Article Text

Abstract
Background: Antibodies directed to proteins containing the non-standard amino acid citrulline, are extremely specific for rheumatoid arthritis (RA). Peptidylcitrulline can be generated by post-translational conversion of arginine residues. This process, citrullination, is catalysed by a group of calcium dependent peptidylarginine deiminase (PAD) enzymes.
Objective: To investigate the expression and activity of four isotypes of PAD in peripheral blood and synovial fluid cells of patients with RA.
Results: The data presented here show that citrullination of proteins by PAD enzymes is a process regulated at three levels: transcription—in peripheral blood PAD2 and PAD4 mRNAs are expressed predominantly in monocytes; PAD4 mRNA is not detectable in macrophages, translation—translation of PAD2 mRNA is subject to differentiation stage-specific regulation by its 3′ UTR, and activation—the PAD proteins are only activated when sufficient Ca2+ is available. Such high Ca2+ concentrations are normally not present in living cells. In macrophages, which are abundant in the inflamed RA synovium, vimentin is specifically citrullinated after Ca2+ influx.
Conclusion: PAD2 and PAD4 are the most likely candidate PAD isotypes for the citrullination of synovial proteins in RA. Our results indicate that citrullinated vimentin is a candidate autoantigen in RA.
- rheumatoid arthritis
- peptidylarginine deiminase
- citrullination
- macrophages
- vimentin
- ACR, American College of Rheumatology
- anti-MC, anti-modified citrulline antibodies
- CCP, cyclic citrullinated peptide
- HUGO, human genome organisation
- NP-40, Nonidet P40
- PAD, peptidylarginine deiminase
- PB, peripheral blood
- PBMCs, peripheral blood mononuclear cells
- PMCA, plasma membrane Ca2+ pump
- RA, rheumatoid arthritis
- RT-PCR, reverse transcriptase-polymerase chain reaction
- SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis
- SF, synovial fluid
- SFMCs, synovial fluid mononuclear cells
- UTR, untranslated terminal region
Statistics from Altmetric.com
- ACR, American College of Rheumatology
- anti-MC, anti-modified citrulline antibodies
- CCP, cyclic citrullinated peptide
- HUGO, human genome organisation
- NP-40, Nonidet P40
- PAD, peptidylarginine deiminase
- PB, peripheral blood
- PBMCs, peripheral blood mononuclear cells
- PMCA, plasma membrane Ca2+ pump
- RA, rheumatoid arthritis
- RT-PCR, reverse transcriptase-polymerase chain reaction
- SDS-PAGE, sodium dodecyl sulphate-polyacrylamide gel electrophoresis
- SF, synovial fluid
- SFMCs, synovial fluid mononuclear cells
- UTR, untranslated terminal region
The blood of patients with rheumatoid arthritis (RA) contains large numbers of circulating antibodies directed against a variety of autoantigens. The most widely known are the rheumatoid factor (RF), antibodies directed against the Fc part of IgG molecules (reviewed by Mageed1). RF can be detected in roughly 75% of patients with RA, but is also present in patients with other rheumatic or inflammatory diseases and even in a substantial percentage of the healthy (elderly) population.2 Its specificity for RA is thus limited.
Autoantibodies directed against citrullinated proteins have a much higher specificity for RA (reviewed by van Boekel et al3). This family of autoantibodies comprises the antiperinuclear factor,4 the so-called “antikeratin” antibodies,5 antifilaggrin antibodies,6,7 and anti-cyclic citrullinated peptide (anti-CCP) antibodies.8,9 These autoantibodies all recognise epitopes containing the non-standard amino acid citrulline; the naming of the antibody is simply determined by the method used to detect them. With the most sensitive assay (CCP2), anticitrullinated protein antibodies can be detected in almost 80% of RA sera with a specificity of 98%.3,9–11 Besides being very specific for RA,9,10,12,13 anti-CCP antibodies can be detected very early in the disease and can predict clinical disease outcome,14–17 making them a useful tool for the clinician.
The relative amounts of anticitrullinated protein antibodies in synovial fluid (SF) and in extracts of synovial tissue of patients with RA is 1.4- and 7.5-fold higher, respectively, than in sera of the same patients.18,19 Furthermore, anticitrullinated protein antibody secreting cells are found in the inflamed RA synovia.19,20 These data suggest the presence of citrullinated proteins in the RA synovium causing an antigen driven maturation of CCP-specific B cells at the site of inflammation. Indeed, the presence of citrullinated proteins in the RA synovium (for example, fibrin) has already been reported.21
Because citrulline is a non-standard amino acid, it is not incorporated into proteins during translation. It can, however, be generated by post-translational enzymatic deimination (citrullination) of arginine residues.22 This conversion is catalysed by the enzyme peptidylarginine deiminase (PAD, EC 3.5.3.15). Therefore, when citrullinated proteins are present in the inflamed RA synovium,21 PAD enzymes must also be present. Several isotypes of PAD exist, each with a different tissue distribution (reviewed by Vossenaar et al23). PAD1 and PAD3 are mainly found in epidermis and hair follicles.24–27 PAD2 is expressed in a variety of tissues, including muscle, brain, and haematopoietic cells,28–30 whereas the human PAD4 (formerly known as PAD5) is found primarily in haematopoietic cells.31–33
The RA synovial membrane contains large numbers of activated macrophages34; cells that have been described as expressing PAD enzymes and, under certain conditions, containing citrullinated proteins.35,36 This makes them good candidates for the generation of synovial citrullinated antigens.
Here we describe the mRNA and protein expression of PAD enzymes in monocytes and macrophages, as well as in synovial fluid mononuclear cells (SFMCs). Possible mechanisms for regulation of PAD expression and activity are discussed.
MATERIALS AND METHODS
Patients and controls
Blood samples from patients matching the American College of Rheumatology (ACR) criteria for RA37 were collected with informed consent from the Department of Rheumatology of the University Medical Centre, St Radboud in Nijmegen. Paired samples of SF and peripheral blood (PB) from patients matching the ACR criteria for RA37 were collected with informed consent from the Department of Rheumatology of the Sint Maartenskliniek in Nijmegen. Blood samples from healthy people were collected from volunteering blood donors. All blood and SF samples were processed immediately to ensure optimal cell preparations.
Cell isolation, culture, and calcium treatment
Heparinised blood (15 ml) was diluted 1:1 with phosphate buffered saline (PBS) citrate (PBS containing 0.455% Na3 citrate) and layered over a 15 ml Ficoll cushion. After centrifugation for 20 minutes at 500 g, the peripheral blood mononuclear cell (PBMC) containing interphase was collected and washed two times with PBS citrate. From here on PBMCs were fractionated either by plastic adherence, or with MACS magnetic beads (Miltenyi Biotec). For MACS fractionation, beads coated with either anti-CD3, anti-CD14, anti-CD19, or anti-CD56 antibodies were used according to the manufacturer’s instructions. For plastic adherence, PBMCs were cultured for 60 minutes in RPMI-1640 medium (with glutamax-I, Dutch modification, Gibco BRL) containing 10% pooled human serum, 1 mM pyruvate, and 50 μg/ml gentamicin (Gibco BRL) in six well culture plates, allowing monocytes to adhere to the plastic. The non-adherent lymphocytes were removed by washing the culture plates with PBS, after which the adherent monocytes were either harvested directly or allowed to differentiate into macrophages in seven days. This combined Ficoll centrifugation/plastic adherence protocol allows for >90% pure cell preparations.38 Mononuclear cells from SF were isolated by the same procedure. To increase cytosolic Ca2+ concentrations, cells were treated with either 1 μM ionomycin (Sigma) or 1 μM thapsigargin (Sigma) in Locke’s solution (150 mM NaCl, 5 mM KCl, 10 mM Hepes.KOH pH 7.3, 0.1% glucose) in the presence of 2 mM extracellular calcium for 15 minutes up to 18 hours.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
RNA was isolated from cells using Trizol reagent (Gibco BRL) according to the manufacturer’s instructions. Synthesis of cDNA from 1 μg of RNA was performed using the reverse transcription system (Promega) with 500 ng of random primers. The reaction mixture was incubated for 10 minutes at room temperature, followed by 90 minutes at 42°C. After incubation, cDNA samples were diluted with distilled water to 100 μl. For RT-PCR 10 μl cDNA was added to 40 μl mastermix containing 75 mM Tris.Cl pH 8.8, 20 mM (NH4)2SO4, 0.01% (vol/vol) Tween-20, 2 mM MgCl2, 0.2 mM dNTPs, 0.5 μM forward and reverse primer, and 1 U Red Hot DNA polymerase (ABGene). PCR was performed using a T3 thermocycler (Biometra): two minutes at 94°C, 25–35 cycles of (30 s, 94°C; 30 s, 56–60°C; 15–30 s, 72°C), two minutes at 72°C. PCR product (15 μl) was analysed on a 1.5% agarose gel using the Geldoc (Biorad) system. The following primer pairs were used (the size of the amplicon is given in parentheses): β-actin (239 bp) 5′-ACTCCATCATGAAGTGTGACG-3′ and 5′-CATACTCCTGCTTGCTGATCC-3′; PAD1 (458 bp) 5′-CCACAGCTGGCTGATG-3′ and 5′-TCTCATTGGAGCCATGAG-3′; PAD2 (173 bp) 5′-CATGTCCCAGATGATCCTGC-3′ and 5′-CATGGTAGAGCTTCCGCC-3′; PAD3 (314 bp) 5′-CTGTGAGGGAAGGCAGG-3′ and 5′-CATGCCTATAGGCCTCACAC-3′; PAD4 (377 bp) 5′-GGACTGCGAGGATGATG-3′ and 5′-GCTGTCTTGGAACACCAC-3′; α2-macroglobulin (147 bp), 5′-CAGCAGCAACCATGTCTTG-3′ and 5′-TGCAAACTCATCCGTCTCG-3′. Cloned cDNAs served as positive control.
Antibodies
Rabbit antibodies directed against chemically modified citrulline (anti-MC) were a kind gift from Dr T Senshu (Tokyo, Japan). Mouse monoclonal antibodies against vimentin (RV202) were a kind gift from Dr F Ramaekers (Maastricht, The Netherlands).
Isotype-specific antibodies directed against PAD types 2 or 4 were produced by immunisation of rabbits with PAD isotype-specific peptides (PAD2: aa 3–8 and aa 516–531; PAD4: aa 210–225 and aa 517–531) (Eurogentec, Seraing, Belgium).
To affinity purify the anti-PAD antisera, 1 mg of each of two peptides was conjugated to 8 mg bovine serum albumin in the presence of 0.05% glutaraldehyde. After one hour at room temperature the reaction was stopped by adding 0.1 volume of 1 M glycine.NaOH pH 7.2. The mixture was dialysed against coupling buffer (0.1 M NaHCO3 pH 8.3, 500 mM NaCl) and then coupled to 0.5 g CNBr activated Sepharose-4B (Amersham-Pharmacia) according to the manufacturer’s instructions. The column was equilibrated in washing buffer (PBS pH 7.4, 350 mM NaCl, 0.05% Nonidet P40 (NP-40)) before serum was added. Unbound antibodies were removed by extensive washing. Anti-PAD antibodies were eluted with buffer (100 mM glycine.HCl pH 2.5, 500 mM NaCl, 0.05% NP-40). Collected fractions were neutralised with 1 M Tris (~pH 11).
Western blot analysis
Cells were washed with PBS and subsequently scraped in ice cold 50 mM Tris.HCl pH 7.4, 100 mM KCl, 20 mM ethylene glycol bis(β-aminoethylether)-N,N′-tetraacetic acid (EGTA), 1 mM dithioerythritol, 1% NP-40, 0.5 mM phenylmethylsulphonyl fluoride and Complete protease inhibitor cocktail (Roche). Extracts were sonified three times for 15 seconds with a microtip sonifier (Branson) at 20% output. For detection of PAD enzymes, extracts were centrifuged at 12 000 g for 15 minutes. Soluble proteins were separated by sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE; 10%) and transferred to nitrocellulose membranes. For detection of citrullinated proteins, extracts were concentrated by acetone precipitation before separation by SDS-PAGE (13%) and transferred to Hybond-C extra (Amersham) membranes. Blots used to detect citrullinated proteins were chemically modified before immunostaining with anti-MC antibodies as described by Senshu et al.39 Blots were blocked in blocking buffer (PBS containing 5% non-fat dried milk and 0.1% NP-40) for one hour at room temperature and incubated for one to three hours with the antibody of interest diluted in blocking buffer. After washing with blocking buffer, bound antibodies were detected by incubation with horseradish peroxidase conjugated swine antirabbit IgG antibodies (DAKO), followed by chemiluminescence. Purified recombinant His-tagged human PAD2 or PAD4 (a gift from S Nijenhuis, Nijmegen, The Netherlands) or in vitro citrullinated human fibrinogen served as a positive control.
Immunoprecipitations of citrullinated vimentin
For each immunoprecipitation 85 μl mouse antivimentin (RV202) was coupled to 20 μl of a 50% suspension of protein G-agarose beads in IPP500 (500 mM NaCl, 10 mM Tris.Cl pH 8.0, 0.05% NP-40) for two hours at room temperature. Beads were washed three times with IPP500 and once with ice cold IPP150 (150 mM NaCl, 10 mM Tris.Cl pH 8.0, 0.05% NP-40). Extracts of ex vivo differentiated macrophages (prepared as described above) were added and the mixture was incubated for two hours at 4°C. After three washes with IPP150, bound proteins were eluted with SDS-sample buffer, separated by SDS-PAGE (13%), and transferred to Hybond-C extra (Amersham) membranes. The blots were stained with anti-MC antibodies to show the presence of citrullinated vimentin.
UTR reporter study
Human PAD2 cDNA (KIAA0994, AB023211, described by Kikuno et al40) was obtained from Kazusa DNA Research Institute, Japan. The 3′ untranslated terminal region (UTR) sequence was subcloned by PCR into the XbaI site of the pGL3-promotor vector (Promega). In six well culture plates, 0.4×106 COS-1 cells were seeded per well and transfected with 1 μg of both plasmids, pGL3-promotor (either with or without PAD2 3′ UTR sequence) plasmid DNA and pG5luc reporter vector (Promega), using 5 μl Fugene transfection reagent (Roche), as described by the manufacturer. After 40–48 hours of growth, cells were harvested using 500 μl Passive Lysis Buffer (PLB; Promega) and the activity of both the firefly luciferase and the control renilla luciferase was determined by the Dual Luciferase Reporter Assay System (Promega) on a Berthold Lumat LB 9507 luminometer. In brief, 100 μl of firefly luciferase substrate solution was added to 20 μl of cell extract and the luminescence was measured to determine the efficiency of the interaction. Next, 100 μl of renilla luciferase substrate solution (containing a quencher for the firefly luciferase activity) was added and again the luminescence was determined to monitor the transfection efficiency. Results are expressed as the ratio of firefly versus renilla luciferase activity (mean (SD)).
RESULTS
PAD type 2 and type 4 mRNAs are expressed in CD14 positive cells
Mononuclear leucocytes are important in the pathology of RA. We investigated the expression of four isotypes of PAD in mononuclear blood cells. Total PBMCs were isolated from a healthy person by Ficoll density gradient centrifugation. Subsequently cells were subdivided with antibody coated magnetic beads into T cell (CD3+), B cell (CD19+), monocyte (CD14+), and NK cell (CD56+) fractions. Total RNA was isolated from the cells and each isotype of PAD was amplified by RT-PCR using PAD isotype-specific primers. Human PAD1 is mainly expressed in epidermis and uterus24,25,27; PAD3 expression is restricted to hair follicles.26 It is therefore not surprising that PAD1 and PAD3 were undetectable in the PBMCs (fig 1). Human PAD2 is expressed in a broad range of tissues, including skeletal muscle, uterus, brain, salivary glands, and pancreas.23 Human PAD4 is found in haematopoietic cells.23 PAD2 and PAD4 mRNA could both be detected in the PBMCs. The vast majority of the expression was found in the CD3–, CD19–, CD56– and CD14+ fractions. PAD2 and PAD4 mRNA are thus predominantly expressed by CD14 positive cells (fig 1, indicated by white squares).

Messenger RNA expression of PAD isotypes in PBMCs. Total PBMCs from a healthy person were fractionated with MACS magnetic beads coated with either anti-CD3 (T cells), anti-CD14 (monocytes), anti-CD19 (B cells) or anti-CD56 (NK cells) for RNA isolation. Expression of PAD was analysed by RT-PCR. PAD1 and 3 were not detectable in any of the PBMC fractions. For PAD2 and PAD4, specific PCR products were produced in all fractions. In the case of CD3, CD19, and CD56, the cells that did not bind to the beads (CD3– CD19–, CD56–) showed higher expression than the cells that did bind (CD3+, CD19+, CD56+). In contrast, the cells that were bound by the anti-CD14 coated beads (CD14+, monocytes) showed higher PAD2 and PAD4 expression than the cells that did not bind the anti-CD14 coated beads (CD14–, all but the monocytes) as indicated by white squares. β-Actin served as a control for mRNA input. Cloned cDNAs served as positive PCR controls, (no positive control included for β-actin), PCR without template as negative control.
Differential expression of PAD during monocyte to macrophage differentiation
To investigate the PAD2 and PAD4 expression by CD14+ cells in more detail, peripheral blood monocytes were differentiated into macrophages. Total PBMCs were isolated from the peripheral blood of patients with RA and healthy controls by Ficoll density gradient centrifugation. Monocytes were isolated by adherence to plastic six well culture plates. After removal of non-adherent cells, the monocytes were differentiated into macrophages in seven days. PAD mRNA expression was evaluated by RT-PCR. Human PAD2 mRNA could be detected in equal amounts in monocytes and in macrophages (fig 2). No significant up or down regulation was observed during monocyte differentiation. PAD4 mRNA, however, could only be seen in the monocytes. The PAD4 mRNA expression was lost during differentiation (fig 2). Cells from patients with RA and healthy controls showed identical expression patterns.
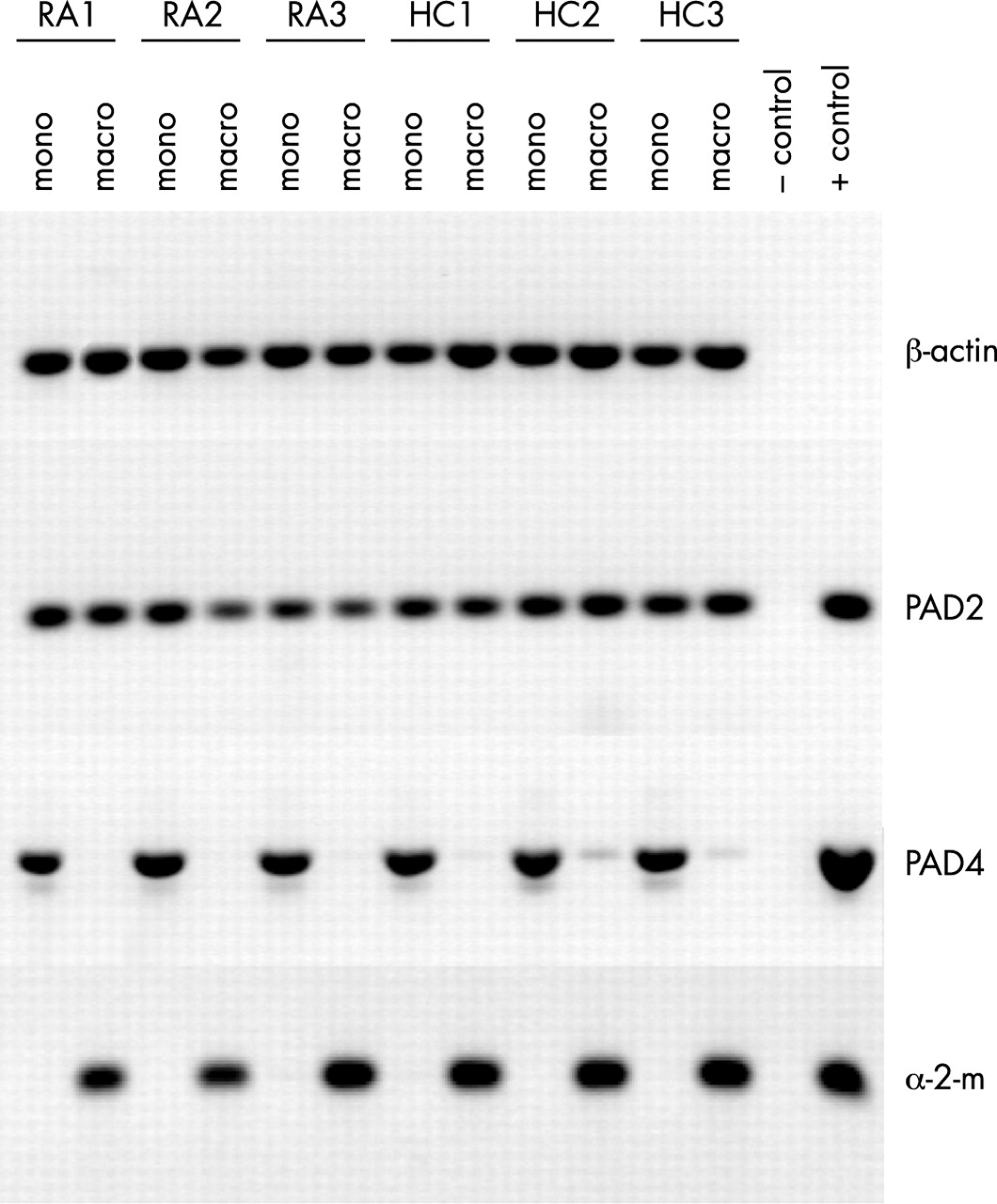
PAD4 mRNA is only expressed in monocytes, not in macrophages. RNA expression of PAD2 and PAD4 in freshly isolated monocytes (mono) from peripheral blood and in ex vivo differentiated macrophages (macro) of patients with RA (RA1–RA3) and healthy controls (HC1–HC3) was analysed by RT-PCR. β-Actin served as a control for mRNA input, α2-macroglobulin (α-2-m) expression was analysed as a control for macrophage differentiation. Cloned cDNAs served as positive PCR controls, (no positive control included for β-actin), PCR without template as negative control.
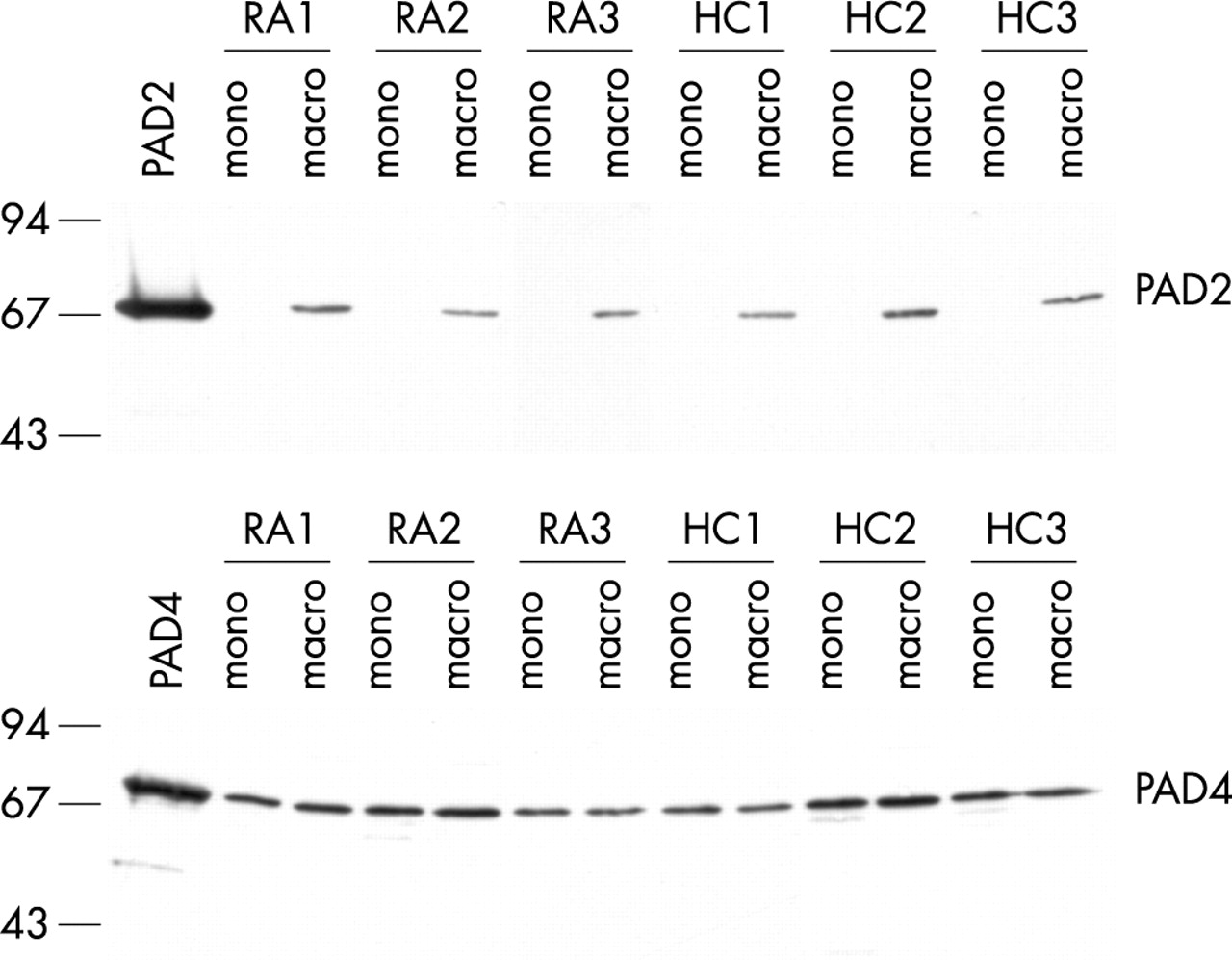
The same cells were also tested for PAD protein expression. Protein extracts were analysed by immunoblotting using PAD isotype-specific antibodies. Surprisingly, PAD2 protein was only present in macrophages. Although PAD2 mRNA was equally expressed in monocytes and macrophages, very little (if any) PAD2 protein could be detected in the monocytes (fig 3). These results indicate that the differential expression of PAD2 is regulated at the translational level. PAD4 protein was detected at similar levels in monocytes and macrophages (fig 3). It appears that, although the message for PAD4 is gradually lost during the differentiation, the PAD4 protein level remains stable during the seven days of differentiation. Again, cells from patients with RA and healthy controls showed identical patterns of expression.

PAD2 protein is only expressed in macrophages. Protein expression of PAD2 and PAD4 in freshly isolated monocytes (mono) from peripheral blood and in ex vivo differentiated macrophages (macro) of patients with RA (RA1–RA3) and healthy controls (HC1–HC3) was analysed by western blotting. Recombinant PAD2 or PAD4 proteins are included as controls. Molecular weight markers (kDa) are indicated on the left.
Differential expression of PAD during monocyte infiltration into the joint
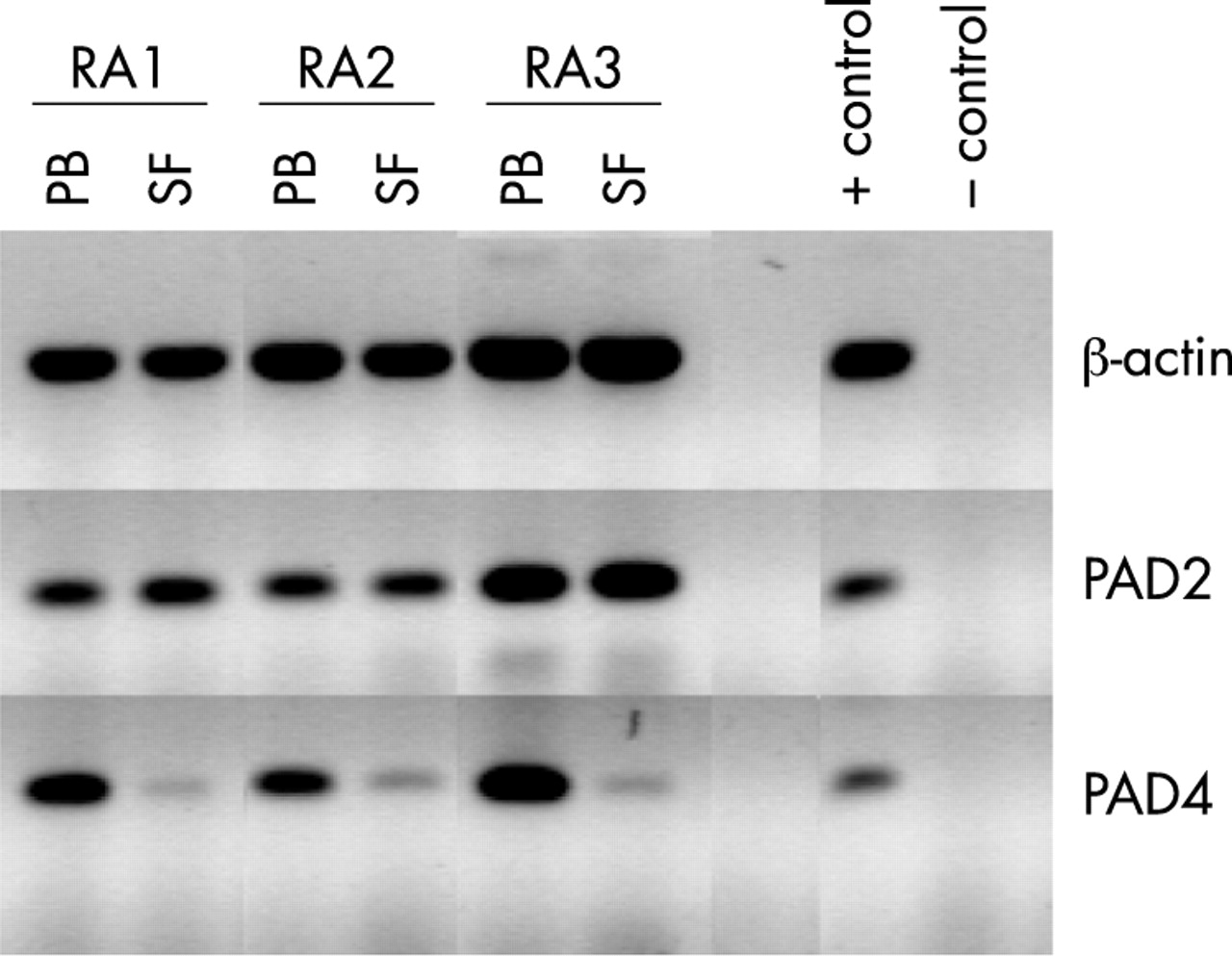
The macrophages used in the previous experiments were differentiated ex vivo from circulating monocytes. Monocytes that infiltrate the synovium during inflammation will also differentiate into macrophages, albeit in an inflammatory environment. The differences between ex vivo and in vivo environments could induce subtle differences in the resulting phenotype of the macrophages. We therefore compared PAD expression in PMBCs and SFMCs of patients with RA. Mononuclear cells were isolated by Ficoll density gradient centrifugation from paired PB and SF samples. Total RNA was isolated for RT-PCR analysis, protein extracts were prepared for immunoblotting analysis.
PAD2 mRNA could be detected in equal amounts in PBMCs and SFMCs (fig 4), whereas PAD2 protein could only be detected in significant amounts in the SFMCs (fig 5). These patterns of expression are thus identical to the patterns observed in ex vivo differentiated cells (figs 2 and 3). PAD2 mRNA expression remains unaltered during infiltration of circulating monocytes into the synovium and subsequent differentiation into macrophages. Because PAD2 protein is only expressed in the cells from the SF it appears that translation of its messenger RNA is initiated during macrophage differentiation.

PAD4 mRNA is only present in PBMCs, not in SFMCs. Mononuclear cells were isolated from PB and SF samples of patients with RA. RNA expression of PAD2 and PAD4 was analysed by RT-PCR. β-Actin served as a control for mRNA input. Cloned cDNAs served as positive PCR controls, (no positive control included for β-actin), PCR without template as negative control.

PAD2 protein is only expressed in SFMCs. Protein expression of PAD2 and PAD4 in mononuclear cells isolated from PB and SF samples of patients with RA was analysed by western blotting. Recombinant PAD2 and PAD4 proteins are included as controls. Molecular weight markers (kDa) are indicated on the left.
The situation for PAD4 is also identical to the observations made with the ex vivo differentiated cells (figs 2 and 3). PAD4 mRNA could only be detected in the PBMCs (fig 4). PAD4 protein could be detected in similar amounts in the PBMC and SFMC fractions (fig 5). The PAD4 mRNA is thus gradually lost during synovial infiltration and subsequent macrophage differentiation; notwithstanding that the PAD4 protein levels remain unchanged. These results show again that the PAD4 protein that is already present in the PB monocytes is not degraded during monocyte infiltration and differentiation in the synovium.
PAD2 translation is regulated by its 3′ UTR
Although monocytes contained comparable amounts of PAD2 mRNA as macrophages or SFMCs, they contained little, if any, PAD2 protein. It thus appears that the PAD2 mRNA is not translated in monocytes and that translation is initiated somewhere during macrophage differentiation. PAD2 is encoded by a 4.5 kb transcript (AB023211),41,42 which contains a very long 3′ UTR (>2 kb). To investigate if this 3′ UTR might be involved in regulation of PAD2 expression at the translational level, we cloned the PAD2 3′ UTR behind a luciferase reporter gene. Constructs containing firefly luciferase with or without PAD2 3′ UTR were transfected into COS-1 cells. A plasmid containing renilla luciferase was cotransfected as a transfection control.
Mean luciferase activity (ratio of firefly to renilla luciferase activity) without PAD2 3′ UTR was set to 100% (±10.3). Addition of the PAD2 3′ UTR caused a sixfold reduction of luciferase activity (16.3±2.0%). This and the observed expression patterns, indicate that the 3′ UTR sequence is important in regulating PAD2 protein expression at the translational level.
Citrullinated proteins are only generated after calcium influx
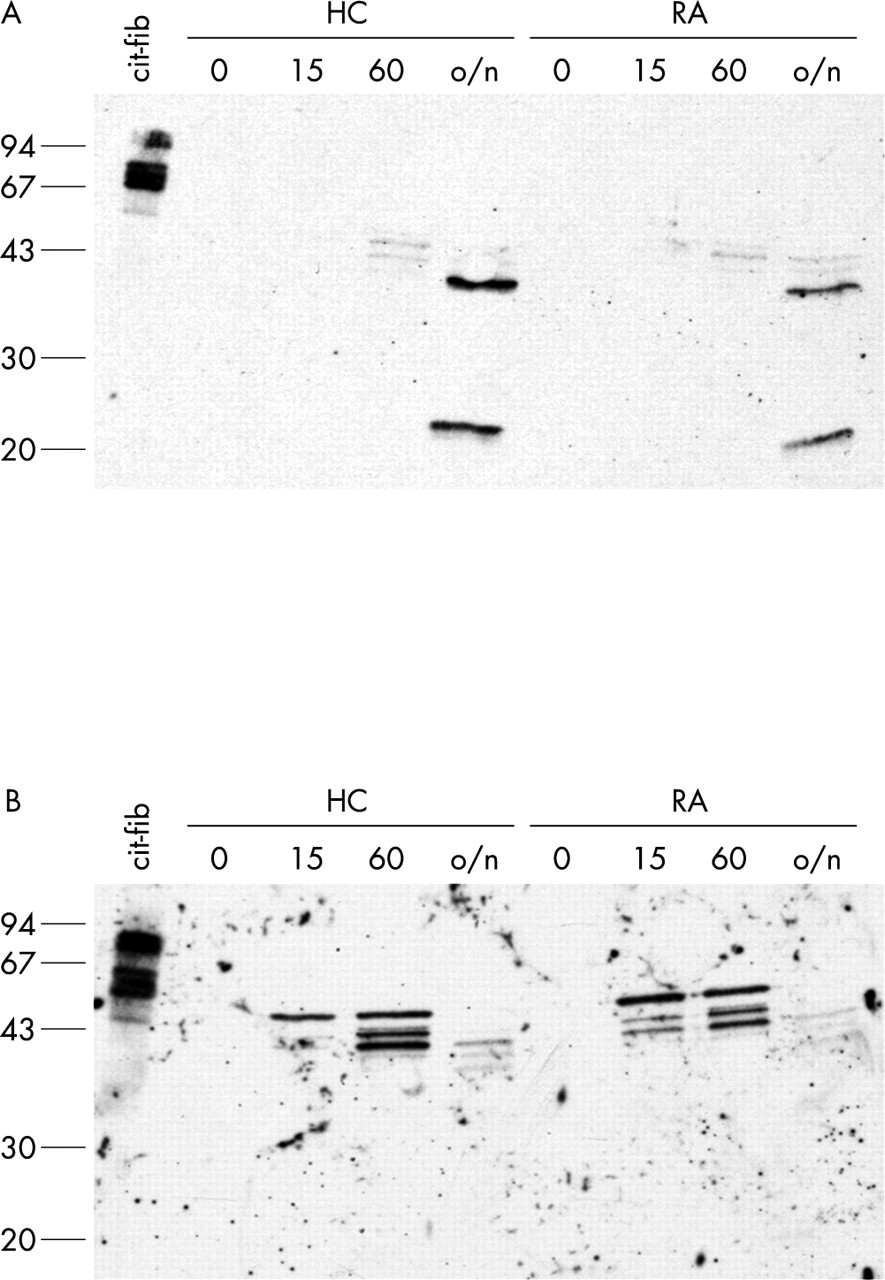
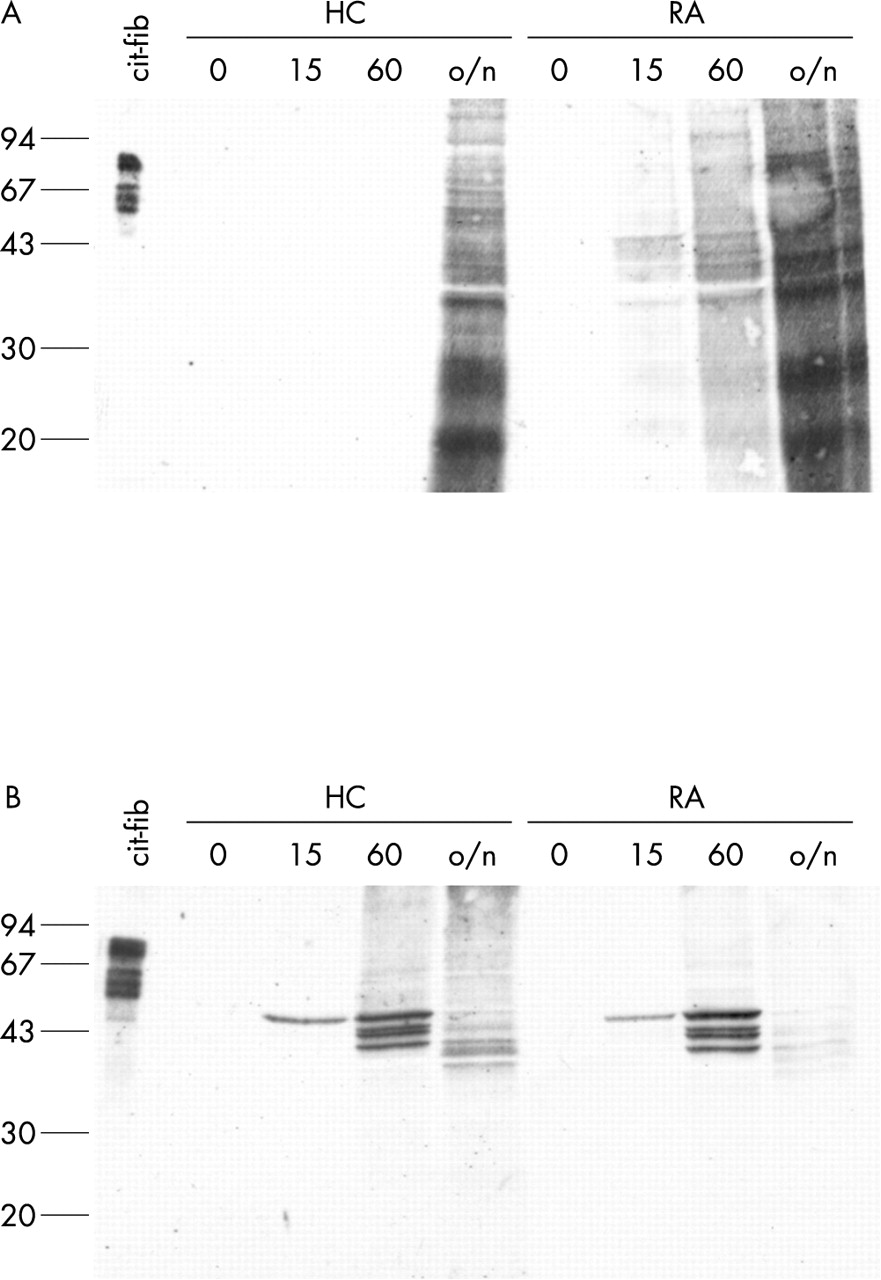
For detection of citrullinated proteins in protein extracts or fixed cells, Senshu and coworkers developed a method in which the citrulline side chain is specifically modified into an artificial amino acid side chain that is so bulky that the influence of flanking amino acids for epitope recognition becomes negligible.39,43,44 We used these anti-MC antibodies to detect citrullinated proteins in freshly isolated monocytes and ex vivo differentiated macrophages. Figure 6 (lanes indicated with 0) shows that no citrullinated proteins could be detected in either the monocytes or the macrophages.

Citrullinated proteins are only generated after calcium influx. Monocytes (A) and macrophages (B) from a healthy control (HC) and a patient with RA were treated for 0, 15 minutes, 60 minutes, or overnight (o/n) with 1 μM ionomycin in the presence of 2 mM CaCl2. Extracts were analysed for the presence of citrullinated proteins by western blotting with anti-MC antibodies. In vitro citrullinated human fibrinogen (25 ng) is included as a positive control. Molecular weight markers (kDa) are indicated on the left.
It thus appears that the mere presence of PAD enzymes in both monocytes (PAD4) and macrophages (PAD2 and PAD4) is not sufficient for the citrullination of intracellular proteins. It is known, however, that calcium ions, and possibly other factors, are required for activity of the PAD enzymes. The threshold Ca2+ concentration for PAD activity is about 10−5 mol/l (Nijenhuis S, Nijmegen, The Netherlands, personal communication)45; the cytosolic calcium concentration in normal cells is much lower (10−7 mol/l). The cytosolic calcium concentration can be artificially raised by addition of various agents like ionomycin or thapsigargin. Ionomycin is a calcium ionophore which facilitates the sustained influx of extracellular calcium. Thapsigargin is an inhibitor of the ubiquitous sarco-endoplasmic reticulum Ca2+-ATPases causing release of calcium from the intracellular Ca2+ stores into the cytosol.46 Monocytes and ex vivo differentiated macrophages were therefore treated with 1 μM ionomycin or 1 μM thapsigargin in the presence of 2 mM extracellular calcium for different periods of time (15 minutes, 60 minutes, or overnight).
As expected, proteins are citrullinated upon ionomycin treatment (fig 6). In macrophages (fig 6B), several distinct bands of citrullinated proteins could be detected. After overnight (~18 hours) treatment with ionomycin, the intensity of the staining decreased and the bands appeared to shift to a lower molecular weight, indicating that the citrullinated proteins are degraded. In monocytes (fig 6A), proteins are citrullinated more slowly than in macrophages (fig 6B). Also different proteins appear to be citrullinated. No differences were seen between cells from patients with RA and healthy controls. Citrullination of intracellular proteins was also observed after treatment with thapsigargin (data not shown).
Citrullinated proteins can be immunoprecipitated by antivimentin antibodies
The four bands that are detectable in the macrophages after 60 minutes of ionomycin treatment are in the molecular weight range of the intermediate filament protein vimentin and the observed banding pattern resembles the banding pattern observed with antivimentin (RV202) monoclonal antibodies (data not shown). Specific citrullination of vimentin during calcium ionophore-induced apoptosis in mouse peritoneal macrophages has been reported previously.35 To investigate if indeed vimentin is the citrullinated protein observed after ionomycin treatment, we immunoprecipitated vimentin from ionomycin treated cell extracts. Immunoblots containing immunoprecipitated vimentin were subsequently stained with anti-MC antibodies (fig 7). No citrullinated vimentin could be detected in untreated cells (lanes labelled 0). In monocytes (fig 7A), citrullinated vimentin or its degradation products were detectable after overnight treatment with ionomycin. In contrast, citrullinated vimentin was already detectable in macrophages after 15 minutes of ionomycin treatment (fig 7B). In the macrophages that were treated overnight, only degradation products of citrullinated vimentin could be detected. Analysis of cells from patients with RA and healthy controls gave identical results.
Vimentin is specifically citrullinated in calcium stimulated macrophages. Immunoprecipitated vimentin from monocytes (A) and macrophages (B) from a healthy control (HC) and a patient with RA, treated for 0, 15 minutes, 60 minutes, or overnight (o/n) with 1 μM ionomycin in the presence of 2 mM CaCl2, was stained with anti-MC antibodies. In vitro citrullinated human fibrinogen (25 ng) is included on the western blot as a positive control. Molecular weight markers (kDa) are indicated on the left.
DISCUSSION
We investigated the expression of four PAD isotypes in PBMCs: PAD1 to PAD4. The human PAD4 was previously known as PAD5 because it showed slightly different reaction kinetics towards artificial substrates than the rodent PAD4 enzymes.31 Sequence data, expression data, intracellular localisation, and the genomic organisation of the PAD genes,23 however, all indicate that human PAD5 corresponds to the rodent PAD4.32,33 Our suggestion to rename the human PAD5 to PAD4 was recently approved by the HUGO Gene Nomenclature Committee.23
The most noticeable difference between the different PAD isotypes is their tissue-specific expression (reviewed by Vossenaar et al23). PAD1 and PAD3 are expressed mainly in epidermis and hair follicles.23–27 It is therefore not surprising that their respective mRNAs were undetectable in PBMCs. Expression of PAD2 and PAD4 has been reported in a variety of tissues, including haematopoietic cells.23,28–33 In the PMBCs, both PAD2 and PAD4 mRNA were predominantly expressed in the monocytes (CD14+, CD3–, CD19–, CD56– cells).
Differentiation of the monocytes into macrophages (both ex vivo and in vivo) has several consequences for PAD expression (summarised in fig 8). The mRNA expression of PAD4 is lost during differentiation. Nevertheless, the PAD4 protein levels remain unchanged, indicating that the enzyme is stable and not degraded. In contrast, PAD2 mRNA levels remain unchanged, while PAD2 protein levels significantly increase during differentiation. It appears that the translation of PAD2 mRNA is blocked in the monocytes. This apparent discrepancy has previously been described in epidermis; although PAD2 mRNA was found in the epidermis,47 PAD2 protein was absent.48

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Model for synovial PAD infiltration. Many leucocytes infiltrate the inflamed synovium in RA. When monocytes from the PB enter the synovium, they will differentiate into macrophages. This differentiation process has several consequences for PAD expression: the mRNA expression of PAD4 is lost during differentiation, but the PAD4 protein levels remain unchanged, indicating that the enzyme is not rapidly degraded. For PAD2, the situation is different: the mRNA levels remain more or less the same, but the mRNA is only translated into PAD2 protein in the macrophages. The patterns of PAD expression are identical in patients with RA and healthy controls, but because of the large numbers of macrophages that can be found in the RA (and not in healthy) synovium, the amount of PAD enzyme in the RA synovium will be much higher than in healthy synovium. During apoptosis or necrosis of the PAD-containing macrophages, free intracellular Ca2+ levels are increased, resulting in activation of the PAD enzymes and citrullination of cellular proteins. In the macrophages, vimentin is specifically citrullinated after increase of cytoplasmic Ca2+. Furthermore, PAD enzymes may leak out of necrotic cells, causing the citrullination of extracellular proteins (for example, fibrin21).
Expression of many proteins is regulated post-transcriptionally by 5′ and 3′ UTR motifs. The 5′ UTR of PAD2 is only 64 nts in length, in contrast, the 3′ UTR measures 2.3 kb. PAD4, in contrast, contains a much shorter 3′ UTR of 245 nts. By cloning the very long 3′ UTR of PAD2 behind a luciferase reporter gene, we showed that the 3′ UTR is responsible for inhibition of translation, as luciferase activity was reduced over sixfold by the PAD2 3′ UTR. Most, if not all, mRNAs regulated by 3′ UTR motifs are subjected to regulation during early embryonic development or during differentiation (reviewed by Standart and Jackson49). Examples of such differentially regulated mRNAs are protamines in developing sperm cells50 and 15-lipoxygenase during erythroid differentiation.51,52 A general mechanism for post-transcriptional regulation by 3′ motifs is the binding of regulatory proteins to specific differentiation control elements in the 3′ UTR region of the mRNA.49 These regulatory proteins interact with the 5′ end of the mRNA by forming a loop structure. In this way they prevent the binding of initiation factors to the mRNA.53 Which parts of the 3′ UTR of PAD2 and which regulatory proteins are involved in the observed cell stage-specific inhibition of translation, is currently under investigation.
Although PAD enzymes were present in monocytes (PAD4) and macrophages (PAD2 and PAD4), they apparently were not active, because no citrullinated proteins were detectable. The cytosolic and nucleoplasmic Ca2+ concentration under normal physiological conditions is much too low (about 100-fold) for PAD activity.45 Raising the cytosolic and nucleoplasmic calcium concentration by treatment with either ionomycin or thapsigargin induced activation of PAD and subsequent citrullination of intracellular proteins. As shown in fig 6, monocytes and macrophages had different banding patterns of citrullinated proteins. PAD activation by treatment with a calcium ionophore has been described previously for mouse macrophages,35 rat keratinocytes,36 and human HL-60 granulocytes.33,54 A rise in cytosolic Ca2+ levels is important for induction of apoptosis.55,56 During apoptosis (or necrosis) the plasma membrane Ca2+ pump (PMCA) is cleaved by caspases, so that influxing calcium can no longer be efficiently cleared by PMCA, which results in Ca2+ overload.57 This may explain why citrullination in vivo takes place during death or terminal differentiation of cells (for example, citrullination of filaggrin and keratin during terminal differentiation of keratinocytes43,58,59), when control of calcium homoeostasis is lost.
Vimentin is specifically citrullinated in the macrophages after Ca2+ influx. This observation has previously been reported for mouse peritoneal macrophages,35 although no suggestions were made at that time about which PAD isotype is responsible for vimentin citrullination. We show here that macrophages contain both PAD2 and PAD4, the former is located in the cytosol, the latter in the nucleus.33 Vimentin is an intermediate filament protein and thus is located in the cytosol. This makes PAD2 the most likely candidate for vimentin citrullination. The high cytosolic Ca2+ levels in the ionomycin treated monocytes and macrophages induce cell death,35,55 which leads to nuclear fragmentation and subsequent release of PAD4 into the cytosol. This may explain why vimentin is citrullinated much more slowly in monocytes, which only contain PAD4 protein (fig 7).
After overnight exposure to high cytosolic Ca2+, citrullinated vimentin was shown to be degraded. In healthy cells, the vimentin cytoskeleton is a highly dynamic structure, as polymerisation and depolymerisation of the filaments are involved in various biological functions. Under physiological conditions, these processes are regulated by reversible phosphorylation at specific sites in the amino terminal head domain of the vimentin molecules.60 In apoptotic cells, the network of vimentin filaments is disrupted; it collapses into perinuclear aggregates. Two processes may contribute to this collapse. Citrullination of the amino terminal head domain by PAD induces disassembly of the vimentin filaments.61 Furthermore, vimentin is subjected to cleavage by caspases early in apoptosis.62–64 Although no data exist on a possible correlation between these two processes in this specific case, in general, citrullination of proteins causes protein unfolding, which makes them more susceptible to proteolysis.23,65–67
Figure 8 shows a schematic representation of the events that may lead to the production of citrullinated vimentin in the inflamed synovium. Monocytes are recruited to the inflamed synovium by various chemoattractant factors. After infiltration of the synovium, monocytes will differentiate into macrophages, which causes up regulation of PAD2 protein and down regulation of PAD4 mRNA expression.
The macrophages that are abundantly present in the RA synovial tissue (in RA 80–100% of synovial lining cells are macrophage-like cells, compared with 20–30% in normal synovium34) show signs of activation.68 Short term activated macrophages are protected from apoptosis, whereas long term (>24 hours) activated macrophages are very prone to cell death.69 Calcium influx in the dying macrophages triggers activation of PAD and subsequent citrullination of vimentin. Furthermore, PAD enzymes may leak out of dead cells, enabling them to citrullinate extracellular proteins, like fibrin.21,70 The likely presence of citrullinated vimentin in the rheumatoid synovium, makes it a potential autoantigen in RA. Interestingly, antibodies directed to the Sa antigen (which might be citrullinated vimentin71) are highly specific for RA and can be detected in about 50% of patients with RA.72
In conclusion, our data show that citrullination of proteins by PAD enzymes is a process regulated at three levels: transcription, translation, and activation (by Ca2+ influx). Vimentin is specifically citrullinated by macrophages after Ca2+ influx and citrullinated vimentin is a candidate autoantigen in RA. PAD2 and PAD4 are the most likely candidate PAD isotypes for the citrullination of synovial proteins in RA.
Acknowledgments
We thank Dr Tatsuo Senshu (Tokyo, Japan) for providing antibodies directed against chemically modified peptidylcitrulline, Suzanne Nijenhuis (Nijmegen, The Netherlands) for providing purified recombinant human PAD2 and PAD4 protein, Drs Ger Pruijn and Reinout Raijmakers (Nijmegen, The Netherlands) for useful discussion and assistance, and Dr Rob Moots (Liverpool, UK) and his staff for help in optimising the RT-PCR procedures. This work would have been impossible without the generous blood donations of all participating patients with RA and healthy subjects.
This work was financially supported by the Netherlands Foundation for Chemical Research and the Netherlands Technology Foundation (grant 349–5077), Het Nationaal Reumafonds of The Netherlands (the Dutch League against Rheumatism, grant 00-2-402), and the Netherlands Foundation for Medical Research (NWO grant 940-35-037).