Article Text

Abstract
Objectives Rheumatoid arthritis (RA) is associated with an increased Epstein–Barr virus (EBV) blood DNA load, a robust immune response to EBV and cross-reactive circulating antibodies to viral and self-antigens. However, the role of EBV in RA pathogenesis remains elusive. Here, we investigated the relationship between synovial EBV infection, ectopic lymphoid structures (ELS) and immunity to citrullinated self and EBV proteins.
Methods Latent and lytic EBV infection was investigated in 43 RA synovial tissues characterised for presence/absence of ELS and in 11 control osteoarthritis synovia using RT-PCR, in situ hybridisation and immunohistochemistry. Synovial production of anti-citrullinated protein (ACPA) and anti-citrullinated EBV peptide (VCP1/VCP2) antibodies was investigated in situ and in vivo in the severe combined immunodeficiency (SCID)/RA chimeric model.
Results EBV dysregulation was observed exclusively in ELS+ RA but not osteoarthritis (OA) synovia, as revealed by presence of EBV latent (LMP2A, EBV-encoded small RNA (EBER)) transcripts, EBER+ cells and immunoreactivity for EBV latent (LMP1, LMP2A) and lytic (BFRF1) antigens in ELS-associated B cells and plasma cells, respectively. Importantly, a large proportion of ACPA-producing plasma cells surrounding synovial germinal centres were infected with EBV. Furthermore, ELS-containing RA synovia transplanted into SCID mice supported production of ACPA and anti-VCP1/VCP2 antibodies. Analysis of CD4+ and CD8+ T-cell localisation and granzyme B expression suggests that EBV persistence in ELS-containing synovia may be favoured by exclusion of CD8+ T cells from B-cell follicles and impaired CD8-mediated cytotoxicity.
Conclusions We demonstrated active EBV infection within ELS in the RA synovium in association with local differentiation of ACPA-reactive B cells.
- B cells
- Rheumatoid Arthritis
- Autoantibodies
- Infections
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) is a chronic inflammatory disease characterised by production of autoantibodies against rheumatoid factor and anti-citrullinated peptides (ACPA). Although the aetiology of the disease is unknown, complex interactions between genetic and environmental factors play a role in RA development.1 Several observations indicate an association between the B-lymphotropic γ-herpesvirus Epstein–Barr virus (EBV) and RA. Compared with EBV seropositive healthy controls, RA patients have higher levels of EBV DNA and abnormally large numbers of EBV-infected B cells in the peripheral blood,2 ,3 higher titres of serum antibodies to EBV antigens,3–6 and an increased frequency of circulating EBV-specific CD8+T cells.3 ,7–9 Accumulation of CD8+T cells specific for EBV lytic antigen in the synovial fluid of RA patients has also been reported.8 However, the mechanisms linking EBV to RA immunopathology remain elusive.
EBV infects most of the world population and establishes a life-long, generally asymptomatic infection that is kept under tight control by the immune system.10 EBV persists in a latent form in rare B cells, and may undergo reactivation resulting in new infection events and, in certain conditions, EBV-associated cell transformation.11 EBV latency is maintained by expression of a limited set of viral proteins that deliver activation, growth and survival signals to the infected B cells.10 Overexpression of latent EBV protein (ie, LMP2A) can lead to breakdown of B-cell tolerance and rescue of self-reactive B cells,12 ,13 suggesting a possible mechanism for EBV involvement in B-cell-related autoimmune diseases. It has also been shown that antibodies to citrullinated EBV peptides are present in the sera of RA patients raising the possibility that EBV infection may play a role in the induction of ACPA.14 ,15
Some of us recently showed a high frequency of EBV-infected B cells in the brain of patients with multiple sclerosis,16 and in the pathological thymus of patients with myasthenia gravis.17 Ectopic B-cell follicles were identified as main sites of EBV latency and reactivation in the inflamed tissues.16 ,17 Although challenged by other investigators,18–20 these findings suggest that EBV persistence in ectopic lymphoid structures (ELS) could contribute to local immune dysregulation. Previous studies investigating the presence of EBV nucleic acids and/or proteins in the RA synovium have yielded highly heterogeneous results, ranging from undetectable to a large prevalence of EBV infection.21–25 Notably, none of the published studies have investigated EBV infection in the context of different synovial histopathology,26 in particular, presence or absence of ELS. Because ELS are capable of sustaining autoreactive B-cell activation and differentiation,27 ,28 in this study, we tested the hypothesis that EBV infection in RA is intimately associated with synovial ELS and in situ activation of ACPA-producing B cells.
Materials and methods
Patients and synovial tissues
Forty-three RA patients (30 women and 13 men, mean age 64.2 years (range 28–82), mean disease duration 4.5 years (range 1–16), mean C-reactive protein (CRP) 38.7 mg/l (range 5–132), mean erythrocyte sedimentation rate (ESR) 34 (range 5–71), 72% RF+, 58.1% ACPA+) fulfilling the revised 1987 American College of Rheumatology (ACR) criteria for RA29 were recruited in the rheumatology department at Queen Mary University of London. Patients were treated with conventional DMARDs in monotherapy or in combination. Synovial tissue was collected after informed consent (LREC 07/Q0605/29) by ultrasound-guided joint biopsy (n=25), total joint replacement (n=15), or synoviectomy (n=3). In seven patients, multiple biopsies from different joints were obtained. Eleven cases with osteoarthritis (OA) (8 women and 3 men; mean age 72.8 years, range 68–82) undergoing total joint replacement were included as controls.
Characterisation of RA synovial tissues
Histological grading and level of inflammation of RA synovia were assessed by immunohistochemical staining for CD20, CD138, CD3, CD68 and CD21 on paraffin sections as described.27 Samples were classified as diffuse synovitis (n=20), aggregate synovitis (n=18) and aggregate synovitis with follicular dendritic cell (FDC) identifying germinal centre (GC)-like structures (n=12) (see online supplementary figure S1). Synovitis and global inflammation scores were obtained by assigning a semiquantivative score (0–4) to individual immune cell markers30–32 (see online supplementary figure S2 and table S1).
Immunohistochemistry and indirect immunofluorescence
Three µm-thick deparaffinised, or 5 µm snap-frozen, acetone-fixed sections were stained using monoclonal antibodies specific for EBV latent (EBNA1, LMP1, LMP2A) and late lytic (p160, gp350/220) proteins or rabbit polyclonal antibody to the EBV early lytic protein BFRF1, as described16 ,17 (see online supplementary table S2). After antigen retrieval and block of endogenous peroxidase and non-specific binding, paraffin sections were incubated with primary antibodies either overnight at 4°C (BFRF1, EBNA1), or for 60 min (LMP1, p160 and gp350/220) or 90 min (LMP2A) at room temperature (RT) followed by biotinylated secondary antibodies (rabbit antimouse, donkey antirat or goat antirabbit immunoglobulin (Jackson Immunoresearch Laboratories, Suffolk, England) and avidin-biotin peroxidase complex (Elite Vectastain ABC kit, Vector Laboratories, Burlingame, California, USA). The final reaction was visualised with diaminobenzidine (Sigma, St Louis, Missouri, USA) containing 0.01% H2O2, and sections were counterstained with Mayer's hematoxilin (Sigma). Negative controls included omission of primary antibody and incubation with immunoglobulin G (IgG) isotypes or preimmune serum.
For immunofluorescence, deparaffinised synovial sections were incubated overnight at 4°C, or 1 h at RT, with antibodies specific for CD20, CD138, CD8, LMP2A, LMP1, BFRF1 or granzyme B, alone or in different combinations, and then with appropriate fluorochrome-conjugated secondary antibodies (Alexa 488-conjugated goat antimouse (Invitrogen, Eugene, Oregon, USA), TRITC-conjugated donkey or goat antirat, Cy5-conjugated donkey antirabbit or AMCA-conjugated donkey antigoat (Jackson Laboratories)) and sealed in DAPI-containing medium (Invitrogen). All sections were visualised using an Olympus BX60 microscope and epifluorescence. Counts of stained cells were performed in five 40X microscopic fields per section.
EBER in situ hybridisation
In situ hybridisation (ISH) was performed on paraffin-embedded synovial sections from RA and OA patients using the EBV-encoded small RNA (EBER) peptide nucleic acid (PNA) probe/fluorescein and PNA ISH detection kit (DAKO) as previously described.16 An irrelevant probe and a probe for GAPDH (Dako) were used as internal negative and positive controls, respectively. A tonsil from a patient with infectious mononucleosis was used as positive control tissue.
Quantitative real-time RT-PCR
Total RNA was extracted from synovial tissues using the RNeasy Mini Kit (Qiagen, Valencia, California, USA) and treated with RNase-Free DNase (Qiagen, Valencia, California, USA). RNA integrity was checked on 1.2% agarose gel and quantification performed using Nanodrop (Thermo Fisher Scientific, Wilmington, Delaware, USA). For EBV transcripts, 1 μg of RNA was reverse-transcribed with random hexamers using MLV Reverse Transcriptase (Invitrogen Life Technologies, Carlsbad, California, USA). PreAMP Master Mix Kit (Applied Biosystem, Foster City, California, USA) was used to enrich for cellular and viral transcripts as described.33 cDNAs were preamplified in a mix containing forward and reverse primers for GAPDH, CD19, EBNA1, LMP2A, EBER1 and BZLF1 (see online supplementary table S3 for primers and preamplification conditions). RT-PCR was performed as described.33 Values were normalised to GAPDH and calculated using the 2-ΔCt formula. RT-PCR for activation-induced cytidine deaminase (AID) and CXCL13 transcripts was performed as described.27
ACPA-producing cells
Fibrinogen was citrullinated and biotinylated as described.27 Synovial samples were first stained for CD20, CD138 or BFRF1, followed by incubation with the specific secondary antibodies conjugated with ALEXA-488 or ALEXA-555. Biotinylated CFb was then added at 1:20 dilution followed by incubation with streptavidin ALEXA-488 or ALEXA-555. Sections were counterstained with DAPI. Biotinylated, unmodified fibrinogen was used as negative control.
RA synovial tissue transplantation in SCID mice
Sixty synovial tissue samples from four RA patients undergoing arthroplasty (2 ELS− and 2 ELS+) were double-transplanted into 30 beige severe combined immunodeficiency (SCID)-17 mice as described.27 Four weeks post-transplantation animals were sacrificed and underwent a terminal bleed; serum was collected and stored at −20°C for subsequent analysis of human ACPA and anti-EBV antibodies. All procedures were performed according to the Home Office regulations (project licence (PPL) 70/7001).
Detection of human IgG in SCID/RA chimeric mice
Human IgG ACPA were detected in HuRA/SCID sera using an anti-cyclic-citrullinated antibody ELISA kit (Axis Shield, Dundee, UK).27 Human IgG antibodies against VCA, EBNA1 and EA were detected by ELISA (Serion, Würzburg, Germany). Antibodies to viral citrullinated peptides VCP1 and VCP2 were detected as described.15 ,34
Statistical analysis
Differences in quantitative variables were analysed by Mann–Whitney U test, or Kruskal–Wallis with Dunn's post-test, when comparing two or multiple groups, respectively. Fisher's exact test was used to evaluate associations of qualitative variables. All statistical analyses were performed using GraphPad Prism V.3.03 (GraphPad Software, San Diego California, USA). A p value <0.05 was considered statistically significant.
Results
Histological characterisation of RA synovia
Among 50 synovial samples from 43 RA patients, 12 displayed perivascular aggregates with B/T cell segregation and ectopic GC (FDC+), 18 had B-cell aggregates without FDC (these two groups were collectively classified as ELS+), and 20 had diffuse synovitis with sparse or no B cells (classified as ELS−) (see online supplementary figure S1 and table S1). Histological, immunohistochemical, ISH and RT-PCR findings in the present series of RA synovia are summarised in online supplementary table S1.
EBV transcripts and EBER+ cells are preferentially expressed in ELS+ RA synovia
Expression of the EBV latent genes EBNA1, LMP2A and EBER1, and of the immediate early lytic gene BZLF1 was analysed in 13 RA synovial samples (9 ELS+, 4 ELS−) selected for good quality RNA, using preamplification quantitative real-time RT-PCR. ELS+ samples were characterised by higher levels of AID, CXCL13 and CD19 mRNA compared with ELS− samples (figure 1A,B), and also displayed a significantly higher signal for EBER1 and LMP2A (figure 1A–D). EBNA1 and BZLF1 transcripts were not detected. Among four OA synovial samples, all were negative for CXCL13 and EBV transcripts and only the most inflamed case expressed CD19 (figure 1A–D).
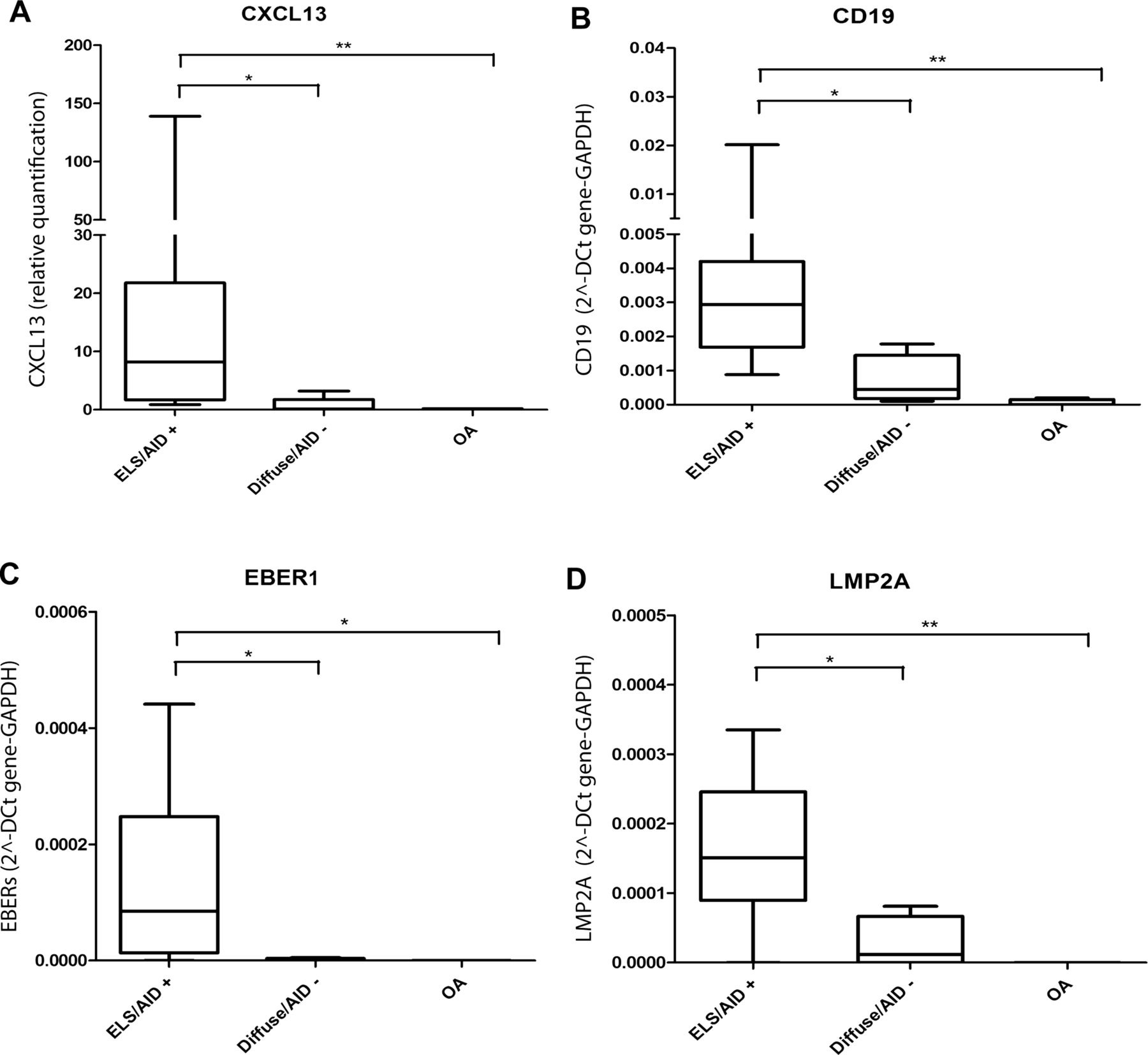
Epstein–Barr virus gene expression in rheumatoid arthritis (RA) and Osteoarthritis (OA) synovial tissue. Quantitative real-time RT-PCR indicates that the level of CXCL13 (A) gene expression is significantly higher in ectopic lymphoid structures (ELS)+/AID+ RA synovia compared with diffuse/AID− RA synovia or OA synovia. Preamplification of selected genes followed by real-time RT-PCR shows that CD19 (B), EBV-encoded small RNA1 (C) and LMP2A (D) gene expression is significantly higher in ELS+AID+ than in diffuse/AID− RA samples. A low level of CD19 mRNA, but no latent viral transcripts, can be detected in OA synovia. *p <0.05, **p <0.01.
Next, we sought to visualise EBV-infected cells using ISH for EBER.35 A tonsil from a case with infectious mononucleosis was used as positive control (figure 2A). Paraffin synovial samples from 19 RA patients (15 ELS+ and 4 ELS−) and eight OA cases were first analysed for GAPDH to assess RNA integrity. Out of 15 GAPDH+ RA samples, 10 displayed EBER signals. Conversely, EBER was not detected in any of the four GAPDH− RA samples and of the eight GAPDH+ OA samples. The highest frequency of EBER+ cells was observed in ELS+ RA samples with GC (n=6), inside and at the border of B-cell follicles (figure 2B–E,G–J), around T-cell areas (figure 2K–M) and in B-cell-rich perivascular cuffs (figure 2N–Q). In ELS+ RA samples without GC (n=4) some EBER+ cells were detected in areas of diffuse B cell/plasma cell infiltration (figure 2R–T). No EBER+ cells were present in four ELS− RA samples. No signal was detected in GAPDH+ samples using an irrelevant probe (figure 2F).

High frequency of Epstein–Barr virus-encoded small RNA (EBER)+ cells in rheumatoid arthritis (RA) synovial samples with ectopic B-cell follicles. EBER in situ hybridisation (ISH) in a tonsil from a patient with infectious mononucleosis (A). CD20 immunostaining and EBER ISH perfomed in serial sections of three different RA synovia with ectopic B-cell follicles (B–E and G,H respectively), reveal numerous EBER+ cells inside (D,E) and at the border of B-cell follicles (G,H). The inset in E shows the nuclear localisation of EBER. ISH with an irrelevant probe on ectopic lymphoid structures (ELS)+ RA synovium (F). EBER ISH (I) and CD138 immunostaining (J) performed in serial sections reveal a high frequency of EBER+ cells in a plasma cell-rich area; this area is marked by a frame in H. CD3 (K) and CD20 (L) immunostainings and EBER ISH (M) performed in serial sections of a ELS+ RA synovium show EBER+ cells accumulating around a T-cell aggregate (marked with an asterisk) that is EBER negative. The inset in M shows EBER+ cells at high-power magnification. Perivascular accumulation of CD20+ and EBER+ cells in serial sections of two different RA synovia with lymphocytic aggregates (N–Q, respectively). Scattered CD20+ cells (R) and EBER+ cells (S) in an area of diffuse immune infiltration in a ELS+/follicular dendritic cell (FDC)− RA synovium. Scattered EBER+ cells in a different ELS+ RA synovium (T). Bars: 200 μm in (B–H), (K and L); 100 μm in (A), (M–O) 50 μm in (I,J) (P–T) 20 μm in the inset in E, 10 μm in the inset in (M).
EBV latent and lytic proteins are exclusively detected in ELS+ RA synovitis
We investigated further the status of EBV infection in a larger set of RA synovial samples (n=38) using immunohistochemistry for EBV latent (EBNA1, LMP1, LMP2A) and lytic (BFRF1, gp350/220, p160) proteins and double immunofluorescence for EBV and B-cell markers. The specificity of anti-EBV antibodies used in this study was previously tested in EBV+ and EBV− tissues and cell lines.16 ,17 ,33
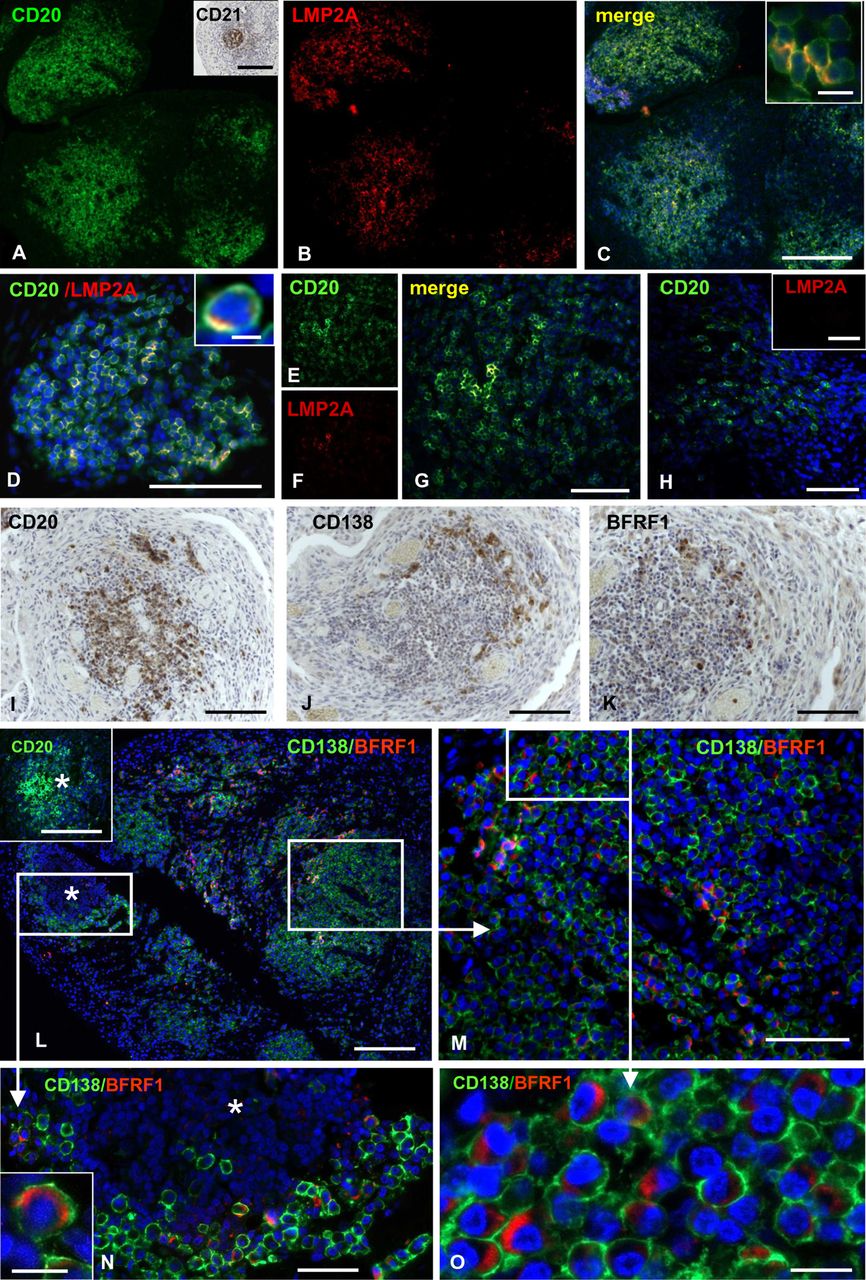
EBNA1, which is expressed during all phases of EBV latency, was detected in an EBV+ lymphoma, but in none of the RA samples analysed (data not shown). LMP2A, a viral membrane protein expressed during the growth and default programmes of EBV latency,10 was found predominantly inside B-cell aggregates in all samples with GC (n=8) and in seven of 14 ELS+ RA synovial samples without GC (figure 3A–G). The percentage of CD20+ B cells expressing LMP2A ranged between 8.2% and 54.7% (mean±SD=19.3±12.8%). All ELS− RA samples were negative for LMP2A (figure 3H). Similarly to LMP2A, LMP1, another viral membrane protein expressed during the growth and default programmes of EBV latency10 was mainly expressed in B-cell aggregates (see online supplementary figure S3A,B).

Detection of Epstein–Barr virus (EBV) latent protein LMP2A and EBV early lytic protein BFRF1 in rheumatoid arthritis (RA) synovia. Double immunofluorescence staining for CD20 (green; A,D,E) and LMP2A (red; B,D,F) performed in ectopic lymphoid structures+ RA synovia with ectopic B-cell follicles reveals expression of LMP2A in intrafollicular B cells (merge; C,D,G). LMP2A+ B cells are scattered throughout the B-cell follicles (A–D), or are confined to the follicle centre (G). In the insets in (C,D) CD20+ B cells coexpressing LMP2A are shown at high-power magnification. The presence of follicular dendritic cell in B-cell follicle is confirmed by CD21 immunostaining (inset in A). CD20+ B cells present in an area with a diffuse inflammatory infiltrate (H) do not display LMP2A immunoreactivity (inset in H). Immunostainings for CD20 (I), CD138 (J) and BFRF1 (K) performed in serial sections of a RA synovial sample with ectopic B-cell follicles reveals that both plasma cells and BFRF1+ cells localise at the B-cell follicle border. Double immunofluorescence staining for CD138 (green) and BFRF1 (red) in a RA synovium with ectopic B cell follicles and massive CD138+ plasma cell infiltration reveals that BFRF1 is expressed in perifollicular plasma cells (L–O). The inset in (L) shows CD20 staining of the B-cell follicle marked with an asterisk. Panels (M,N) show at high-power magnification the perifollcular plasma cell-rich area and the B-cell follicle marked with frames in L. The inset in (N,O) (corresponding to the area marked with a frame in (M)) highlight the typical cytoplasmic localisation of BFRF1. Bars: 200 μm in (C,L) and inset in (L) 100 μm in (D–K), (M) and inset in (H) 50 μm in (N) 10 μm in (O) and insets in (C,N) 5 μm in inset in (D).
BFRF1, a protein associated with the early phase of the EBV lytic cycle36 was detected in all the seven RA synovia with GC analysed. BFRF1+ cells were identified as CD138+ plasma cells (figure 3I–O), and the overall percentage of synovial CD138+ plasma cells expressing BFRF1 in ELS+ samples was 10.1±7.3% (mean±SEM). Of relevance, BFRF1+/CD138+ plasma cells preferentially localised at the periphery of B-cell follicles comprising 29.8±3.1% (mean±SEM) of the perifollicular CD138+ population (figure 4E). Sparse BFRF1+ plasma cells were observed in five out of 14 ELS+ RA samples without GC (see online supplementary figure S3C,D) and none was found in 13 ELS− RA synovia. Expression of the EBV structural proteins gp350/220 and p160 was analysed in six BFRF1+ RA samples with GC. Two small clusters of gp350/220+ cells and p160+ cells were detected in two different samples, respectively (see online supplementary figure S3E,F).

Detection of the Epstein–Barr virus early lytic protein BFRF1 in anticitrullinated protein/peptide antibodies-producing plasma cells in ectopic lymphoid structures+ rheumatoid arthritis (RA) synovia. Double immunofluorescence staining for CD20 (red) and CD138 (green) shows a representative B-cell follicle surrounded by plasma cells (A). At the border of the same B-cell follicle (marked with an asterisk in A,B,D,F) many of the CD138 plasma cells (green in B–D) react with citrullinated fibrinogen (cFb; red in B,C), indicating that they produce APCA. No binding of non-citrullinated Fb is detected (D). In the same perifollicular area, a substantial fraction of CD138+ plasma cells (green) coexpresses BFRF1 (red) (F) the area marked with a frame in (F) is shown at higher magnification in (G) while most BFRF1+ cells (red) are cFb-reactive (green), and vice versa (H–J). The insets in (H,I) highlight the colocalisation of cFb and BFRF1 in the cell cytoplasm. Panels (K–N) show double immunostainings for CD138/cFb (K), CD138/BFRF1 (L), and cFb/BFRF1 (M,N) the area marked with a frame in (M) is shown at higher magnification in (N) in consecutive sections of a different RA synovial sample. The column bar graph in (E) shows the results of cell counts performed in six synovial RA samples. Values are means±SEM of the percentages of double positive CD138+/cFb+ and CD138+/BFRF1+ cells on total CD138+ plasma cells, and of double positive cFb+/BFRF1+ cells on total BFRF1+ and cFb+ cells. Bars: 100 µm in A; 50 µm in (B,D,F,K–M); 20 µm in (C,G,H–J,N) 10 µm in the insets in (K,L) 5 µm in the insets in (H,I,N).
ACPA-producing synovial plasma cells are infected with EBV
Synovial plasma cells associated with ELS in the RA joints produce ACPA.27 Thus, we investigated whether ACPA-producing plasma cells, identified by binding of biotinylated citrullinated fibrinogen (cFb)27 were infected with EBV. Serial sections of eight ELS+/EBV+ RA synovia with GC (allserum ACPA+) and one ACPA+ RA with diffuse synovitis were double-stained for cFb and CD138 (to identify ACPA-producing plasma cells), CD138 and BFRF1 (to identify EBV lytically infected plasma cells), and BFRF1 and cFb (to identify EBV-infected anti-citrullinated protein antibodies (APCA)-producing plasma cells). Six out of eight ELS+/EBV+ RA synovia displayed evidence of plasma cells binding cFb. In these samples, nearly 30% of the CD138+ plasma cells at the periphery of ectopic GC bound cFb, but not unmodified Fb (figure 4A–E,K). Analysis of consecutive sections demonstrated that in the same perifollicular areas, a similar proportion of CD138+ plasma cells coexpressed BFRF1 (figure 4E–G,L). Double immunostainings for cFb and BFRF1 (figure 4H–J,M,N) revealed that most ACPA-producing perifollicular plasma cells (73.7±11.1%, mean±SEM) coexpressed BFRF1 (figure 4E), and that a large proportion of BFRF1+ cells (60.2±5.3%; mean ± SEM) bound cFb (figure 4E). These findings indicate that most plasma cells differentiating in synovial GC and producing RA-associated autoantibodies are infected with EBV.
ELS+ RA synovia engrafted into SCID mice produce antibodies against unmodified and citrullinated EBV antigens
Owing to previous studies having shown higher levels of antibodies specific for unmodified EBV antigens4–6 and presence of antibodies recognising citrullinated EBV peptides14 in the sera of RA patients, we investigated in an in vivo model whether production of such antibodies takes place within the RA synovium and correlates with ACPA production. Hu-RA/SCID mice chimaeras were generated by implanting ELS+ or ELS− RA synovial tissue into SCID mice and presence of circulating synovium-derived human antibodies was analysed after 4 weeks. As previously observed,27 human ACPA (IgG) were significantly higher in sera of mice transplanted with ELS+ RA synovia (figure 5A). ELS+ RA synovial grafts also produced significantly higher levels of IgG against the EBV proteins, EBNA1 and VCA (figure 5B,C), as well as human IgG reacting with EBNA1- and EBNA2-derived citrullinated peptides (VCP1 and VCP2) (figure 5E,F),15 whose level positively correlated with that of ACPA. These findings indicate that citrullinated EBV proteins are recognised by antibodies generated in ectopic GC in the RA synovium.

Unmodified and citrullinated anti-Epstein–Barr virus (EBV) antibodies are locally produced in rheumatoid arthritis (RA) synovia engrafted into SCID mice and correlate with anticitrullinated protein/peptide antibodies (ACPA) production. Synovial specimens from four RA patients were transplanted into 30 SCID mice. Four weeks after transplantation, mice sera were analysed by ELISA for the presence of ACPA and anti-EBV antibodies. Ectopic lymphoid structures (ELS)+ RA synovia engrafted into SCID mice produce significantly higher levels of ACPA (A), anti-EBNA1 (B) and anti-VCA (C) compared with ELS− synovia, while no differences were observed for anti-EA antibodies (D). ACPA+/anti-EBNA1+/anti-VCA+ sera were also analysed for production of antibodies to citrullinated viral antigens VCP1 and VCP2. In vivo production of anticitrullinated EBV antibodies positively correlates with ACPA production (E,F). **p<0.01, *** p<0.001.
Relationship between cytotoxic cells and EBV in RA synovia
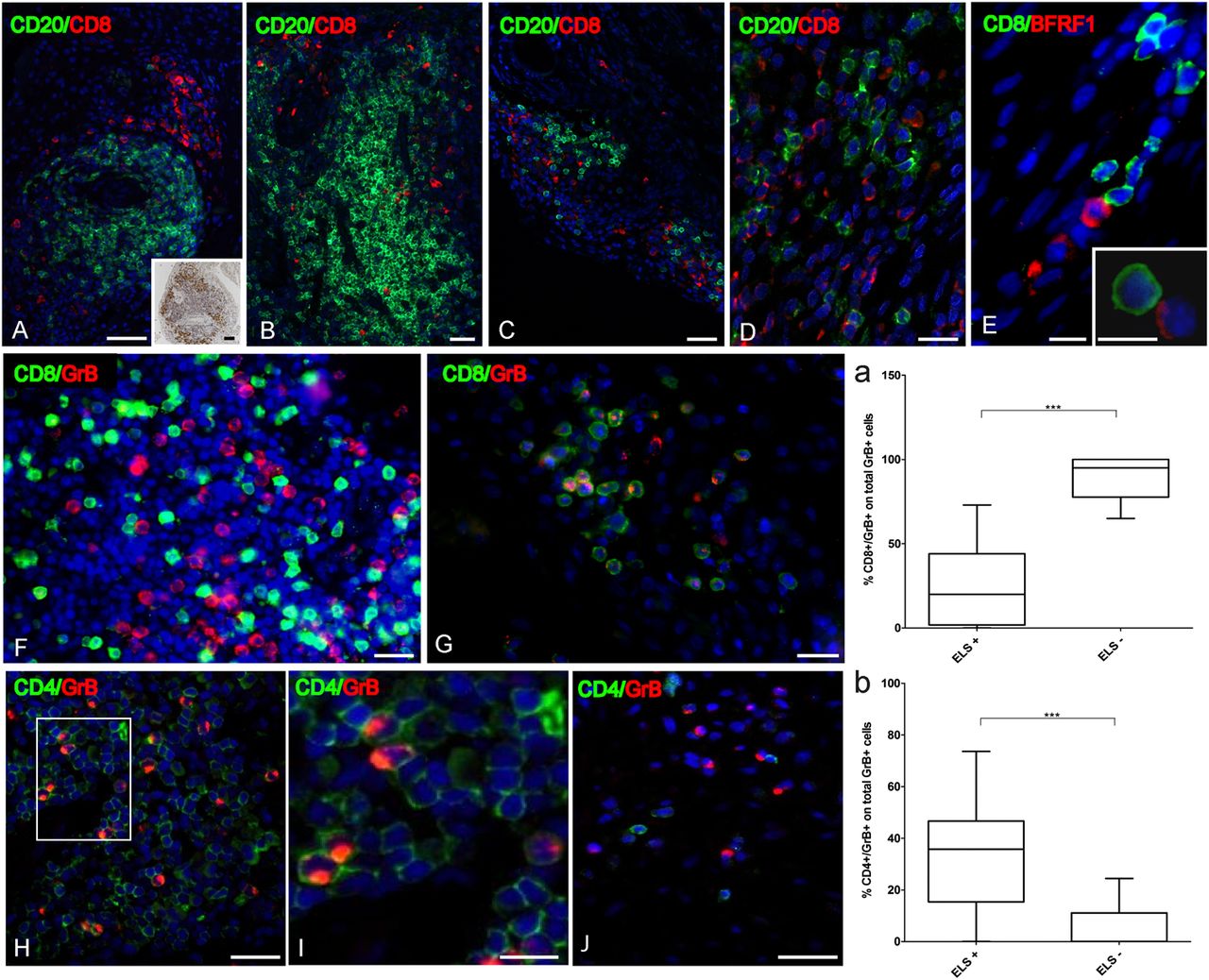
Because CD8+ T cells are essential in the control of EBV infection,37 we investigated the relationship between granzyme B+ CD8+ T cells38 and EBV in 13 RA synovial samples (3 ELS+ FDC+, 2 ELS+ FDC− and 8 ELS−). As reported previously,39 CD8+ T cell infiltration was higher in ELS+ than ELS− RA synovia (61.4±44 cells vs 7.1±8.5 cells counted in five random fields; mean±SEM, p=0.02). However, CD8+ T cells were mostly excluded from synovial B-cell follicles (figure 6A–E). Although more granzyme B+ cells were found in ELS+ versus ELS− RA samples (37.6±28.6 vs 3.9±5.5; mean±SEM, p=0.0001), the percentage of CD8+ cells coexpressing granzyme B was twofold lower in ELS+ than in ELS− RA samples (17.3±4.6% vs 37.3±5.6%; mean±SEM, p=0.02) indicating that most CD8+ T cells infiltrating ELS+ synovia do not display cytotoxic activity, as suggested.40 Accordingly, the percentage of granzyme B+/CD8+ cells in the total granzyme B+ synovial population was significantly higher in ELS− compared with ELS+ RA synovia (87.8±3.6% vs 25.9±5.4%; mean±SEM, p=0.0001; figure 6 graph a, and panels G, H).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Distribution and frequency of CD8+, CD4+ and granzyme B+ cells relatively to Epstein–Barr virus-infected cells in rheumatoid arthritis (RA) synovial. CD8+ cells in ectopic lymphoid structures (ELS)+ RA synovia do not enter large B-cell follicles and mainly colocalise with plasma cells (A,B). The distribution of CD138+ cells is shown in the inset of panel A. In ELS− RA synovia CD8+ cells are found intermingled with B cells (C,D). BFRF1+ and CD8+ cells at the border of a B-cell follicle in ELS+ RA synovium (E). Rarely, CD8+ cells contact BFRF1+ cells (E and inset). CD8+ cells are mainly granzyme B (GrB) negative in ELS+ RA synovium (F). In ELS− RA synovium a high percentage of CD8+ cells expresses granzyme B (G). In ELS+ RA synovium many granzyme B+ cells are CD4+ (H,I). In ELS− RA synovium the majority of CD4+ cells are granzyme B negative (J). Bars: 50 µm in (A–C,H,J) and inset in (A); 20 µm in (D,F,G, and I) 10 µm in the inset in E. The percentage of CD8+/granzyme B+ cells on total granzyme B+ cells is significantly higher in ELS− than ELS+ RA synovia (graph a). The percentage of CD4+/granzyme B+ cells on total granzyme B+ cells is significantly higher in ELS+ than ELS− RA synovia (graph b). ***p<0.001.
A substantial proportion of granzyme B+ cells in ELS+ RA synovia were identified as CD4+ T cells, both inside and outside B-cell follicles (figure 6I,J). CD4+/granzyme B+ cells accounted for 32.8±6.0% versus 5.9±2.1% (mean±SEM, p=0.0003) of total granzyme B+ cells in ELS+ versus ELS− RA synovia (figure 6I–K and graph b), suggesting that in ELS+ RA synovia, a larger subset of CD4+ T cells acquires lytic activity, as previously reported in chronic viral infections, including EBV and RA.41–43
Discussion
It has long been known that EBV infection is perturbed in patients with RA.44 While there is robust evidence of higher EBV load and increased humoral and cellular immune responses to this virus in the peripheral blood of RA patients,2–9 EBV dysregulation in the RA synovium has remained a point of contention.21–25 ,45 Using well-characterised RA synovial biopsies, and combining real-time RT-PCR, ISH and immunohistochemistry to detect EBV RNA and proteins, we consistently found that abnormal EBV infection in the RA joint is associated with ELS, a pathological feature observed in 40–50% of RA patients.46 ,47 Perturbation of EBV infection in ELS+ RA synovia was indicated by a high frequency of infected B cells and plasma cells within and at the periphery of ectopic GC, and by expression of viral genes and proteins that are not usually expressed by EBV-infected B cells in peripheral blood.10 Conversely, EBV was not detected in RA synovia with diffuse immune cell infiltration, which typically lack an established B-cell infiltrate, and in OA synovia. These results provide an explanation for the high variability in the levels of EBV nucleic acids and frequency of EBV-infected cells in RA synovial tissues reported in previous studies which failed to (1) stratify patients according to their synovial pathotype, (2) correlate EBV presence with synovial B-cell infiltration and (3) interpret EBER ISH findings taking into account the degree of RNA preservation of each tissue sample analysed.21–25 ,45
Abnormal EBV infection in synovial B-cell follicles adds RA to the list of autoimmune diseases, that is, multiple sclerosis and myasthenia gravis, in which EBV persistence in pathological tissues has been associated with lymphoid neogenesis,16 ,17 suggesting that EBV colonisation of ELS might be considered a signature of organ-specific autoimmunity. ELS may enable an active EBV infection to be established outside secondary lymphoid tissues where circulating EBV-infected B cells preferentially home, and much of the viral life cycle normally occurs.10 This scenario raises the question of whether EBV is primarily involved in ELS formation through its unique ability to infect and activate B cells,10 ,11 or if ELS-associated EBV dysregulation is secondary to B-cell activation that characterises autoimmune diseases. In either case, the outcome would be the establishment of extralymphatic niches of EBV infection with a direct impact on B-cell function and EBV immune surveillance.
EBV infection in the RA synovium was characterised by expression of both latency and lytic gene expression programmes. The latent EBV transcripts EBERs and LMP2A were detected in all ELS+ RA synovia using real-time RT-PCR, and high numbers of EBER+, LMP1+ and LMP2A+ B cells were found associated with ectopic GC. According to the GC model of EBV persistence, EBV employs four sequentially expressed, virally encoded latency transcription programmes which drive infected naive B cells to become proliferating blasts and participate in GC reactions (where both LMP1 and LMP2A are expressed) in secondary lymphoid tissues, and finally enter the memory B-cell compartment where the virus establishes persistent infection.11 LMP1 and LMP2A possess the signalling properties of CD40 and the B-cell receptor, respectively, and have the capacity to promote the development of infected B cells in the absence of cognate antigen.48 Consistent with the GC model, we show here that both LMP1 and LMP2A are expressed in EBV-infected B cells transiting ectopic GC in the RA synovium, suggesting that these viral proteins may be involved in the abnormal expansion and survival of EBV-infected B cells observed in the pathological tissue. The demonstration that BFRF1, an antigen associated with the early lytic cycle of EBV infection,36 is expressed in a substantial proportion of plasma cells at the edge of synovial GC, and indicates that viral reactivation is initiated as a consequence of differentiation of EBV-infected B cells in these structures. By contrast, EBV structural proteins, like p160 and gp350/220, were rarely detected in synovial ELS suggesting abortive viral replication or very efficient elimination of cells expressing EBV late-lytic antigens.
Because LMP2A has been shown to break tolerance mechanisms and induce autoreactive B-cell activation when constitutively expressed in transgenic mice,12 ,13 EBV infection has been discussed as a biologically plausible mechanism allowing potentially pathogenic autoreactive B-cell clones to escape deletion and cause autoimmunity.49 The RA synovium represents an ideal experimental setting to test this hypothesis, since synovial GC supports the in situ differentiation of ACPA-producing plasma cells.27 ,28 Here, we show for the first time that a remarkable proportion (about 70%) of ACPA-producing plasma cells at the periphery of ectopic GC are infected with EBV, and that about 60% of EBV lytically infected cells in the same areas are ACPA-producing plasma cells. These findings raise the possibility that EBV infection may favour the survival of pathogenic autoreactive B cells that participate in the formation of ectopic GC in the RA synovium. These structures and the local availability of self-antigens likely provide the appropriate environment that allows EBV-infected autoreactive B cells to selectively expand, undergo affinity maturation28 and differentiate into autoantibody-producing plasma cells. The recent finding that EBV persists in self-reactive memory B cells50 lends support to the idea that EBV may contribute to the survival of self-reactive B cells that home to the RA synovium.
Induction of antibodies that cross-react with self-antigens is another possible mechanism through which EBV infection may contribute to autoimmunity.51 ,52 In support of this hypothesis, circulating ACPA in the blood of RA patients recognise citrullinated EBV antigens.14 ,15 Using SCID mice engrafted with ELS+ RA synovia, in which the only possible source of human antibodies is the engrafted tissue, we showed that antibodies against citrullinated EBV antigens (VCP1 and VCP2) are detectable in the serum of transplanted mice and positively correlate with the level of ACPA. This observation demonstrates that potentially cross-reactive antibodies are actively manufactured by GC-like structures in the RA synovium and suggest that post-translationally modified EBV antigens contribute to stimulate intrasynovial humoral immunity, possibly amplifying local inflammation and tissue destruction.
Because EBV latent and lytic proteins are the targets of a cytotoxic immune response,53 the presence of EBV in RA synovia prompted us to investigate whether EBV might contribute to local inflammation through stimulation of a cytotoxic T-cell response. We found that in ELS+ RA synovia cytotoxic CD8+ T cells were largely excluded from EBV+ ectopic GC, raising the possibility that these structures represent immune privileged niches for EBV persistence. Furthermore, the majority of granzyme B+ cells infiltrating ELS+ synovia were CD4+ rather than CD8+, while granzyme B+ CD8+ T cells predominated in ELS− RA synovia. These observations are in keeping with the reported preferential accumulation of non-cytotoxic, IFNγ-producing CD8+ T cells in ELS+ RA synovia,40 and impaired cytotoxic function of EBV-specific CD8+ T cells in RA patients.7 Accumulation of granzyme B+ CD4 T cells in ELS+ synovia is also consistent with studies showing that an uncommon subset of circulating cytotoxic CD4+ T cells is present in chronic viral infections (including EBV) and in RA patients.41–43 ,54 Taken together, these findings can be interpreted as an attempt of CD4+ T cells to compensate a possible defect of CD8+ T cells in controlling increased EBV loads in RA. In this regard, a recent study in humanised mice has shown that EBV infection can induce erosive arthritis suggesting that strong T-cell responses to the virus may play a role in the development of joint pathology.55
In conclusion, our study supports a complex role for EBV in RA pathogenesis through its ability to establish a persistent infection in autoreactive B cells that expand and differentiate in synovial ELS, stimulate production of antibodies to citrullinated and unmodified EBV antigens and, possibly, induce an immunopathological response. The present results significantly advance our understanding of the interplay between EBV infection and establishment of humoral autoimmunity in RA, and will stimulate further studies on the role of altered EBV host interactions in the etiopathogenesis of RA and other conditions associated with ELS.
Acknowledgments
The authors would like to thank Ms Estella Sansonetti for help in figure preparation.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
Footnotes
-
Handling editor Tore K Kvien
-
CC and BS share first authorship; FA and CP share senior authorship
-
Contributors All Authors provided essential contribution to the manuscript and all agree to the current submission.
-
Funding This work was partly funded by the Arthritis Research UK (project grant 18 399 to CP and MB), the William Harvey Research Foundation (to MB) and the Italian Ministry of Health, Ricerca Finalizzata 2007 (to FA and EMC). CC has been recipient of an EMBO short-term fellowship (Ref ASTF 357.00-2010). MB is recipient of an Arthritis Research UK Clinician Scientist Fellowship (grant18 237).
-
Competing interests None.
-
Ethics approval LREC07/Q0605/29, East London The City Research Ethics Committee 3.
-
Provenance and peer review Not commissioned; externally peer reviewed.