, patients with JIA reconstitute more rapidly and, moreover, thymically derived TReg may contribute to long-term remission of symptoms.42 Thus, measures aimed at reversing age-associated thymic atrophy (currently in clinical trial43 44) could significantly improve the outcome of ASCT in adults with autoimmune diseases.
The normal thymus produces T cells that are self-tolerant, with a polyclonal TCR repertoire. Age-related thymic involution results in a proportionally greater decline in thymocytes with rearranged TCRα genes than with TCRβ chain genes,20 with potential implications for repertoire diversity,45 and therefore the maintenance of self-tolerance, responses to neoantigens and tumour surveillance.46 We determined a ratio that reflected the number of cellular divisions between these two events. The similarity between patients (when collectively considered) and controls suggests that this step in thymocyte development is intact in JIA. When individually considered, however, there was a significant increase in the ratio for the polyarticular RF+ patients. A single extreme value contributes to this effect, but the remaining two individual values were also greater than for all other subtypes (fig 4A), a consequence of both low βTREC and high αTREC values (data not shown). The significant contribution of the αTREC value to the ratio and the variance of this value in both patients with JIA and HCs (fig 1) should also be noted, and duplication of our results is required before further conclusions about this difference can be drawn. If confirmed, however, this result would suggest enhanced intrathymic proliferation in the JIA subtype that most closely resembles adult RA, in which there is evidence of impaired thymic function.3 TCR diversity and TRECs are positively correlated,47 suggesting that the TCR repertoire in JIA is also likely to be maintained.
De Kleer et al distinguished two subsets of TReg in JIA peripheral blood following ASCT.42 Expansion of mature, non-depleted or grafted TReg in the lymphopenic environment was followed 6–8 months later by the appearance of TReg of recent thymic origin (which may have greater suppressive function than memory CD45RO+ TReg48), together replenishing the depleted pretransplant TReg pool.42 Although we did not distinguish TReg subsets, we found a higher proportion of TReg in persistent oligoarticular disease than in non-oligoarticular disease although we could not confirm a difference between either group and HCs, possibly reflecting our use of FOXP3 to quantify TReg, rather than CD25bright.42 We have previously shown that adults with self-limiting reactive arthritis have a higher proportion of TReg than adults with RA,49 and murine studies have demonstrated that Foxp3+ T cells protect against autoimmune disease and inflammation in vivo.50 Collectively, these data suggest that control of inflammatory disease may be associated with this subset of T cells. Indeed, all four patients with systemic JIA studied here had a very low proportion of TReg in peripheral blood. However, their otherwise normal thymic function suggests that their TReg deficit may reflect a failure of peripheral TReg homeostasis in progressive, chronic disease.
In conclusion, using a variety of measurements, we have demonstrated age-appropriate thymic function in patients with JIA. We found no clear influence of therapy on thymic function, although DMARD use was more common in patients with JIA with a poor prognosis and was associated with trends toward lower WBLogTREC/ml values. We also found similar TReg values in HCs and patients with JIA, although patients with JIA of the better prognosis subtype had higher values. Our findings differ from those in adult patients with autoimmune disease, in whom thymic dysfunction has been implicated as a possible aetiological factor.3–6
Acknowledgments
We gratefully acknowledge D Douek and RP Sekaly for providing TREC standard constructs, and Mick Eltringham for considerable help with collection of study samples. This work was funded by an Arthritis Research Campaign Clinical Research Fellowship to ARL (Grant Reference L0547) and an Arthritis Research Campaign Integrated Clinical Arthritis Centre Grant (Grant Reference ICAC 16361).
REFERENCES
Footnotes
Competing interests: None.
Funding: This work was funded by a Clinical Research Fellowship (ARL) and a Studentship (TAM) from the Arthritis Research Campaign (UK)
Ethics approval: Ethical approval was obtained
Patient consent: Informed consent obtained was from each child and/or a parent or guardian.
This is an open-access article distributed under the terms of the Creative Commons Attribution Non-commercial License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.