Article Text

Abstract
Objective To profile quantitatively the T-cell repertoire in multiple joints and peripheral blood of patients with recent onset (early) or established rheumatoid arthritis (RA) using a novel next-generation sequencing protocol to identify potential autoreactive clones.
Methods Synovium of patients with recent onset (early) RA (<6 months) (n=6) or established RA (>18 months) (n=6) was screened for T-cell clones by sequencing over 10 000 T-cell receptors (TCR) per sample. T cells from paired blood samples were analysed for comparison. From two patients synovial T cells were obtained from multiple inflamed joints. The degree of expansion of each individual clone was based on its unique CDR3 sequence frequency within a sample. Clones with a frequency of over 0.5% were considered to be highly expanded clones (HEC).
Results In early RA synovium, the T-cell repertoire was dominated by 35 HEC (median, range 2–70) accounting for 56% of the TCR sequenced. The clonal dominance in the synovium was patient specific and significantly greater than in established RA (median of 11 HEC (range 5–24) in established RA synovium accounting for 9.8% of T cells; p<0.01). 34% (range 28–40%) of the most expanded T-cell clones were shared between different joints in the same patients, compared with only 4% (range 0–8%) between synovium and blood (p=0.01).
Conclusions In RA, a systemic autoimmune disease, the inflamed synovium forms a niche for specific expanded T-cell clones, especially in early disease. This suggests that, at least in RA, autoreactive T cells should be addressed specifically in the inflamed tissue, preferably in the early phase of the disease.
Statistics from Altmetric.com
Autoreactive T cells are likely to play an important role in many autoimmune diseases but the identification and quantification of these cells in autoimmune tissue inflammation has proved difficult in humans.1,–,3 Identifying these T cells is important to understand their role in autoimmune inflammation, to identify autoantigens, and might help to develop strategies to induce tolerance selectively against autoantigens.
A strong rationale for the involvement of autoreactive T cells comes from genome-wide association studies in autoimmune diseases such as rheumatoid arthritis (RA), autoimmune diabetes (T1DM), systemic lupus erythematosus and multiple sclerosis. These studies all confirmed that the most influential genetic predispositions to these diseases are located in the HLA locus.4,–,7 Furthermore, a pivotal role for autoreactive T cells has been demonstrated in many models of autoimmune diseases such as collagen-induced arthritis, non-obese diabetic mice and experimental autoimmune encephalitis.8,–,10 In several human autoimmune diseases autoreactive T-cell clones have been observed in peripheral blood (T1DM, multiple sclerosis).11 12 However, only few studies have demonstrated the presence of autoreactive T cells in the primary target organ or in its surrounding lymph nodes.13 Therefore, the relationship between autoreactive clones in blood and the inflamed tissue is poorly understood.
Identification of autoreactive T-cell clones in human tissue is hampered by several factors. Most target tissues in autoimmune diseases are difficult to obtain, especially in the early phase of inflammation. Second, in most autoimmune diseases the autoantigens are either unknown (eg, RA, systemic lupus erythematosus) or consist of candidate antigens (T1DM, multiple sclerosis). In this setting many studies focus on the identification of clonally expanded T cells, because this approach does not require knowledge of putative autoantigens. Classically, this approach uses a broad screening method (eg, spectratyping) to find major repertoire aberrations followed by cloning and sequencing of T-cell receptors (TCR) within the aberrations to identify expanded clones. Unfortunately, this approach has a keyhole perspective that has important drawbacks. First, it does not give a full-repertoire perspective. Second, it cannot quantitatively compare the clonal aberrations found across the repertoire. Third, it is prone either to miss clonal aberrations in the first screening step or experience false positive aberrations due to low input.1 14 15
To overcome these limitations we and others recently developed protocols based on next-generation sequencing that provide quantitative data on the degree of expansion of individual clones within the complete TCR repertoire.15,–,18 It thus not only identifies expanded clones, but also obtains detailed information on the impact of these clones on the repertoire as a whole. Furthermore, these protocols for the first time allow quantitative comparisons of clonality between different samples.
Here, we utilise this approach to investigate the TCR repertoire in early autoimmune inflammatory tissue. We selected RA as a model immune-mediated inflammatory disease as it allows for sampling of the primary target tissue, the synovium. Previous studies showed clonal expansions in RA synovitis leading to a ‘skewed’ repertoire compared with peripheral blood.14 19,–,24 Whereas these studies indicate the presence of expanded synovial T-cell clones, their frequency and relation to the total repertoire in synovium and blood are unknown. Using next-generation sequencing to visualise the TCR repertoire, we addressed this challenge by assessing: (1) which part of the repertoire is taken up by expanded clones? (2) does the repertoire change over the course of disease? (3) how does the repertoire in synovium compare with the repertoire in peripheral blood? (4) does the repertoire overlap between different sites of inflamed tissue? and, finally, (5) can we find clones that are shared between different patients?
Materials and methods
Patients
We included six patients with a disease duration of less than 6 months (median 3.5 months, IQR 2.5–4.3 months) who were naive to treatment with disease-modifying antirheumatic drugs and six RA patients with a disease duration of over 1.5 years (median 166 months, IQR 96–274 months) using methotrexate. The first group is denoted as ‘early RA’ patients while the latter is denoted as ‘established RA’ patients. All patients fulfilled the classification criteria for RA, were autoantibody positive (anticyclic citrullinated peptide test >25 kAU/l and/or IgM-rheumatoid factor >12.5 kU/l), had active RA (disease activity score evaluated in 28 joints (DAS28) >3.2) and were naive for treatment with biological agents (see supplementary tables S1 and S2, available online only, for characteristics and MHC typing).25 One additional patient was included for synovial fluid analysis. This patient (no 13) had established active disease despite treatment with rituximab (last infusion was 9 months before sampling). The study was performed according to the Declaration of Helsinki and approved by the local medical ethics committee. All patients gave written informed consent.
Synovial biopsy, peripheral blood and synovial fluid sampling
At inclusion a peripheral blood sample was obtained and an arthroscopic synovial biopsy was performed from a clinically inflamed joint as described previously.26 From three early RA patients who had persistent arthritis after inclusion despite methotrexate treatment (7.5–25 mg/week) additional synovial biopsies were obtained from the same joint after 6 months, when these joints were still inflamed. From one of the patients with established RA synovial biopsies were obtained from both inflamed knee joints within a 10-day period, to study the overlap of clones. From patient no 13 synovial fluid was obtained by arthrocentesis from an inflamed elbow and knee at the same time.
Linear amplification and next-generation sequencing
The linear amplification and next-generation sequencing protocol has been described extensively before.15 27 Details are described in the supplementary methods (available online only).
Statistics
Values are either expressed as mean or median depending on criteria for (non-)normal distribution. Differences between groups were analysed using the Student's t test or Mann–Whitney U tests if appropriate. p Values of less than 0.05 (two-sided test) were considered statistically significant. Graphpad Prism software (version 5.1) was used to perform the analyses.
Results
First we profiled the TCR repertoire in synovium of six patients with early RA (<6 months) who were naive to disease-modifying antirheumatic drugs or biological agents. Per patient more than 10 000 TCR sequences were obtained. We detected multiple clearly expanded clones at the full-repertoire level in all synovium samples in a background of hundreds of less expanded clones (figure 1A). A frequency distribution showed a right skewed distribution in which the majority of the clones (84% of clones; mean, SEM 5.9%) was of low frequency (<0.1% of total TCR analysed; figure 1B). Above this value the distribution decreases quickly towards the x-axis, which it approaches at a clonal frequency of 0.4–0.5%. Therefore, we here defined clones with a frequency of more than 0.5% of the analysed TCR to be highly expanded clones (HEC). Only 8.4% (mean, SEM 4.0%) of the clones was expanded beyond this value. In absolute numbers this corresponded to a median of 35 clones (range 2–70).

Degree of expansion and impact of T-cell clones in synovium of patients with early rheumatoid arthritis. (A) Scatterplot showing all clones recovered from the synovial samples (six patients), in which each dot represents one clone. The size of the clones is depicted as a percentage of the total T-cell receptors (TCR). (B) Frequency distribution of synovial clones showing a right-skewed distribution. Mean and SD are shown. (C) Cumulative size of most expanded clones (mean (black dots) and individual measurements (grey lines) are shown) from six patients. The x-axis depicts the number of clones included (always starting from the most expanded clones). The y-axis shows the percentage of TCR sequences that are attributable to these clones.
Subsequently, we determined the contribution of HEC to the total T-cell repertoire. We took a stepwise approach in which we included an ever increasing number of clones starting with the most expanded one, working our way down through the 250 most expanded clones (figure 1C). Strikingly, we found that the majority of the repertoire was taken up by a small number of HEC. Together, the six most expanded clones accounted for 27% of the T-cell sequences present (median, SD 5.2%), while the 35 most expanded clones (median number of HEC) constituted 56% of the T-cell sequences (median, SD 10.4%). To investigate whether these findings were specific for the synovium, we performed the same analyses on peripheral blood in three of the same patients (see figure 2 and an extensive description of data in the supplementary results, available online only). In blood only 0.08% of the clones had a frequency of more than 0.5%. The impact of the most expanded clones was significantly less compared with synovium.

Degree of expansion and impact of T-cell clones in blood in patients with early rheumatoid arthritis (RA) compared with synovium. (A) Scatterplot showing all clones recovered from peripheral blood in early RA. Each dot represents one clone. The degree of expansion of the clones is depicted as a percentage of the total T-cell receptors (TCR). (B) Enlargement of 0–2% showing that clones are present, but less expanded. (C) Frequency distribution of T-cell clones. Blood (black bars) is compared with synovium (white bars). Mean and SD are shown. *p<0.05 (Mann–Whitney test). (D) Cumulative size of most expanded clones of peripheral blood (black) and synovium (grey) (mean and 95% CI (dashed lines) are shown). The x-axis depicts the number of clones included (always starting from the most expanded clones). The y-axis depicts the percentage of TCR sequences that are covered by the included clones. The impact of the expanded clones is significantly higher in the synovium compared with the peripheral blood for any number of clones up to 125 clones (Student's t test).
Collectively, these findings show that although hundreds of clones were observed, the T-cell repertoire in the inflamed synovium in early RA is dominated by a few HEC only.
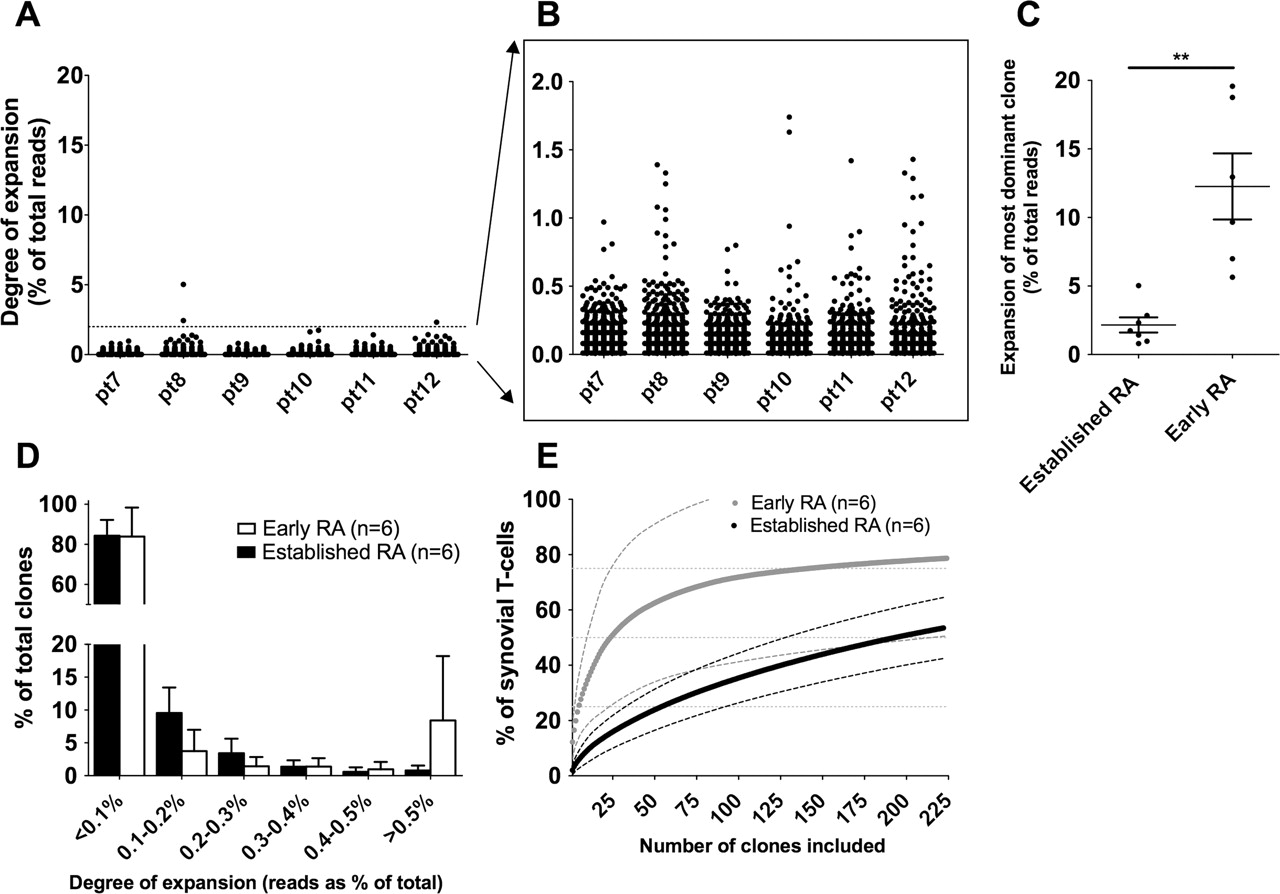
The available literature is undecided about the question of whether prolonged inflammation influences clonality.22 28 29 We therefore analysed the T-cell repertoire in synovium of six patients with ‘established’ RA (disease duration >1.5 years) who also had active disease (mean DAS 7.3, SD 0.8). In these patients we did observe clonal expansions, but the degree of expansion was less compared with that in early RA patients (figure 3A,B). Illustrative was the finding that the median frequency of the most expanded clone was lower compared with synovium of early RA patients (2.2% (SD 0.5) and 12.2% (SD 2.4), respectively; p<0.001; figure 3C). To exclude the possibility that this difference was due to the use of methotrexate in established RA patients, we analysed additional synovial biopsies from three of the early RA patients after 6 months of (ineffective) methotrexate treatment. These additional samples all showed expanded clones with frequencies of 6–27% and were comparable to the samples described in figure 1 (and see supplementary figure S1, available online only).

Degree of expansion and impact of T-cell clones in established rheumatoid arthritis (RA) compared with early RA. (A) Scatterplot showing all clones recovered from peripheral blood in early RA. Each dot represents one clone. The size of the clones is depicted as a percentage of the total T-cell receptors (TCR). (B) Enlargement of clonal size 0–2% showing that clones are present, but that they are less expanded. (C) Comparison of the degree of expansion of the most expanded clones in each sample shows that clones in early RA are significantly more expanded (median shown, Mann–Whitney test, **p<0.01). (D) Frequency distribution of T-cell clones. Early RA (white bars) is compared with established RA (black bars). Mean and SD are shown. (E) Cumulative size of most expanded clones (mean and 95% CI (dashed lines) are shown). The x-axis depicts the number of clones included (always starting from the most expanded clones). The y-axis shows the percentage of TCR sequences that are covered by the included clones. The impact of the expanded clones is significantly higher in early compared with established RA for any number of clones up to 73 clones (Student's t test).
The overall frequency distribution of clones in established RA synovium did not differ significantly from that in early RA, although there was a trend towards a lower number of HEC (mean 0.8% (SD 0.7%) in established RA vs 8.4% (SD 9.7%) in early RA; p=0.11; figure 3D). In established RA this corresponded to a mean of 11 HEC.
Finally, we determined the impact of the expanded clones on the total repertoire (figure 3E). The cumulative size of the 11 HEC corresponded to a mean percentage of the TCR repertoire of 9.8%, which was significantly less compared with the 35 HEC in early RA (56%; p<0.01). When we included the 35 most expanded clones for comparison we could only explain 18.4% of the TCR, which is significantly less compared with early RA (p<0.001). These data show that there are still HEC present during established RA, but the degree of expansion of these HEC is significantly less than in early RA.
Next we investigated whether the HEC overlap between multiple joints of the same patient and between joints and blood. To study overlap between joints we analysed (rare) synovial samples that were obtained from multiple joints at the same time. From one patient synovial tissue was obtained from both inflamed knees within a 10-day interval. From the second patient synovial fluid T cells were obtained from an inflamed elbow and knee joint at the same time. To be able to compare synovium, synovial fluid and blood we arbitrarily chose to study the 25 most expanded clones in each sample. When comparing synovium we observed that 15 (60%) of the top 25 clones from the left knee could also be recovered in the right knee joint (figure 4A). Vice versa nine of the 25 (36%) selected clones from the right knee could be recovered in the left knee. In both knees seven (28%) of the 25 most expanded clones overlapped with the 25 most expanded clones from the other joint. In synovial fluid we observed similar findings; all 25 selected clones (100%) from the elbow could be recovered in the knee and vice versa (figure 4B). In both joints 10 of the 25 most expanded clones (40%) were among the 25 most expanded clones from the other joint.

Overlap of T-cell clones between different joints and between synovium and blood. (A) Overlapping clones in two synovium (ST) samples taken within 10 days from right and left knee respectively in a patient with rheumatoid arthritis (RA). Each dot represents one clone. The frequency (percentage of repertoire) in each knee is depicted on both axes. The lines delineate the frequency of the 25 highest expanded clones; clones depicted above or to the right of these lines represent the clones that are among the 25 highly expanded clones in each knee. Clones in the upper right quadrant represent clones that are found among the 25 highest clones in both knees. (B) Overlapping clones in synovial fluid (SF) taken at the same time from an elbow and knee in a patient with established RA. (C) Representative graph of overlapping clones between peripheral blood (PB) and ST in an early RA patient (n=3 analyses). (D) Representative graph of overlapping clones between PB and ST in an established RA patient (n=4 analysed). (E) Percentage of the 25 most expanded clones that is found in the upper right quadrant of each comparison (ST–ST, SF–SF, ST–PB) *p<0.05.
To compare synovium with blood we studied paired synovium and peripheral blood from three of the early RA patients and four of the established RA patients (figure 4C,D). In this comparison we found that only 4% (median value, range 0–8%) of the 25 most expanded synovium clones could be detected among the 25 most expanded clones in blood, which is significantly less compared with the overlap between joints 34% (range 28–40%; p=0.01; figure 4E). These findings illustrate that there is considerable overlap of highly expanded T-cell clones between different joints, but not between joints and peripheral blood.
Finally, we addressed the question of whether the HEC identified in the synovium showed similarities in their TCR sequence between different patients. Such similarities have been suggested by previous studies, although findings are highly contradictory.30 To this end we compared the TCR sequences of all the synovial HEC detected in this study. We found no clones that were shared by different patients (all 275 CDR3 sequences are shown in supplementary table S3, available online only). Moreover, we found great variation in the use of the (V)ariable and the (J)oining genes within individual patients and between different patients (figure 5). No preferential use of certain VJ combinations could be observed. Therefore, on the sequence and peptide level these findings do not provide evidence of strong similarities of HEC between different patients despite the presence of at least one DRB1 04-allele in at least nine of 12 patients.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Structural similarity in T-cell receptors of highly expanded clones (HEC) derived from different patients. All 275 recovered HEC from all 12 patients with early rheumatoid arthritis (ERA) or established rheumatoid arthritis (ESRA) are shown. A balloon represents each HEC. The x-axis show the V(ariable) gene segment used by the clones, the y-axis the J(oining) gene segment. The balloon is proportional to the degree of expansion of the clone (a full-colour version of the figure is available in the on-line version).
Discussion
We described the findings of a quantitative and sensitive full-repertoire analysis of inflamed tissue in the context of an autoimmune disease (RA). Our observations suggest that the inflamed synovium in multiple joints forms a specific inflammatory niche for expanded T-cell clones especially in the early stages of RA.
Previous studies showed that expanded T-cell clones could be found in the synovium.14 19,–,24 However, both the extent of clonal expansion and the degree of repertoire skewing varied greatly between the studies.30 Overlap of clones between different joints was reported, but the exact relation between clones in different joints and peripheral blood could not be addressed.21 29 Our data show clear oligoclonal expansions in the synovium, with great overlap between different joints. In contrast, the TCR repertoire in peripheral blood is polyclonal and there is hardly any overlap between synovium and blood. In fact, many of the HEC from the synovium could not be recovered from the peripheral blood, suggesting that if they are present in the blood their frequency is lower than 0.01% (lower detection limit). This observation is in accordance with previous studies in T1DM reporting an extremely low frequency of pancreas-specific T cells in the blood (<0.001%) and enrichment for autoreactive T cells in the (T1DM) pancreas compared with the peripheral blood.3 31 These findings imply that analysis of clonality in blood samples might not be informative on the T-cell repertoire in the inflamed synovium. The observed selective oligoclonality in synovium implies that there is highly effective retention and/or proliferation of certain T-cell clones in the synovial niche. The notion that this process is not random is supported by the observed overlap of expanded clones between different inflamed joints. It is tempting to speculate that this retention is caused by the presence of specific (auto)antigens in the synovium.
A striking observation is the difference in the degree of expansion in synovium when comparing early and established RA. Previous literature on such temporal differences in synovial T-cell clonality is conflicting.29 Here, looking at the full repertoire using the quantitative next-generation sequencing approach we observed clear and significant differences. A possible explanation for this temporal difference might be ‘epitope spreading’, in which chronic inflammation leads to neo-epitopes that trigger new autoreactive clones. This phenomenon has been demonstrated in models of autoimmune disease, including models of RA and T1DM.32 In accordance with this model, Monach et al33 recently showed that in synovium of established RA patients many different autoantigens are being recognised by multiple immunoglobulins. If epitope spreading was the leading process behind the differences observed in our study, the HEC in early RA would lose their dominance during chronic inflammation as more clones are being triggered by neo-epitopes. If epitope spreading does take place it is a strong rationale for very early intervention in RA to prevent the disease from spreading. Another theoretical explanation could be that the differences are caused by a change in the presence of T-cell subsets that are known for their vigorous expansions (eg, CD8, CD28null, or CD57+).15 23 34 However, in well-controlled studies such differences between early and long-standing disease have never been detected for phenotypic markers, cytokines or other markers.35,–,37 Future studies might combine immunophenotyping with repertoire sequencing to characterise the HEC further. Finally, although we did not observe reduced clonality after 6 months of methotrexate treatment, the influence of medication cannot be fully ruled out at this point. These effects, and the effects of disease activity itself (both systemically and locally) should be explored in detail in additional (longitudinal) studies.
To estimate the degree of expansion of clones we chose to measure TCR messenger RNA over TCR DNA as an mRNA-based analysis uses 13 times fewer primer combinations and does not require a multiplex PCR amplification. Measuring mRNA as read-out assumes that TCR mRNA levels are equal in different T cells. Although this assumption has been used for many years, it has not been studied extensively. In-vitro studies showed that TCR mRNA levels and TCR surface expression are equal for quiescent CD4 and CD8 T cells (including memory cells). Effector T cells are thought to have at most twofold higher levels of TCR surface expression than quiescent cells, with related mRNA levels.38 39 In our study this means that the estimates of clonal expansions ‘as a percentage of all TCR present’ are likely to be accurate, but estimates on clonal expansions in terms of the number of cells might be slightly inflated up to a factor of two.
The search for autoreactive T-cell clones in autoimmune disease, especially in RA, has proved more difficult than in models of autoimmune disease. The lack of knowledge on both the autoantigens and the autoreactive T cells forms a dilemma. For instance, it has been shown that clones against latent viruses such as Epstein–Barr virus also reside in inflamed synovium.40 41 Therefore, to prove that expanded T cells are truly autoreactive one needs autoantigens, but to find these antigens one needs the autoreactive T cells. Therefore, the advent of new approaches as described in this study may help to identify potential autoreactive clones that subsequently need to be coupled to autoantigens. Our results clearly show that such ‘coupling’ studies (eg, using stimulation approaches) need to take into consideration the fact that there are hundreds of small clones in the synovium. These clones might represent ‘bystander’ clones as they do not overlap between different joints. Therefore, the most sensible approach may be to separate the HEC from the small clones before characterising these clones and interrogating their specificity.
The inflamed tissue niche therefore seems an attractive location to identify and characterise further potential autoreactive clones especially in early RA. This in turn will help to develop novel therapies targeting the underlying autoimmune processes in a more selective and effective way.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Web Only Data - This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Footnotes
-
Contributors All authors fulfilled the International Committee of Medical Journal Editors requirements for authorship/contributorship. All authors made substantial contributions to conception and design, or analysis and interpretation of data, drafting the article or revising it critically for important intellectual content, and final approval of the version to be published.
-
Competing interests None.
-
Funding This work was supported by the IMI JU funded project BeTheCure, contract no 115142-2.
-
Ethics approval The study was performed according to the Declaration of Helsinki and was approved by the local medical ethics committee.
-
Patient consent Obtained.
-
Provenance and peer review Not commissioned; externally peer reviewed.