Article Text

Abstract
Objectives Although myeloid-derived suppressor cells (MDSCs) have been linked to T cell tolerance, their role in autoimmune rheumatoid arthritis (RA) remains elusive. Here we investigate the potential association of MDSCs with the disease pathogenesis using a preclinical model of RA and specimen collected from patients with RA.
Methods The frequency of MDSCs in blood, lymphoid tissues, inflamed paws or synovial fluid and their association with disease severity, tissue inflammation and the levels of pathogenic T helper (Th) 17 cells were examined in arthritic mice or in patients with RA (n=35) and osteoarthritis (n=15). The MDSCs in arthritic mice were also characterised for their phenotype, inflammation status, T cell suppressive activity and their capacity of pro-Th17 cell differentiation. The involvement of MDSCs in the disease pathology and a Th17 response was examined by adoptive transfer or antibody depletion of MDSCs in arthritic mice or by coculturing mouse or human MDSCs with naïve CD4+ T cells under Th17-polarising conditions.
Results MDSCs significantly expanded in arthritic mice and in patients with RA, which correlated positively with disease severity and an inflammatory Th17 response. While displaying T cell suppressive activity, MDSCs from arthritic mice produced high levels of inflammatory cytokines (eg, interleukin (IL)-1β, TNF-α). Mouse and human MDSCs promoted Th17 cell polarisation ex vivo. Transfer of MDSCs facilitated disease progression, whereas their elimination in arthritic mice ameliorated disease symptoms concomitant with reduction of IL-17A/Th17 cells.
Conclusions Our studies suggest that proinflammatory MDSCs with their capacity to drive Th17 cell differentiation may be a critical pathogenic factor in autoimmune arthritis.
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) is a progressive inflammatory autoimmune disease characterised by synovial hyperplasia and bone destruction in affected joints.1 Although the precise aetiology of RA remains unclear, the involvement of CD4+ T cells in RA pathogenesis has been documented.2 Accumulating data show that CD4+ T cells, especially interleukin (IL)-17 producing T helper (Th17) cells, play a significant role in chronic inflammatory diseases including autoimmune arthritis.3 ,4 There is good circumstantial evidence supporting an important role of IL-17 in RA.5 ,6 Studies in experimental arthritis, for example, collagen-induced arthritis (CIA) model7 that recapitulates histopathological changes in human RA, have provided insights into the proinflammatory functions of Th17 cells and IL-17.8–10 While the understanding of Th17 cells has brought new perspective on the treatment of inflammatory diseases,11 ,12 elucidating additional inflammatory mediators in the pathogenic process may lead to new therapeutic management of RA.
Myeloid-derived suppressor cells (MDSCs), a heterogeneous myeloid cell population that is often expanded in tumour-bearing hosts, act as a suppressor of antitumour immune responses.13–15 MDSCs is one of the mechanisms by which cancers escape from immune surveillance or immunotherapy.13–15 They can broadly be characterised as CD11b+Gr-1+ cells in mice16 ,17 and CD11b+CD14−CD33+ in humans.18 Accumulation of MDSCs has also been reported in inflammatory conditions, for example, inflammatory bowel diseases,19 autoimmune hepatitis,20 ,21 experimental autoimmune encephalitis22–25 and experimental autoimmune arthritis26 in mice. However, the involvement of MDSCs in the immunopathology of these diseases, RA in particular, is largely unknown. The question as to why these immunosuppressive MDSCs fail to restrict the disease progression remains unanswered. The association between MDSCs and a proinflammatory Th17 response has also not been examined in autoimmune arthritis.
In this study, we show that expansion of MDSCs coincides with arthritic progression in mice or disease activity in patients with RA. While MDSCs from arthritic mice retain a T cell suppressive capacity, these inflammatory cells are highly efficient in stimulating the Th17 differentiation. Elimination of MDSCs in arthritic mice reduces the disease symptoms as well as the levels of Th17 cells, which could be reversed by adoptive transfer of monocytic MDSCs. Our data support an underappreciated role of MDSCs in promoting tissue inflammation and the pathogenesis of RA.
Results
Expansion of MDSCs during the progression of CIA correlates with tissue inflammation
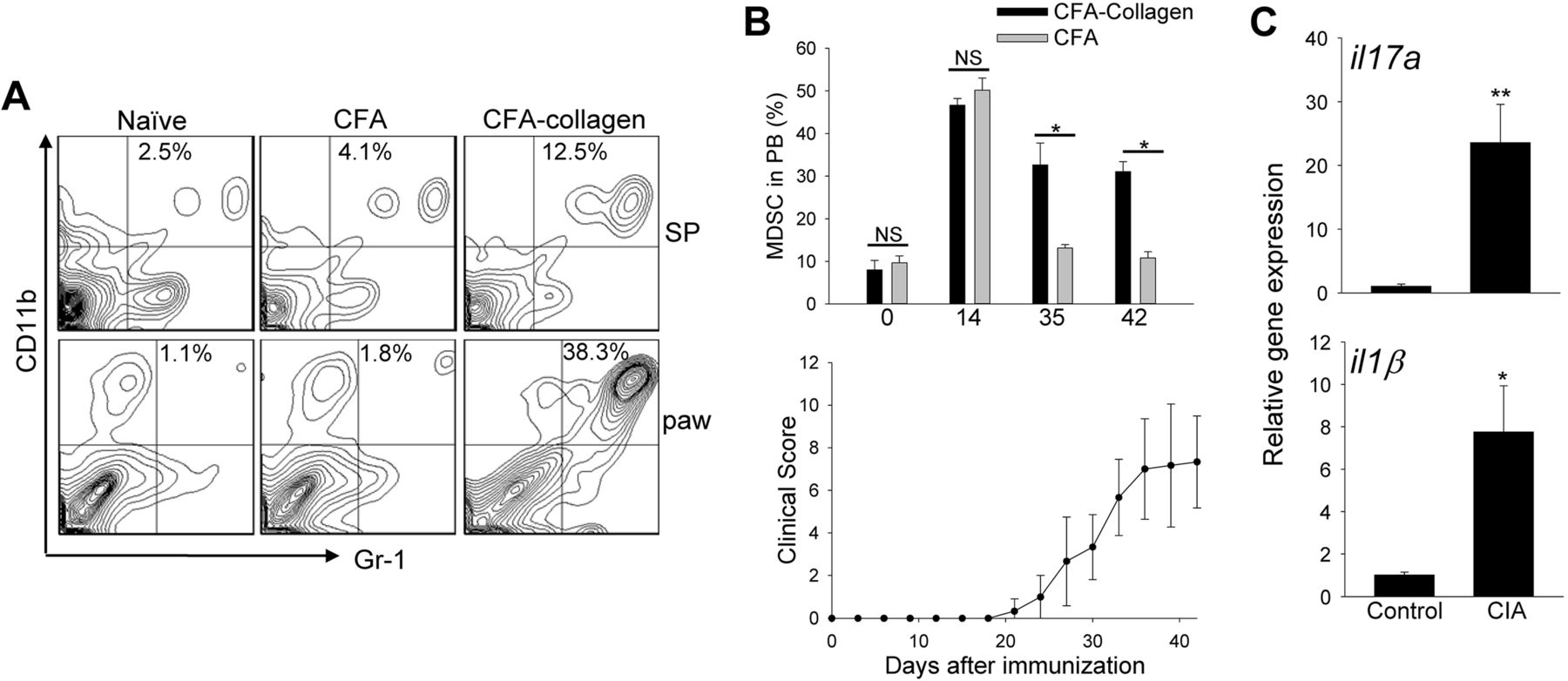
The percentage of CD11b+Gr-1+ MDSCs in arthritic C57BL/6 mice exhibited a marked elevation in the spleen, paws and blood compared with naïve mice or mice immunised with Complete Freund’s Adjuvant (CFA) alone (figure 1A, B). CFA alone induced myeloipoiesis that, however, was not sustained, and CD11b+Gr-1+ cells returned to the basal levels after 4 weeks (figure 1B). The accumulation of MDSCs correlated with increased clinical scores during disease progression (figure 1B). This was associated with upregulation of inflammatory genes in the paws, including il17a, il1β (figure 1C), tnfα, il6, ifnγ, and inos, as well as increased infiltrating Th17 cells (see online supplementary figure S1).

Myeloid-derived suppressor cell (MDSC) expansion is associated with arthritic progression and tissue inflammation. (A) Cells were prepared from spleen (SP) or paws 7 weeks after induction of collagen-induced arthritis (CIA), and stained for CD11b and Gr-1. Naïve mice or mice-treated with Complete Freund's Adjuvant (CFA) alone were used as controls. The representative quantile contour plots from at least three independent experiments are shown. (B) Kinetic changes of MDSCs in blood (upper) of mice after CFA-collagen immunisation correlates with disease progression (lower). (C) The inflammatory genes in the paws were examined by qRT-PCR. The results represent two independent experiments. NS, not significant; *p<0.05; **p<0.001.
MDSCs in arthritic mice display T cell suppressive activity
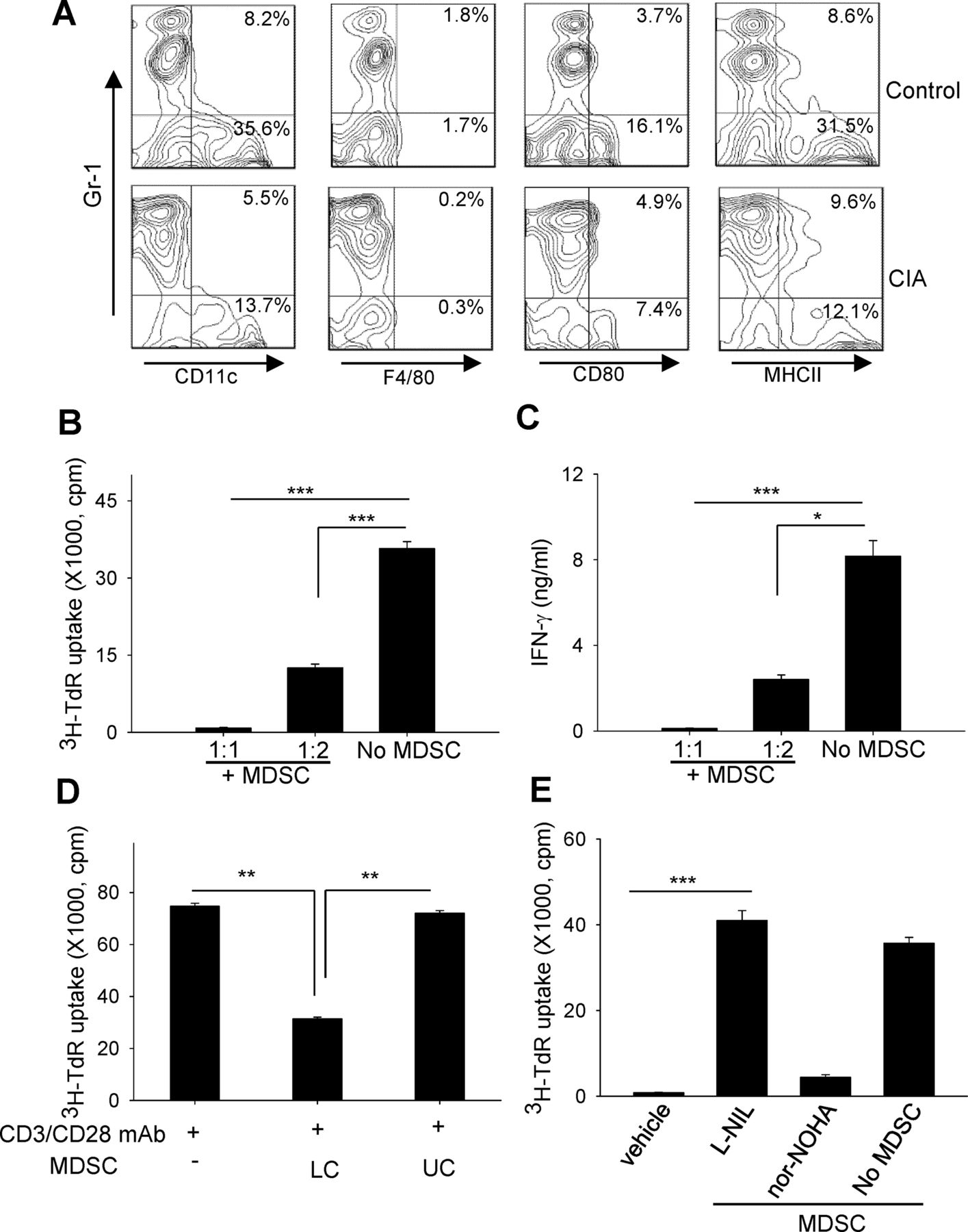
MDSCs in arthritic mice express very low levels of CD11c, F4/80, CD80 and MHCII, which resembles an immature or undifferentiated phenotype (figure 2A). To functionally define this cell population, we examined their ability to suppress T cell activation. A coculture with MDSCs inhibited T cell proliferation (figure 2B) and interferon (IFN)-γ production (figure 2C) stimulated by anti-CD3/anti-CD28 antibodies. However, separating MDSCs from responder splenocytes in Transwell assays rescued T cell activity (figure 2D). The presence of L-N6-(1-iminoethyl)lysine (L-NIL) (an inhibitor of inducible nitric oxide synthase, iNOS), not N-omega-hydroxy-nor-L-arginine (nor-NOHA) (an arginase inhibitor), blocked the suppressive effect of MDSCs (figure 2E), which is consistent with a previously reported role of iNOS in the action of tumour-associated MDSCs.13–15

Myeloid-derived suppressor cells (MDSCs) in arthritic mice suppress T cell activation. (A) Immature phenotype of splenic MDSCs-collagen-induced arthritis (CIA), examined for CD11c, F4/80, CD80 or MHCII after gating on CD11b+ cells. (B and C) MDSCs were cocultured at different ratios with splenocytes in the presence of anti-CD3/anti-CD28 antibodies. T cell proliferation was measured based on 3H-thymidine (TdR) incorporation (B). Interferon (IFN)-γ in the supernatants were assayed by ELISA (C). (D) T cell proliferation was examined using Transwell assays by seeding MDSCs in either the upper chambers (UC) or lower chambers (LC) containing splenocytes. (E) Effect of L-N6-(1-iminoethyl)lysine (L-NIL) or N-omega-hydroxy-nor-L-arginine (nor-NOHA) (500 mM) on MDSC-mediated suppression. *p<0.05, **p<0.01, ***p<0.001.
MDSCs in CIA mice contained two major subsets, CD11b+Ly6ClowLy6G+ granulocytic MDSCs (G-MDSCs) and CD11b+Ly6ChighLy6G− monocytic MDSCs (M-MDSCs) (figure 3A) and confirmed by Wright-Giemsa staining (see online supplementary figure S2). M-MDSCs shared phenotypical markers with inflammatory monocytes, for example, CD115, CCR2 and CD62L (figure 3B); however, M-MDSCs in the paws expressed very low levels of F4/80 (data not shown), implicating their difference from F4/80+ inflammatory monocytes in the infection sites.27

Characterisation of subsets of myeloid-derived suppressor cells (MDSCs) and their correlation with disease severity. (A) CD11b+ splenocytes in arthritic mice were examined for expression of Ly6C and Ly6G. (B) Phenotypical analyses of M-MDSCs after staining for CD115, CCR2 or CD62L. (C) Splenic monocytic MDSCs (M-MDSCs) or granulocytic MDSCs (G-MDSCs) were sorted and their capability to inhibit T cell proliferation (upper) or interferon (IFN)-γ production (lower) was assessed. (D) The frequencies of MDSC subsets and CD4+IL-17A+ Th17 cells in arthritic mice with a clinical score of 1 or 3. Results are representative of three independent experiments. CIA, collagen-induced arthritis; NS, not significant.
To assess the immunosuppressive activity of MDSC subpopulations, we sorted CD11b+Ly6ClowLy6G+ and CD11b+Ly6ChighLy6G− cells from arthritic mice. M-MDSCs were much more efficient than G-MDSCs in inhibiting T cell activation (figure 3C), which agrees with earlier reports on functionally distinct subsets within tumour expanded MDSCs.28 ,29 Similarly, the frequency of M-MDSCs and G-MDSCs correlated positively with the disease severity, as well as the levels of Th17 cells (figure 3D, see online supplementary figure S3A). Analysis of M-MDSCs at either disease onset or peak showed that the inflammatory activation of MDSC-CIA increased during arthritic progression (see online supplementary figure S3B). CFA alone induced M-MDSCs functionally differed from those in arthritic mice, indicated by low expression of TNF-α (see online supplementary figure S3C).
MDSCs in arthritic mice are involved in disease severity and a Th17 response
The failure of MDSCs to prevent the disease progression prompted us to test the possibility of MDSCs as a potential pathogenic factor. We depleted this cell population in arthritic mice (average clinical score of 6) using anti-Gr-1 antibodies (RB6-8C5).30 MDSC depletion alleviated the swelling of paws (figure 4A). Control IgG had little effect on arthritic lesions, evidenced by cartilage erosion of articular surface, synovial hyperplasia and inflammatory exudation in the articular cavity. In contrast, administration of anti-Gr-1 antibodies reversed the pathology of CIA (figure 4B). Removing MDSCs also decreased the frequency of Th17 cells in the spleens (figure 4C) and the il17a gene expression in the paws (figure 4D). Alternative ablation of MDSCs in arthritic mice using gemcitabine, as we previously described,23 also reduced the disease severity and Th17 response (see online supplementary figure S4). Considering the limitation of anti-Gr-1 antibodies that may also deplete neutrophils, we performed an adoptive transfer study using M-MDSCs sorted from CIA mice. Compared with MDSC-ablated mice, M-MDSC transfer resulted in increased disease severity (figure 4E) and incidence as well as elevation of Th17 cells (see online supplementary figure S5).

Depletion of myeloid-derived suppressor cells (MDSCs) reduces disease severity and Th17 response. (A) Arthritic mice (n=10) were treated with control IgG or anti-Gr-1 antibodies on days 25, 28, 31 and 38. **p<0.01, anti-Gr-1 vs IgG. (B) Representative H&E-stained sections of intraphalangeal joints two weeks after antibody treatment. IgG-treated arthritic mice show the fibrovascular synovial and periarticular proliferation (fp), erosion of articular cartilage (ac), and intra-articular exudates (ie). The even and clear joint space (js) and smooth articular cartilage (ac) are seen in arthritic mice treated with Gr-1 antibodies. Magnification, ×100. Bar=50 μm. (C) Reduced frequency of splenic CD4+IL-17A+ cells after MDSC depletion. (D) Diminished expression of il17a gene in the paws after MDSC depletion, as determined by qRT-PCR. **p<0.01. (E) Disease severity enhanced by M-MDSCs. Arthritic mice depleted of MDSCs by gemcitabine (GEM) were adoptively transferred with splenic M-MDSCs sorted from collagen-induced arthritis (CIA) mice. *p<0.05. The results are representative of two independent experiments.
MDSCs in arthritic mice promote the differentiation of Th17 cells
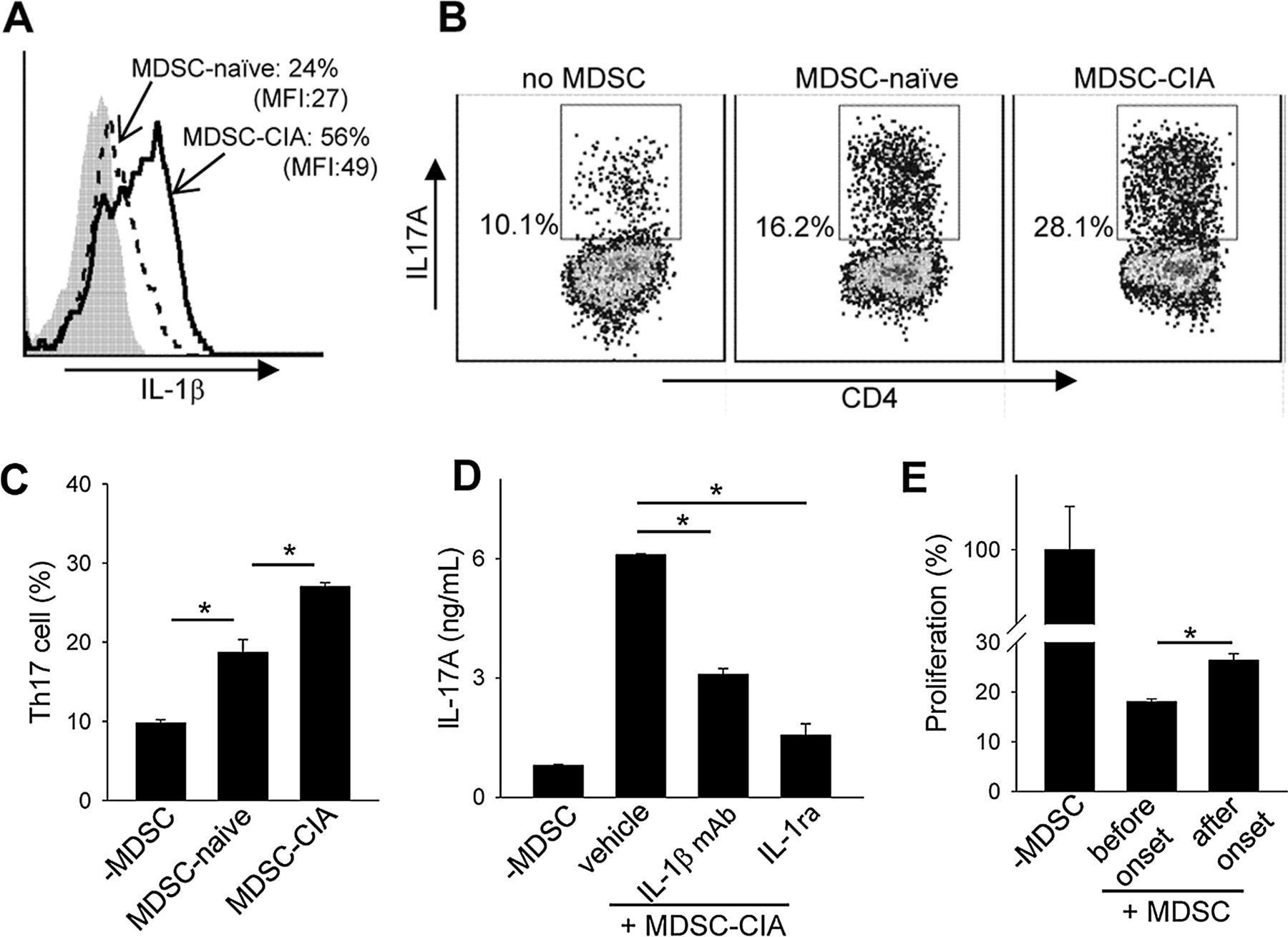
Our recent study in experimental autoimmune encephalitis revealed that MDSCs can facilitate a Th17 response that is dependent on IL-1β signalling.23 Compared with those from naïve mice, MDSCs in arthritic mice (ie, MDSC-CIA) produced higher levels of IL-1β (figure 5A).31 Like TNF-α, increased IL-1β expression in MDSCs was also seen to correlate with CIA progression (see online supplementary figure S6). Coculture of MDSC-CIA with CD4+CD25−CD62Lhigh T cells profoundly increased the differentiation of Th17 cells (figure 5B, C). MDSCs from arthritic mice were notably more efficient than those from naïve mice in this respect. Moreover, blockade of IL-1β signalling using either IL-1β neutralising antibodies or IL-1 receptor antagonist (IL-1ra) diminished MDSC-enhanced production of IL-17A (figure 5D). Intriguingly, M-MDSCs were highly efficient in supporting Th17 differentiation compared with G-MDSCs (see online supplementary figure S7).

Myeloid-derived suppressor cell (MDSCs) from arthritic mice are highly proinflammatory and facilitate Th17 cell differentiation. (A) Increased IL-1β expression by MDSC-collagen-induced arthritis (CIA) compared with MDSC-naïve, assessed by intracellular staining. (B and C) CD4+CD25−CD62Lhigh naïve T cells were cultured in the presence of MDSC-naïve or MDSC-CIA under Th17-polarising conditions. (D) Involvement of IL-1β in MDSC-CIA enhanced Th17 differentiation. IL-17A in the supernatants was examined after Th17 polarisation in the presence of IL-1β antibodies or IL-1R antagonist (IL-1ra). (E) Decreased T cell suppressive activity of MDSCs after disease onset. Splenic MDSCs were isolated 14 days (before onset) or 28 days (after onset) post immunisation and examined for their suppressive activity.
To examine whether the immunosuppressive activity of MDSCs might alter during the disease progression, we isolated MDSCs at different stages (ie, day 14 vs day 28) after collagen immunisation and compared their relative T cell suppressive activities. We found that MDSCs harvested after disease progression were significantly weaker than those from mice before disease onset (figure 5E). Co-transfer of MDSCs from different disease stages with antigen-specific T cells further confirmed a reduced suppressive activity of MDSCs in vivo during CIA progression (see online supplementary figure S8).
Human MDSCs correlate with disease activity and levels of Th17 cells in patients with RA
Compared with healthy donor controls, human CD11b+CD33+ MDSCs increased significantly in patients with RA with high disease activity but remained unchanged in those with low disease activity (figure 6A, B). The increase of these cells was also seen in the synovial fluids of patients with RA when compared with patients with osteoarthritis (figure 6C, D). Additionally, the frequency of human MDSCs in the synovial fluids of patients with RA positively correlated with the levels of IL-17A (figure 6E). Similar to mouse MDSCs, MDSCs from patients with RA also showed the capability to enhance the differentiation of human Th17 cells in vitro (figure 6F, G).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Elevation of myeloid-derived suppressor cells (MDSCs) correlates with disease activity and Th17 response in patients with rheumatoid arthritis (RA). (A and B) Increased frequency of CD11b+CD33+ blood MDSCs in high disease activity (28-joint disease activity score, DAS28>5.1) patients compared with low disease activity (DAS28≤5.1) patients or healthy controls. The representative flow chart (A) and the statistical analysis (B) are shown. (C and D) Increased MDSCs in synovial fluids from patients with RA compared with patients with osteoarthritis (OA). (E) Positive correlation between the frequency of MDSCs and the level of IL-17A in synovial fluids of patients with RA (n=18). (F and G) MDSCs from patients with RA promote Th17 cell differentiation. CD4+CD25− T cells were cultured for 6 days with or without MDSCs isolated from blood of patients with RA. The representative histogram (F) and the statistical results (G) are shown. The data are representative of five independent experiments.
Discussion
Our study of MDSCs in arthritic mice and patients with RA elucidates an association of MDSC expansion with disease severity and Th17 response. The capacity of MDSCs from arthritic mice or patients with RA to efficiently promote Th17/IL-17 suggests a pathogenic role of MDSCs in RA.
Analyses of the frequency of MDSCs in lymphoid tissues and inflamed paws of arthritic mice clearly establish a positive correlation between MDSC accumulation, Th17 cells and disease severity. The ability of these MDSCs to inhibit T cell activation ex vivo in a cell contact-dependent and iNOS-dependent fashion defines an immunosuppressive characteristic retained in these cells.14 ,20 ,22 Our data also show that MDSCs from arthritic mice express higher levels of inflammatory cytokines (eg, TNF-α, IL-1β) than those from corresponding naïve mice or those treated with CFA alone, supporting an inflammatory activation in these cells. Moreover, MDSCs from arthritic mice are highly efficient in facilitating the Th17 differentiation, indicating their proinflammatory nature. Promoting Th17 polarisation by MDSCs from arthritic mice appears to be mediated, at least in part by, MDSC-derived IL-1β. Using complementary approaches (ie, cell depletion, cell transfer) we further demonstrate that MDSCs or M-MDSCs are directly involved in arthritic progression and inflammatory Th17 response. Recently, human Th17 cells were reported to be driven by nitric oxide (NO)-producing MDSCs associated with endogenous iNOS signalling.32
Despite the fact that MDSCs from arthritic mice retain an immunosuppressive activity, the mobilised MDSCs fail to limit the immunopathology. We believe that excessive expansion or persistent accumulation of these cells amplifies tissue inflammation by producing inflammatory factors and facilitating a Th17 response, as we recently reported in the experimental model of multiple sclerosis.23 Indeed, MDSCs show reduced T cell suppressive activity during the arthritic progression. Overriding the intrinsic suppressive property by a proinflammatory feature in MDSCs would result in a detrimental effect, for example, exacerbation of inflammation, bone damage.
The current work also shows that circulating MDSCs elevate significantly in patients with RA with high disease activity compared with healthy donors or those with low disease activity. The positive correlation between MDSCs infiltration and IL-17A levels in synovial fluid of patients with RA and the ability of human MDSC to facilitate Th17 differentiation support the notion that MDSCs may play a critical role in autoimmune arthritis. Interestingly, the frequency of MDSCs in the peripheral blood appears to be negatively associated with Th17 cells in human RA (Hu et al, unpublished), which is consistent with a recent report.33 The reason for this is unclear and more studies are necessary to elucidate the ‘functional’ interaction between MDSCs and Th17/IL-17 in different sites.
Our finding is in contrast with a recent study by Fujii et al26 reporting a protective role of MDSCs in arthritic DBA/1 mice. This discrepancy may be due to the phenotypically distinct MDSC subsets expanded in mice of different genetic backgrounds. In our study M-MDSCs and G-MDSCs are expanded in arthritic C57BL/6 mice, whereas only G-MDSCs were seen upon CIA induction in DBA/1 mice. Considering the extremely high levels of MDSCs in arthritic mice (over 30% in blood), we perform MDSC depletion prior to transfer of M-MDSCs sorted from arthritic mice,23 as opposed to a direct cell transfer procedure in the other study. The disease progression accelerated by M-MDSCs, which are also highly efficient in promoting Th17 response/differentiation in our study, further supports its pathogenic effect.
In summary, our findings in an experimental arthritic model and patients with RA suggest that MDSCs are an inflammatory factor capable of supporting the Th17 differentiation. Although additional studies are needed to further define the precise role of MDSCs, we propose that MDSCs play an important role in RA, and therapeutic approaches directed against MDSCs may lead to alleviation of disease.
Methods
Mice
C57BL/6 8–10 week-old male mice were obtained from the National Cancer Institute (Bethesda, Maryland, USA). All procedures involving mice were approved by the Institutional Animal Care and Use Committee of Virginia Commonwealth University.
Patient samples
Whole blood samples were collected from patients with RA (n=35), including 22 high disease activity (28-joint disease activity score, DAS28>5.1) patients and 14 low disease activity (DAS28≤5.1) patients, or healthy controls (n=16). Synovial fluid samples were obtained from patients with osteoarthritis (n=15) or RA (n=23). The study protocols and consent forms were approved by the Institutional Medical Ethics Review Board of Peking University People's Hospital.
Induction of experimental arthritis
Arthritic induction in mice was performed according to the protocol previously described.34 Briefly, chicken type II collagen (4 mg/mL) was dissolved in 0.05 M acetic acid and emulsified in an equal volume of CFA containing 4 mg/mL heat-killed Mycobacterium tuberculosis H37 (Difco laboratories, Detroit, Michigan, USA). Mice were subjected to a single intradermal injection at the base of the tail. The severity of arthritis was scored as described.35 To deplete MDSCs, anti-Gr-1 antibodies (100 μg) were administrated on days 25, 28, 31 and 38. For adoptive cell transfer, mice were immunised with CFA-collagen and treated on days 6 and 10 with gemcitabine (100 mg/kg).23 On days 14 and 21 mice received M-MDSCs sorted from arthritic mice.
Flow cytometry analysis
Single cell suspensions were incubated with anti-CD16/-CD32 antibodies for 20 min on ice, followed by staining with fluorescently labelled antibodies for surface markers (see online supplementary materials and methods) and analysed on a FACSCalibur.
MDSC isolation and T cell suppression
MDSCs were isolated from the spleens of CIA mice with clinical score higher than 6 unless indicated using CD11b+ magnetic beads (Miltenyi Biotec, Auburn, California, USA).23 In some experiments, M-MDSCs or G-MDSCs were isolated using a FACSAria II cell sorter. T cell suppression assays were performed as we previously described.23 For Transwell experiments, MDSCs were seeded to the upper or the lower chambers, while splenocytes were plated in the lower wells with anti-CD3/anti-CD28 antibodies.
Th17 cell differentiation
CD4+CD25−CD62Lhigh naïve T cells were cultured with MDSCs under Th17-polarising conditions as we described.23 In some experiments 30 μg/mL IL-1β antibodies or 200 ng/mL IL-1 receptor antagonist were used. For human Th17 cell differentiation, 5×105 CD11b+CD33+ MDSCs and CD4+CD25− T cells were cocultured at the ratio of 1 : 1 in the presence of plate bound anti-CD3 (3 µg/mL), anti-CD28 (3 µg/mL), anti-IFN-γ (1 µg/mL), and anti-IL-4 (2.5 µg/mL) (eBioscience, San Diego, California, USA), TGF-β1 (2.25 ng/mL), IL-6 (30 ng/mL), IL-1β (20 ng/mL) and IL-23 (30 ng/mL) (Perprotech, Rocky Hill, New Jersey, USA).
Intracellular IL-17A staining
Splenocytes were stimulated with denatured collagen (50 μg/mL) for 48 h and treated with phorbol myristate acetate (10 nM) plus ionomycin (1 mM) for 6 h as previously described.23 Brefeldin A (10 µg/mL) was added for the last 3 h of culture before cell permeabilisation and staining with anti-IL-17A antibodies. Cells prepared from the paws or synovial fluids were used without collagen stimulation.
Quantitative PCR
Expression of inflammatory genes was quantified relative to that of β-actin using an ABI prism 7900HT System, and normalised to that measured in controls by standard 2(−ΔΔCT) calculation.23 The probe sets are listed in online supplementary materials and methods.
Statistical analysis
Data are presented as means±SE. Student's t test or analysis of variance (ANOVA) was used for statistical analysis. p Values less than 0.05 were considered statistically significant.
Acknowledgments
The authors thank Ms Annicole Buranych for her editorial assistance.
References
Supplementary materials
Supplementary Data
This web only file has been produced by the BMJ Publishing Group from an electronic file supplied by the author(s) and has not been edited for content.
Files in this Data Supplement:
- Data supplement 1 - Online supplement
- Data supplement 2 - Online figures
Footnotes
Handling editor Tore K Kvien
CG, FH, HY, and ZF contributed equally.
Contributors CG, FH, HY and X-YW designed experiments. CG, FH, HY, ZF, CL, LS, YL, HL, XY and HW performed experiments; CG, FH, HY, JL and ZL analysed data. ZL and X-YW supervised the research. CG, FH and X-YW wrote the manuscript.
Funding The present study was supported in part by National Institutes of Health (NIH) Grants CA175033 and CA154708 (X-YW), the Natural Science Foundation of China (81302554 to FH, 81030057 to ZL and 81173456 to JL), 973 Program of China (2010CB529100, ZL), and Peking University People's Hospital Research and Development Fund (RDB2013-04, FH). Flow cytometry facility was supported in part by NCI Cancer Center Support Grant to VCU Massey Cancer Center P30CA16059. X-YW is the Harrison Endowed Scholar in Cancer Research.
Competing interests None.
Patient consent Obtained.
Ethics approval Institutional Medical Ethics Review Board of Peking University People's Hospital.
Provenance and peer review Not commissioned; externally peer reviewed.