Article Text

Statistics from Altmetric.com
Over the past 40 years rheumatoid arthritis (RA) has been the subject of extensive immunological and histological investigations. Rheumatoid synovial tissues, obtained from synovectomy, joint arthroplasty or biopsy have been examined by various pathologists and researchers using a range of ever more sophisticated techniques. From conventional histology using haematoxylin and eosin, to immunolocalisation, in situ hybridisation, and more recently to the use of extracted RNA for specific gene analysis. From early studies describing synovial A and B cells as the major components of rheumatoid synovium, we now have a better understanding of T cell subsets, monocyte-macrophage populations, and the expression of disease related genes such as cytokines and proteinases. This knowledge undoubtedly provides a better understanding of the inflammatory processes that operate in rheumatoid synovial tissue; but the cardinal feature of RA is cartilage and bone degradation. Far fewer studies have been reported on these sites of cartilage erosion or bone loss, yet it is here that the catabolic processes responsible for joint destruction are manifest.
Our interest in the ‘rheumatoid lesion’, a term used to describe the cartilage-pannus junction or sites of cartilage erosion, started in 1976 with the finding of the enzyme collagenase at sites of cartilage loss.1 ,2 Since then we have examined numerous cartilage-pannus specimens derived from more than 150 different cases of knee joint surgery, using a variety of immunological and histochemical techniques to identify specific cell types and different matrix degrading enzymes. During these studies we have become aware of the broad range of histological findings and the variations in proteinase expression.3 ,4 As knowledge of the mechanisms responsible for joint destruction is essential for the development of preventative pharmacological treatments, this overview presents our observations on the complex cellular and enzymic processes involved in cartilage degradation and subchondral bone remodelling of the rheumatoid joint.
The rheumatoid lesion
Although for surgical reasons the rheumatoid tissues examined were derived exclusively from middle or late stage disease, many of the total knee arthroplasty specimens provided tissue samples that showed both advanced and ‘new’ cartilage lesions, as judged by the relative thickness of the cartilage and the extent of pannus development or synovial cell overlay. Cartilage-pannus junctions were obtained from 150 different RA knee joints, and of these approximately 30 provided samples of subchondral cartilage or bone erosion sites. In many cases four or five sites of cartilage erosion were sampled from the same specimen with each site usually showing quite different histological features.3 ,5 ,6 The residual cartilage of each specimen has also allowed an assessment of intrinsic or extrinsic chondrolysis.7 Our histological findings for these three anatomical sites are described separately.
CARTILAGE-PANNUS JUNCTIONS
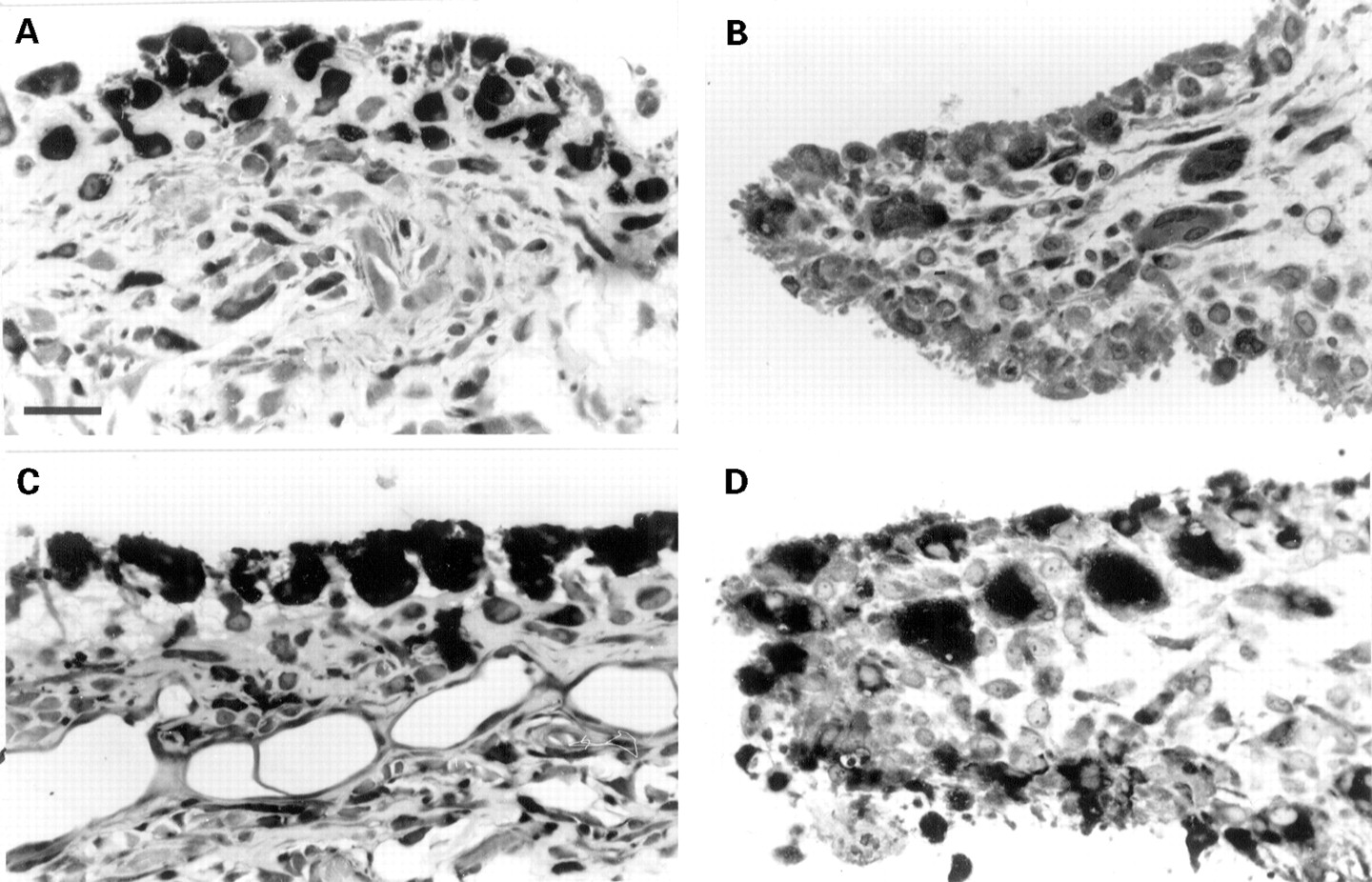
More than half of the samples examined demonstrated ‘cellular’ junctions with pockets of invasive cells penetrating the cartilage matrix (figs 1 and 2). It was evident that the cells that occupied the cartilage interface usually showed a different cellular density and morphology compared with those of the supporting pannus tissue (figs 1and 2). The cell types that occupy cartilage-pannus junctions include macrophages, fibroblasts, mast cells,8 polymorphonuclear leucocytes (PMNs),9 and displaced, probably dedifferentiated chondrocytes.10 Macrophages and fibroblasts were commonly observed as majority cell types in most samples (figs 1 and 2E, F), but in a significant proportion of specimens these were often interspersed by local accumulations of mast cells or PMNs.6 ,11 Early lesions with a relatively thin mononuclear cell overlay often showed PMNs as the dominant cell type (figs 2A, B), but these cells were rarely seen at the erosion sites of specimens showing well developed pannus tissue with vascularisation and fibrous matrix deposition. Such findings are in accord with previous studies reporting either the presence9 12 or the relative absence6 13 14 of PMNs in the rheumatoid lesion.

Bidirectional erosion of articular cartilage and variations in cellular composition at sites of cartilage erosion. (A) Photomicrograph showing synovial pannus overlay of cartilage and bone. Note increased synthesis of proteoglycan by chondrocytes. Methylene blue, bar = 70 μm. (B) Bidirectional attack on articular cartilage with multinucleated osteoclasts/chondroclasts (arrows) on the subchondral side and PMNs and monocytes of the synovial tissue. Methylene blue, bar = 70 μm. (C) Bidirectional erosion of cartilage with a mixed population of cells and blood vessels at the subchondral interface, and a clearly defined synovial cell layer at the cartilage interface. Toluidine blue, bar = 70 μm. (D) Cartilage erosion site stained for α napthyl acetate esterase (αNA). Note strongly stained cells, especially in close association with cartilage matrix. Bar = 70 μm. (E) Focal accumulation of αNA positive cells at cartilage-pannus junction. Bar = 70 μm. (F) Cellular heterogeneity at cartilage erosion site. Note the variable morphologies of synovial cells and the lack of chondrolytic activity by the chondrocytes. Toluidine blue, bar = 50 μm.

Examples of microenvironmental proteinase expression at the rheumatoid lesion. (A) and (B) PMNs stained for chloroacetate esterase at the cartilage-pannus junction. Observations of PMNs were usually confined to thin overlays of synovial tissue, usually representing early erosive lesions. Bars = 30 μm. (C) and (D) Mast cells stained for tryptase with evidence of extracellular enzyme often associated with stromal oedema and matrix disruption (arrows). Bars = 70 μm and 30 μm respectively. (E) and (F) Extensive expression of acid phosphatase by cells at the cartilage-pannus junction (E), and by a proportion of larger macrophagic cells (F). Bars = 30 μm.
Our observations indicate that the cellular composition along most cartilage-pannus junctions show many variations and show that even adjacent cartilage erosion sites are often occupied by different proportions or permutations of these four cell types. Moreover, tissue samples from different sites of the same joint often showed relatively acellular, thickened pannus indicative of a rather quiescent, non-inflammatory lesion. Such disparate findings in discrete microfoci along the cartilage-pannus junctions of the same specimen are best explained as microenvironments that resemble different phases of inflammatory or reparative cycles. Thus histological differences exist not only between specimens, but also within samples from the same RA joint.3 No evidence was found for a common or regular pattern for the cellular erosion of cartilage. Of special significance, considering the importance attached to the initiating role for T lymphocytes in rheumatoid disease,15 ,16 was the realisation that these cells were seldom seen at the cartilage interface. It has been suggested that T cells are not important for chronic synovitis and cartilage erosion,17 but the presence of T cells remote from cartilage erosion sites was observed for many specimens. As T cell lymphokines are known to stimulate a variety of cell types, especially in relation to the production of autoantibodies, cytokines, and prostanoids, it seems probable that T lymphocytes continue to influence the pathophysiological processes associated with the advanced rheumatoid lesion.
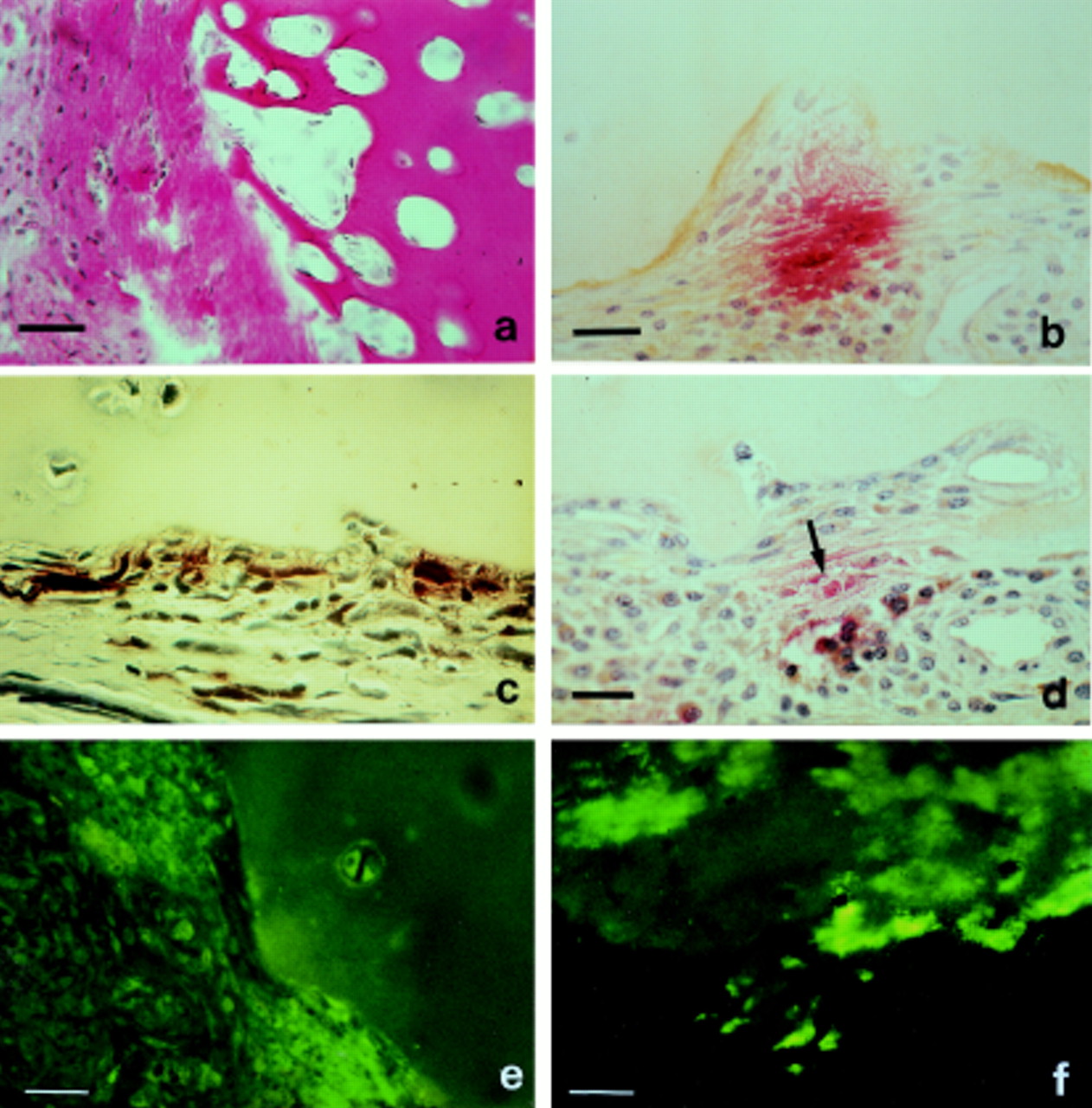
Histological findings of bone-pannus junctions usually showed the number of inflammatory cells to be much reduced (fig 3A, B). However, focal accumulations of cells, often acid phosphatase positive compared with the surrounding synoviocytes, were often observed along the bone-pannus lesion (fig 3A, B). These indications of increased lysosomal content appeared typical of cells located close to the mineralised tissue, possibly suggesting a change in the resorptive phenotype favouring the production of acid hydrolases, a common feature of osteoclasts and chondroclasts.5 ,18

Heterogeneity of specific cell types at bone:pannus junctions and subchondral erosion sites. (A) Bone:pannus junction showing focal variations in cellularity and the expression of acid phosphatase activity. Bar = 70 μm. (B) High power magnification of (A) showing acid phosphatase positive cells in juxtaposition with mineralised bone. Bar = 30 μm. (C) Subchondral bone fragment showing alkaline phosphatase staining of osteoblastic cells interspersed with an osteoclast (arrow). Bar = 30 μm. (D) Alkaline phosphatase activity of osteoblasts in subchondral bone at site of extensive remodelling. Bar = 30 μm. (E) Osteoclasts stained for acid phosphatase at site of subchondral bone resorption. Bar = 30 μm. (F) Multinucleated chondroclasts stained for acid phosphatase at subchondral site. Note resorptive attachment to hyaline cartilage. Bar = 30 μm.
SUBCHONDRAL BONE AND CARTILAGE EROSION
Osteoporosis and juxta-articular bone loss are common features of late stage rheumatoid disease.5 ,19 ,20 Approximately 40% of all samples that included bone showed extensive subchondral remodelling of the mineralised tissue. The expression of alkaline phosphatase by osteoblasts was especially prominent in some areas suggestive of new bone formation, but not in others. Even within tissue sites where many osteoblastic cells were stained for alkaline phosphatase these cells were often interspersed with multinucleated osteoclasts (fig 3C). Such observations indicate the complexities of bone remodelling in subchondral sites of rheumatoid knee joints, where osteolytic activity of bone resorbing cells is often countered by attempts at new bone formation (fig 3C, D).
The subchondral sites of bone and cartilage degradation showed different cellular compositions to that of the synovial pannus tissue. Increased numbers of multinucleated cells were often seen at resorption sites for bone, as well as calcified and hyaline cartilage (fig3E, F). Chondroclasts are morphologically and histochemically similar to osteoclasts, but are defined by their intimate association with the cartilaginous matrix, in contradistinction to osteoclasts, which are exclusively associated with mineralised bone.5 These multinucleated cells are important features of subchondral lesions (fig3F); once the calcified cartilage is breached the hyaline cartilage often appears to be infiltrated by bone derived cells, especially acid phosphatase positive macrophages and small blood vessels (fig 4C, D). Such findings seem analogous to those reported for the normal development of the tibial epiphyseal growth plate where hyaline cartilage resorption was associated with capillary ingrowth whereas mineralised cartilage was removed by chondroclasts.21 ,22

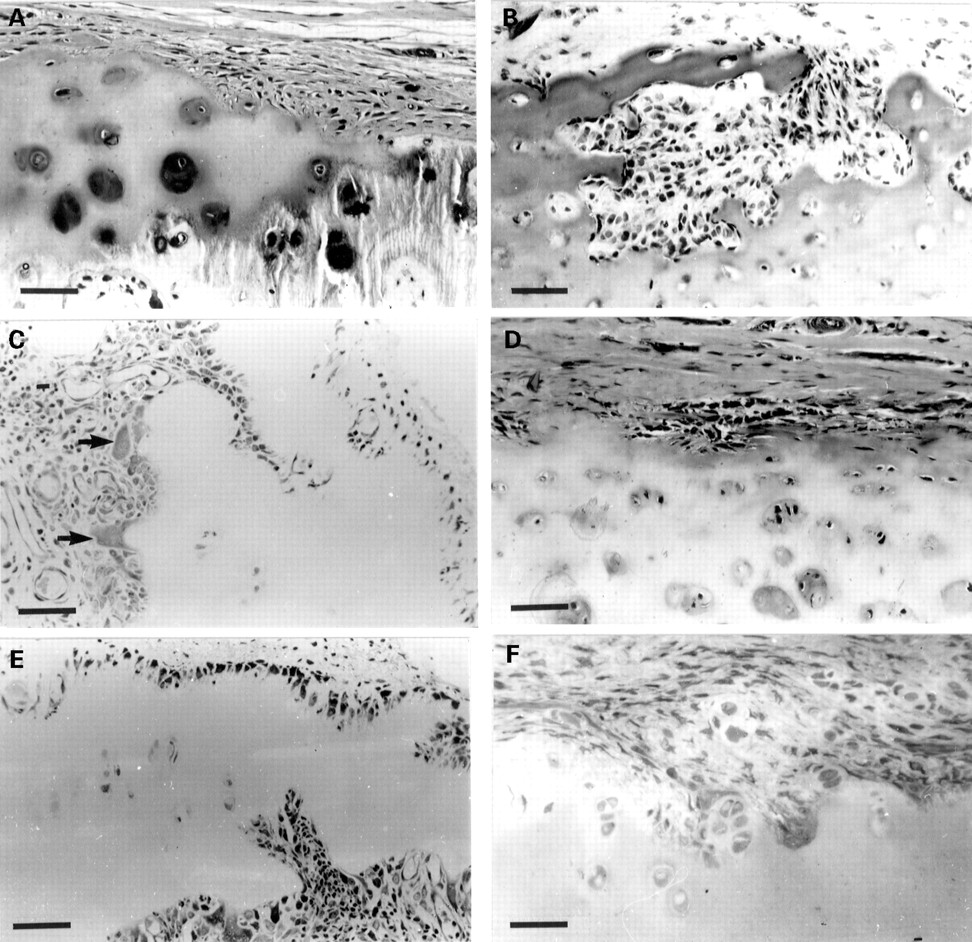
Examples of some unusual histological observations of the rheumatoid lesion. (A) Cartilage erosion site in which the chondrocytes demonstrate a ‘normal’ morphology at the junction and the retention of this by the apparently displaced chondrocytes (arrow). Methylene blue, bar = 30 μm. (B) Cartilage-pannus junction characterised by focal accumulations of mast cells (arrows) at the cartilage interface. Note the pronounced chondrolytic activity of subjacent chondrocytes (compared with (A)). Methylene blue, bar = 40 μm. (C) and (D) Unusual accumulations of acid phosphatase positive cells within hyaline cartilage. Evidence of blood vessels and blood cells (arrows) suggests a subchondral infiltration with mono and binucleated cells strongly positive for acid phosphatase—possibly immature multinucleated cells. Bars = 70 μm and 30 μm, respectively. (E) Cartilage-pannus junction with multinucleated giant cells stained for acid phosphatase in contact with hyaline cartilage. Bar = 30 μm. (F) Cartilage-pannus junction showing a group of small blood vessels at the forefront of the invasive pannus tissue. Methylene blue, bar = 30 μm.
The bidirectional attack on articular cartilage observed here for late stage rheumatoid knee tissues indicates that the two erosive fronts, synovial and subchondral, are characterised by different cell types.18 This is seldom seen in small joint histopathology, which often illustrates a continuous pannus tissue brought about by the ‘capping’ or ‘underpinning’ process.3 ,23 ,24 Our studies have shown that multinucleated cells with acid phosphatase activity are seldom seen at cartilage-synovial pannus junctions, in contrast with their regular presence in subchondral erosion sites. One rare example of multinucleated cells at the cartilage-pannus junction is shown in figure 4E; another unusual or infrequent histological finding was the presence of small blood vessels at the forefront of the invasive synovial pannus tissue (fig 4F), this being much more common for subchondral erosion fronts.
CHONDROCYTE MEDIATED CHONDROLYSIS
The contribution of chondrocytes to early pannus development has been suggested25 26 and in a minority of specimens we have observed hyperplastic and hypertrophic outgrowths of chondrocytes from the cartilage, in accord with other reports.27 ,28 However this is relatively uncommon and may reflect secondary osteoarthritic processes rather than a rheumatoid lesion with the presence of inflammatory cells. No more than 20-25% of the cartilage-pannus junctional specimens examined showed signs of chondrolytic activity by chondrocytes, and few have shown evidence of cartilage necrosis. While we agree that extrinsic cartilage degradation brought about either by the synovial pannus or bone derived erosion fronts is the usual observation 7 there is good evidence for intrinsic chondrolytic activity in a minority of specimens, not least the finding of collagen degradation products around chondrocytes in even the deeper zones of the cartilage matrix.22 Chondrocyte mediated cartilage degradation was often observed where the invasive synovial pannus contained local concentrations of mast cells (fig 4B), or exceptionally dense distributions of inflammatory cells.29Serial sectioning confirmed that the enlarged lacunae were not occupied by invasive synovial cells and suggested that chondrocytes within the vicinity of proinflammatory mediator production may respond in much the same way as activated synoviocytes.16 In vitro, articular chondrocytes behave in several respects similar to synovial fibroblasts; both have receptors for cytokines such as interleukin 1 (IL1) and tumour necrosis factor (TNF)α and β, and respond to these ligands with increased production of prostaglandin E2(PGE2) and metalloproteinases.16 ,30 Mast cell histamine is another potent mediator with stimulatory effects on chondrocytes, as well as synoviocytes and endothelial cells. It is of special interest that human chondrocytes express both histamine H1 and H2 receptors, their stimulation resulting in increased PGE2 and cyclic-AMP production.31 ,32
In general, most specimens showed chondrocytes with a histologically normal appearance close to the erosive front, and occasionally some displaced chondrocytes in the pannus tissue appeared to retain both their morphology (fig 4A) and their synthetic phenotype.10Moreover, some tissue samples from the same joint specimen demonstrated either localised chondrolytic activity, as judged by enlarged lacunae, or localised proteoglycan synthesis determined by differential toluidine blue staining.3 Such observations indicate that, as with the extensive subchondral bone remodelling processes, chondrocytes may assume both catabolic and anabolic activities in neighbouring locations16 22 presumably in response to the cells and signals of the rheumatoid lesion.
MULTINUCLEATED SYNOVIAL GIANT CELLS
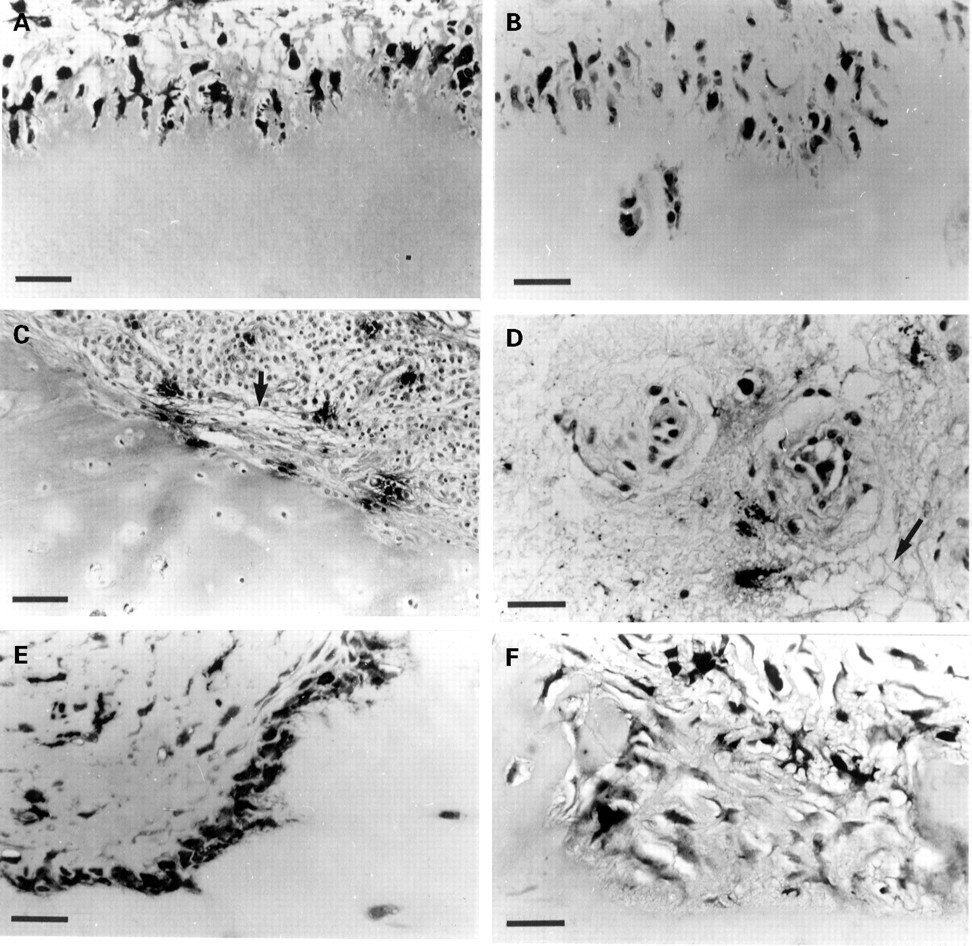
Synovial giant cells were observed only in a minority of the rheumatoid synovial specimens examined, usually as components of either the synovial lining layer or intimal layers (fig 5). These multinucleated cells were shown to contain non-specific esterase (fig5A, B) and acid phosphatase (fig 5C and D), properties purportedly characteristic for macrophage derived (CD68 positive) giant cells,33 as well as multinucleated osteoclasts and chondroclasts.5 Unlike the latter, which were consistently associated with resorptive subchondral sites, the synovial giant cells seldom occupied the synovial-cartilage interface, and did not appear to be associated with any disruption or breakdown of their surrounding extracellular matrix.
Esterase and acid phosphatase production by giant cells of rheumatoid synovium. α Napthyl esterase staining of giant cells in the synovial lining (A) and intimal (B) layers. Acid phosphatase staining of giant cells in the synovial lining (C) and intimal (D) layers. Bars = 30 μm all micrographs.
Microenvironmental nature of cartilage degradation
The photomicrographs of figure 6 are selected to illustrate some of the microenvironmental features of the rheumatoid lesion. Fig 6A demonstrates the localised loss of collagen from articular cartilage brought about by the invasive front of synovial cells; note that the supporting pannus tissue shows little evidence of collagen degradation. Such observations suggest either that different cell types occupy the erosion sites or that the synovial cells exhibit a different proteolytic phenotype in that location. We have histological evidence for both these possibilities.
Microenvironmental observations related to sites of cartilage erosion. (a) Cartilage collagen loss at the rheumatoid lesion. Note abundant collagenous matrix (red) of the overlying pannus tissue. Van Giesons, bar = 70 μm. (b) Extracellular mast cell tryptase (red) indicative of mast cell degranulation at cartilage erosion site, together with cartilage bound stromelysin 1 (brown). Dual immunostaining, 4 bar = 30 μm. (c) Intra and extracellular non-specific esterase (brown) produced by cells at the cartilage-pannus interface. Note the relative absence of enzyme from pannus cells remote from the junction. α Napthyl acetate esterase.6 Bar = 30 μm. (d) Focal production of interleukin 1β (brown) by a group of cells adjacent to area of extracellular mast cells tryptase (red, arrow). Dual immunostaining.4 Bar = 30 μm. (e) Stromelysin 1 distribution at cartilage-pannus junction. Note patchy distribution with relatively little enzyme in supporting pannus tissue. FITC immunostaining. 4 Bar = 30 μm. (f) Collagenase 1 distribution at cartilage-pannus junction restricted to the cartilage surface and showing diffusion within the matrix. Note relative absence of enzyme in overlying pannus tissue. FITC immunostaining. 1 Bar = 30 μm.
Figure 6B shows localised mast cell degranulation as judged by prominent extracellular staining for mast cell tryptase. Tryptase is an enzyme unique to mast cell granules and has proved to be an excellent marker, as well as an indicator for mast cell degranulation or activation. Mast cells at sites of cartilage erosion is a comparatively new observation, but their accumulation at the cartilage-pannus junction is not uncommon.4 ,11 As the release of granular tryptase is accompanied by a concomitant release of other potent mediators such as histamine, heparin, leukotrienes, cytokines, and growth factors, it seems probable that mast cell activation would bring about changes in the behaviour of all neighbouring cells. Indeed, histological evidence for mast cell activation and its association with localised oedema and disruption of the connective tissue architecture has recently been reported.4 Moreover, observations of mast cell degranulation in some specimens has been colocalised with the production of the proinflammatory cytokines IL1 and TNFα, as well as the matrix degrading enzymes collagenase 1 and stromelysin 1.4 Such findings were usually restricted to microfoci of cartilage erosion sites, and have suggested a key role for this cell in matrix degradation, inflammation, and tissue repair.4 ,34-36
α Napthyl acetate or non-specific esterase activity is a recognised marker of macrophages6 33 and is especially prominent in synovial lining cells, as well as activated synovial fibroblasts.37 Esterase staining was commonly observed for cells occupying cartilage erosion sites and occasionally for chondrocytes in close proximity to such sites (fig 6C). However it is emphasised that such observations were restricted to specific sites and rarely extended along the whole cartilage-pannus junction of any one specimen.
The proinflammatory cytokines IL1 and TNFα are reported to play important parts in cartilage and bone degradation,16 ,30and TNFα has been proposed as having a pivotal role in the pathogenesis of RA.38 Its expression by monocyte-macrophages has been demonstrated both in synovial tissue and at the cartilage-pannus junction.39 Similarly, IL1 and its receptor have been demonstrated at sites of cartilage erosion, both by pannus cells as well as some chondrocytes.40 The localised production of IL1 is shown in figure 6D; although mainly restricted to macrophages of the synovial lining layer IL1 is also expressed at cartilage erosion sites.40 It is of interest that although macrophages are recognised sources of both TNFα and IL1, in many specimens it is only a proportion of the macrophage numbers that express these cytokines at the time of surgery and tissue fixation. Thus it seems that these two potent cytokines, both capable of upregulating metalloproteinase and prostaglandin production by synoviocytes or chondrocytes,16 ,30 are also subject to regulation. The stimulatory agents for TNF and IL1 production include such diverse factors as endotoxin, immunoglobulins, heparin, and rheumatoid factors,16 but their localised expression in the rheumatoid lesion has also been correlated with mast cell activation,4 which is also a recognised source of TNFα.41
CARTILAGE DEGRADING ENZYMES
The proteinases involved in cartilage degradation are potentially numerous and inevitably relate to the specific cell types that occupy erosion sites. Matrix metalloproteinases such as collagenase 1, stromelysin 1 and others are primarily produced by synovial fibroblasts and chondrocytes.42 ,43 By contrast, the serine proteinases such as elastase, cathepsin G, chymase, and others are provided by granulocytes such as PMNs and mast cells.43 Most of the granulocytic serine proteinases are stored intracellularly, their release being stimulated by exposure to specific activating factors.44 However, metalloproteinase expression by mesenchymal cells is tightly regulated with several stimulatory factors, such as IL1 and TNFα, being responsible for the induction and release of these cartilage degrading enzymes.30 ,42 The important realisation is that enzyme release requires ‘activation’ of specific cell types, and although the regulation of proteinase expression is considered multifactorial it is very much a microenvironmental process. For example, figure 6E and F show the patchy distribution of stromelysin 1 and collagenase 1, respectively, at the cartilage:pannus junction. Although the same cell types are observed along other portions of these junctional specimens most were not producing these enzymes. Compared with collagenase 1 our studies have shown that stromelysin was more frequently demonstrated in pannus tissue, and especially at cartilage-pannus junctions.4 However, most specimens showed relatively few cells producing enzyme at the time of surgery and fixation of the tissue, indicating that production of these matrix metalloproteinases is a transient, intermittent process rather than a persistent, autonomous process.1 ,3 ,4
Although several proteinases are known to attack and degrade the proteoglycan components of cartilage,22 ,43 degradation of the fibrillar collagen is restricted to the collagenase enzymes. As the loss of the ‘skeletal’ collagenous component of cartilage is essentially irreversible,2 ,3 the recognition of three different collagenases with the ability to degrade fibrillar type II cartilage collagen is currently of special interest.45These three collagenases show distinct tissue specificity, with collagenase 1 produced by synovial cells and chondrocytes, collagenase 2 by neutrophils and collagenase 3 by chondrocytes. All three share many similar properties but have different abilities to degrade cartilage type II collagen; collagenase 3 being far the most effective.46 Neutrophil collagenase and collagenase 3 have both been identified as products of articular cartilage in osteoarthritis47 48; as yet their precise role in cartilage degradation associated with the rheumatoid lesion is uncertain. Although many studies have focused on the importance of one particular enzyme in cartilage erosion, especially the matrix metalloproteinases, it is probable that a combination of enzymes, not least the serine proteinases, function in concert for effective chondrolysis.3 ,4 ,43
Our interpretation of the many disparate histological observations is that individual cartilage erosion sites are spatially and temporally separated, with the presence of specific cell types and degradative proteinases being regulated by locally coordinated signals. These include the cytokines, growth factors, neuropeptides, prostaglandins, complement and local degradation products, which may all contribute to the local recruitment of cells and the regulation of their proteolytic expression. Such factors have comparatively short-term functions, their stimulatory effects probably contribute intermittently to the progressive long term destruction of articular cartilage. However, it is probably the autoimmune nature of RA that provides the primary stimulus for progressive cartilage degradation, an observation partly borne out by the finding that total removal of cartilage from a diseased joint suppresses or eliminates further inflammation.22
CARTILAGE AUTOIMMUNITY
Cartilage has long been recognised as an immunologically reactive tissue that can elicit both humoral and cellular immunity. The immune response is of paramount importance in the chronicity of RA, and evidence for the presence of circulating antibodies to various components of the cartilage matrix is now substantiated for a significant proportion of RA patients.22 ,49 Autoantibodies are often directed to the cartilage collagens type II and XI, and most frequently to their denatured or fragmented forms.50 ,51Antibodies specific for chondrocyte cell surfaces, proteoglycan core protein, aggrecan, and keratan sulphate are also well documented.22 Interestingly, these autoantibodies only appear after the clinical onset of rheumatoid disease, probably as a result of cartilage damage and the release of cartilage degradation products. This observation argues against a direct role for these antibodies in the initial pathogenesis of RA, but the formation of immune complexes and resultant cellular responses may well be causally related to the development of chronic inflammation and progressive cartilage destruction.22
The production of serum IgG, IgA, and IgM autoantibodies to cartilage collagens is estimated at approximately 10-15% of patients with rheumatic disease.49 ,51 Recently it was reported that approximately 7% of patients with rheumatoid disease produced an IgE response to the cartilage collagens.52 It was also noted that variations were seen for the ‘autoimmune profiles’ of each patient in as much that different isotypes were generated against different cartilage collagens.52 No studies to date have attempted a detailed serum analysis to obtain a profile of IgG, IgA, IgM, and IgE responses to a broad selection of potential cartilage antigens; but it seems possible that the autoimmune profile may well relate to the extent or prognosis of joint destruction. Whereas IgE autoantibodies are likely to sensitise mast cells to cartilage matrix, possibly eliciting mast cell degranulation, it seems likely that the other antibodies could form immune complexes within the rheumatoid lesion; especially at sites of cartilage erosion where the matrix epitopes would become exposed. Such complexes would be expected to elicit a variety of cellular and immune responses, not least the activation of neutrophils, macrophages, and complement to induce the release of proteases, cytokines, and free radicals.22 This concept of antigen driven mechanisms of joint destruction was initially developed by Poole and colleagues,53 but it is possible that the number of potential epitopes from the numerous cartilage components, and the various immunoglobulin responses, is considerably underestimated.52
Normal cartilage is generally resistant to invasion by a variety of tumour cells and blood vessels, a property originally explained by its high levels of proteinase inhibitors54 and supramolecular architecture of the cartilage surface.55 Thus the observations of cartilage erosion shown here, especially the bidirectional attack via the synovial pannus and subchondral erosion fronts, might suggest that ‘rheumatoid’ cartilage has become deficient in its resistance to cellular invasion.5 This susceptibility might reflect changes in the structure or compositional properties of the cartilage matrix, or changes and exposure of immunogenic epitopes. At present the nature of any subtle changes in the cartilage of affected rheumatoid joints remains unclear, yet much evidence points to RA being a ‘cartilage disease’.
The diagrammatic illustration in figure 7 attempts to summarise our current understanding of the histological observations made from cartilage erosion sites derived from rheumatoid knee joints, admittedly at late stage disease where arthroplasty was the last resort. Although it is possible we may have missed some of the cellular events that characterise early lesions, the broad spectrum of histological features indicates that there was no regular pattern of cells or enzymes that occupy cartilage erosion sites. Numerous factors are likely to regulate the cellular and proteolytic responses, but the generation of cartilage autoantibodies and the formation of immune complexes would seem to provide the underlying basis for the persistence of chronic inflammation and progressive cartilage erosion. The cycle of events depicted in figure 7 would presumably continue until such time as one of the contributors was eliminated or arrested. This hypothesis is borne out to some extent by the total removal of cartilage, which histologically results in quiescent scar formation and a gross reduction in inflammation; and also by pharmacological studies where clinical benefits are claimed by the control of TNFα or IL1,56 ,57 or by the action of anti-inflammatory steroid treatment.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Schematic illustration of potential cellular interactions and cyclical events that may contribute to chronic inflammation and cartilage erosion at the cartilage:pannus junction. Cartilage represented by shaded area. RF, rheumatoid factors; PA, plasminogen activator.
Conclusions
This article describes the microenvironmental nature of cartilage degradation in rheumatoid arthritis with respect to variations in both the distribution of specific cell types and the expression of proteolytic enzymes. It reflects detailed histological analyses of more than 150 different specimens of cartilage:pannus junctions, and subchondral erosion sites, all derived from rheumatoid knee joints. The specific cell types observed at sites of cartilage erosion include macrophages, fibroblasts, mast cells, and PMNs; T cells are rarely seen at the cartilage:pannus interface. Although macrophages and fibroblasts represent the predominant cell types for many junctional specimens, local accumulations of mast cells or PMNs are also observed in discrete microfoci along cartilage:pannus junctions. No regular or consistent pattern has been observed for the cellular invasion/erosion of cartilage during middle or late stage disease, and the observations are best explained as microfocal inflammatory cycles characterised by cellular interchange. As yet the sequence of cellular events, and the hierarchy of factors regulating the production of specific matrix degrading enzymes at one site remain uncertain.
Of special note in approximately 30% of late stage disease specimens was the subchondral erosion of both mineralised and hyaline cartilage. Unlike the cartilage:pannus junction the predominant cells of the subchondral lesion included multinucleated osteoclasts/chondroclasts, macrophages, and small blood vessels. In most specimens chondrocytes showed negligible chondrolytic activity, but when observed in approximately 20% of specimens this was usually restricted to chondrocytes close to the cartilage:pannus junction.
The proteinases involved in cartilage degradation must relate to the cells that occupy erosion sites. Whereas fibroblasts and chondrocytes, when activated, primarily elaborate the matrix metalloproteinases, the serine proteinases are more commonly expressed by granulocytes such as mast cells and PMNs. Multinucleated osteoclasts and chondroclasts, as well as macrophages at subchondral resorptive sites, were commonly characterised by intracellular lysosomal acid phosphatase. Synovial giant cells, although relatively uncommon, characteristically stained for α naphthyl acetate esterase and acid phosphatase. Macrophages and mast cells not only release a variety of degradative enzymes, but also have the potential to produce proinflammatory cytokines and mediators, such as IL1, TNF, histamine, and heparin, which stimulate prostanoid and proteinase expression by neighbouring cells.
The histological studies strongly suggest that both erosive and reparative processes were functional at different sites within the same joint specimen. This microenvironmental matrix remodelling, although eventually resulting in cartilage loss, is probably regulated by locally derived mediators such as cytokines, prostanoids, growth factors, and immune complexes. The autoimmune response is of major importance in the chronicity of RA; the formation of immune complexes between various cartilage antigens and their antibodies of different isotypes may well provide the prime stimulus for the inflammatory and degradative cycles. Such factors would seem essential for the recruitment, maturation, and phenotype of different cell types, and to regulate their catabolic or anabolic functions. Thus it is evident that within the same specimen individual cartilage erosion sites are spatially and temporally separated; with microenvironmental proteinase production, quite probably an intermittent or transient process, being dependent upon a complex sequence of cellular events brought about by locally coordinated signals.
Acknowledgments
Many thanks are due to Michael Bromley for early studies, to Harvey Bertfield, Michael Morris, and Tim Dunningham for the regular supply of surgical material, and to Tracy Bent for preparation of the typescript. Special thanks also to David and Joan Cowie, Langrigg for providing such excellent facilities. This work has been supported by the Arthritis & Rheumatism Council.