Article Text

Abstract
Background: Leflunomide and its active metabolite A77 1726 reversibly inhibits the enzyme dihydro-orotate dehydrogenase, the rate limiting step in de novo synthesis of pyrimidines and progression of the cell cycle in different cell lines, mainly activated T lymphocytes.
Objective: To analyse in vitro the possible anti-inflammatory effects exerted by A77 1726, on cultured macrophages, obtained from the synovial tissues of patients with rheumatoid arthritis (RA).
Methods: The effects of different doses of A77 1726 on intracytoplasmic expression and extracellular concentration of inflammatory cytokines (tumour necrosis factor α (TNFα), interleukin (IL) 1β, IL6), as well as the influence on production and expression of intercellular adhesion molecule-1 (ICAM-1) and cyclo-oxygenase 2 (COX-2) by primary cultures of synovial macrophages from patients with RA, were evaluated by immunocytochemistry and western blot analysis. The observations were made at four and 24 hours.
Results: A progressive and significant time and dose dependent decrease of the number of positive macrophages for intracellular TNFα and IL1β, treated with different doses of A77 1726, was found in comparison with untreated cells. The extracellular concentration of TNFα was found to be significantly decreased in media containing cultured macrophages at 24 hours for all tested doses of A77 1726. At 24 hours, a significant time and dose dependent decrease of ICAM-1 and COX-2 expression by cultured macrophages after A77 1726 treatment was found.
Conclusions: In conclusion, the mechanism of antiproliferative activity exerted by leflunomide on activated T lymphocytes seems to be the same mechanism (alteration of the cell cycle progression) which interferes with the functions of other activated cells—namely, the monocytes/macrophages, which are strongly involved in the inflammatory reaction in RA synovial tissue. The positive clinical results seem to confirm that leflunomide exerts an anti-inflammatory action on phagocytic cells in short and long term treatment of RA.
- leflunomide
- rheumatoid arthritis
- macrophages
- inflammation
- cytokines
- COX-2, cyclo-oxygenase 2
- DRMRI, dynamic gadolinium enhanced magnetic resonance imaging
- ICAM-1, intercellular adhesion molecule-1
- IL, interleukin
- MTT, methyl thiazolyl tetrazolium
- PBS, phosphate buffered saline
- PBS/T, phosphate buffered saline/Tween 20
- RA, rheumatoid arthritis
- TNFα, tumour necrosis factor α
Statistics from Altmetric.com
- COX-2, cyclo-oxygenase 2
- DRMRI, dynamic gadolinium enhanced magnetic resonance imaging
- ICAM-1, intercellular adhesion molecule-1
- IL, interleukin
- MTT, methyl thiazolyl tetrazolium
- PBS, phosphate buffered saline
- PBS/T, phosphate buffered saline/Tween 20
- RA, rheumatoid arthritis
- TNFα, tumour necrosis factor α
Synovial inflammation and subsequent degradation of the joints in rheumatoid arthritis (RA) are initiated by the homing of activated lymphocytes and monocytes into the synovial tissue.1,2
Definition of the role and activities of different lymphocyte subsets, as well as of monocytes/macrophages at the site of the synovial inflammatory reaction, might lead to the development of effective and targeted treatments for RA.3 In particular, unlike other cells, activated lymphocytes in RA require roughly an eightfold increase in their levels of rUMP (and other pyrimidine ribonucleotides) in order to progress from G1 to the S phase of the cell cycle and they must use de novo synthesis of pyrimidines.4,5
Identification of drugs that regulate cell progression through the cell cycle in an attempt to modulate proliferation shows that leflunomide (N-(4-trifluoro-methylphenyl)-5-methylisoxazole-4-carboxamide) is an antirheumatic drug with a unique ability to regulate such progression.6,7
Leflunomide and its active metabolite A77 1726 at low, therapeutically applicable doses reversibly inhibits the enzyme dihydro-orotate dehydrogenase,8,9 the rate limiting step in de novo synthesis of pyrimidines in different cell lines.10 At higher doses leflunomide inhibits expression of tyrosine kinases and induces down regulation of cell signalling in the G0/G1 phase of the cell cycle.11,12
However, recent studies suggest that the observed anti-inflammatory effects exerted by A77 1726, the active metabolite of leflunomide, may be related to its ability to suppress tumour necrosis factor α (TNFα) and interleukin (IL) 1β production during the cell-cell contact activation between T lymphocyte and monocytes.13,14
A77 1726 has also been shown to suppress the activation of nuclear factor κB (NF-κB), a potent mediator of inflammation when stimulated by inflammatory agents.10 In addition, continuing research indicates that A77 1726 seems to down regulate the glycosylation of adhesion molecules such as intercellular adhesion molecule-1 (ICAM-1), inducing a further reduction of the cell-cell contact activation and homing of inflammatory cells during the inflammatory reaction.15–17 Recently, a significantly reduced number of macrophages, a significantly decreased expression of ICAM-1 and vascular cell adhesion molecule-1, and a reduction of TNFα and IL1β was detected in synovial tissue samples obtained from patients with RA after four months of treatment with leflunomide.18
Because monocytes and macrophages are considered to be the major source of inflammatory mediators in RA synovial tissue, including the inducible cyclo-oxygenase 2 (COX-2), this study aimed at analysing in vitro the possible anti-inflammatory effects exerted by A77 1726 on cultured macrophages obtained from the synovial tissue of patients with RA. Therefore, the effects of A77 1726 on intracytoplasmic expression and extracellular concentration of inflammatory cytokines (TNFα, IL1β, IL6), as well as on the production and expression of ICAM-1 and COX-2 by cultured synovial RA macrophages as a consequence of TNF signalling on NF-κB, were evaluated.19,20
MATERIALS AND METHODS
Cell cultures
Primary cultures of synovial macrophages were obtained from five patients with RA classified according to the American College of Rheumatology criteria (four women, mean (SD) age 52 (3) years) with a mean (SD) duration of disease of 3 (1) years.21
Synovial samples were obtained during arthroscopic synovectomy of the knee. At the time of surgery, patients had been treated exclusively with non-steroidal anti-inflammatory drugs for at least three months.
The synovial RA tissue was cut into small pieces, washed in Dulbecco phosphate buffered saline (PBS; Sigma-Aldrich, Milan, Italy), and incubated in RPMI-1640 (Sigma-Aldrich, Milan, Italy) supplemented with collagenase (0.75 mg/ml) for one hour at 37°C. The digest was passed through a wire 150 mesh to separate dissociated cells from the tissue debris. The synovial macrophages were washed three times with Dulbecco PBS, resuspended in RPMI-1640 medium supplemented with 10% fetal bovine serum (5% CO2 humidified atmosphere at 37°C), and treated with lipopolysaccharide 10 μg/ml. The cells were cultured in the absence or presence of different concentrations (1, 10, 30 μmol/l) of A77 1726 (HMR1726, provided by Aventis Pharma Deutschland GmbH, Frankfurt am Main Germany) for four or 24 hours. These concentrations were close to the therapeutic dose.22
At the end of the incubation time, the cells were harvested, washed in PBS, and used for different test assays. Experiments were done in triplicate.
Cell survival evaluation
A cell survival test was performed to establish the possible cytotoxicity of A77 1726 on synovial macrophages. Cell survival was evaluated by the methyl thiazolyl tetrazolium (MTT) salt test, a quantitative colorimetric assay used to detect cell survival and proliferation and based on the ability of living cells to cleave the tetrazolium ring in active mitochondria.
Cells were seeded into 96 well microtitre plates and were treated according to the experimental design. At the established time, 50 μl of MTT labelling reagent (3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide 5 mg/ml in PBS); Sigma-Aldrich, Milan, Italy) was added to each well and incubated in a humidified atmosphere at 37°C. Four hours later, 100 μl of dimethyl sulphoxide was added to each well and mixed thoroughly. The absorbance was calculated at 540 nm with a scanning multiwell spectrophotometer. The number of cells was obtained from this optical density reading using a standard reference curve. Our data indicate that A77 1726 caused no cellular damage in any of the cultures (data not shown).
Cytokine immunoassay
Macrophages were incubated with different concentrations (1, 10, 30 μmol/l) of A77 1726. After four and 24 hours the culture medium was harvested and stored at –20°C until analysis. The enzyme immunoassay for quantitative determination of TNFα (sensitivity <0.115 pg/ml) and IL1β (sensitivity 2 pg/ml) was carried out with a microplate kit system (Biosource Europe SA, Nivelles, Belgium). The results were obtained with a multiplate automatic processor (Thecno Genetics, Milan, Italy).
Intracytoplasmic cytokine staining
Macrophages were transferred into 15 ml centrifuge tubes and washed twice, then a permeabilisation buffer with saponin (Sigma-Aldrich, Milan, Italy) was added at concentration of 0.5% for 10 minutes at room temperature. The cells were centrifuged and resuspended in a permeabilisation buffer together with cytokine-specific antibodies (anti-TNFα, anti-IL1β, anti-IL6; Santa Cruz Biotecnhology, Santa Cruz, CA, USA) at a dilution of 1:100 for 30 minutes at room temperature. The cells were then treated with FITC conjugated secondary antibody (Santa Cruz Biotechnology) at a dilution of 1:28 for 30 minutes at room temperature and analysed by fluorescent microscopy.
Western blot analysis
After treatment with different concentrations (1, 10, 30 μmol/l) of A77 1726, macrophage pellets were lysed in buffer containing 20 mM Tris-HCl, 150 mM NaCl, 1 mM phenylmethylsulphonylfluoride, 5 mg/ml aprotinin, and 0.5% Nonidet (Promega) P-40 for one hour at 4°C. The lysates were then centrifuged for 10 minutes at 13 000 rpm. Samples were diluted with reducing sample buffer and separated by 10% sodium dodecyl sulphate-polyacrylamide gel electrophoresis. The proteins were transferred to a Hybond-C-nitrocellulose membrane (Amersham Pharmacia Biotech Europe, Freiburg Germany). After transfer, the membrane was blocked overnight at 4°C in PBS containing 5% non-fat powdered milk.
For immunoblot analysis, the blot was incubated with COX-2 antibody (Santa Cruz, Biotechnology) at a dilution of 1:500 and ICAM-1 antibody (Santa Cruz Biotechnology) at a dilution of 1:500 for one hour at room temperature, shaken, washed in 0.05% PBS/Tween 20 pH 7.4 (PBS/T) and, finally, incubated in secondary horseradish peroxidase labelled goat antirabbit antibody (Santa Cruz Biotechnology) at a dilution of 1:5000 for one hour at room temperature. After three further washes with PBS/T, bound secondary antibody was detected by emitting chemiluminescent light (Amersham Pharmacia Biotech Europe, Freiburg Germany).
Immunocytochemistry assay
Macrophages harvested after different treatments were incubated on polylysinated glass slides for 40 minutes at 4°C. Then, the spots were air dried and fixed in methanol/acetone (1:1 vol/vol) for 30 seconds and stored at −20°C until the immunodetermination of COX-2 and ICAM-1. After rehydration in PBS, spots were incubated with rabbit anti-COX-2 and ICAM-1 human polyclonal antibody at room temperature (Santa Cruz Biotechnology).
Immobilised antibodies were detected by antirabbit immunoglobulin serum and alkaline phosphatase anti-alkaline phosphatase complex (Vector Laboratory Inc, Burlingame, CA, USA). Each step was followed by two washes in PBS. Alkaline phosphatase was developed with a mixture of naphthol AS-BI phosphate and New Fuchsin (Merck, Milan, Italy). Levamisole was added to the development solution. Finally, slides were counterstained with haematoxylin and cover slipped with Eukitt. Controls were treated identically, but the primary or secondary antibodies were omitted.
Image analysis
Image analysis was performed with the Leica Q500 MC image analysis system (Leica, Cambridge, UK). A measure of the positive area was given by the number of pixels/μm2 detected in the field falling within the frame. For each sample about 100 cells were analysed. For the digital image analysis, several areas (at least 10) were evaluated on the labelled slides, and the mean values are reported.
Statistical analysis
Data were obtained from experiments carried out in triplicate and are expressed as means (SEM). Statistical analysis was performed by an analysis of variance Tukey-Kramer test, and a value of p<0.05 was considered significant.
RESULTS
Effects of A77 1726 on cytokine production by cultured synovial macrophages
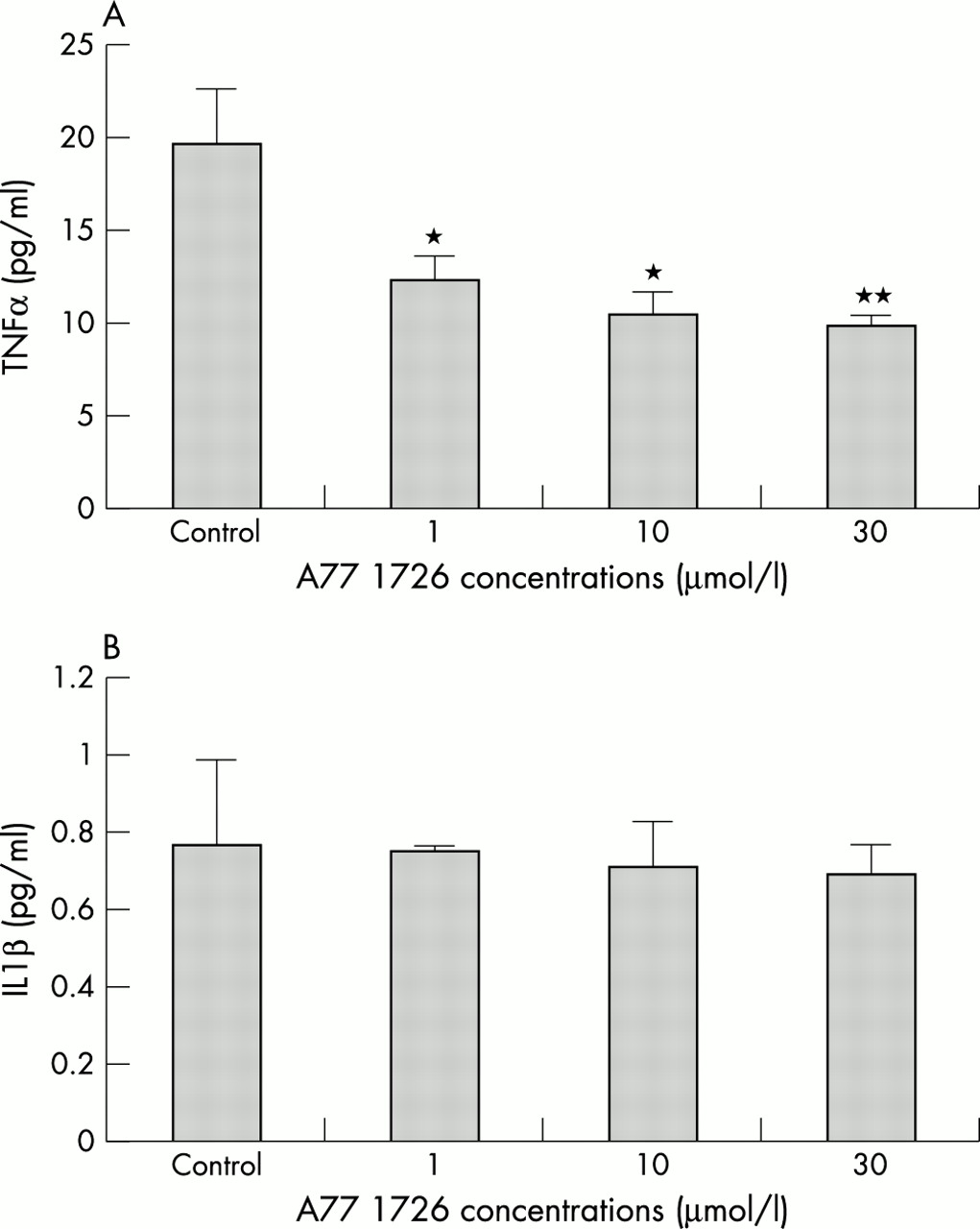
A significant decrease (between 40 and 50%) in TNFα levels was seen at 24 hours in the cultured macrophages treated with A77 1726 at all concentrations compared with untreated controls (p<0.05 for cells treated with 1 and 10 μmol/l, p<0.01 for cells treated with 30 μmol/l versus controls, respectively) (fig 1A). In contrast, no significant differences in IL1β levels were found under the same conditions at 24 hours after treatment with A77 1726 (fig 1B).

(A) TNFα concentrations at 24 hours in culture media after incubation of RA synovial macrophages with A77 1726 at concentrations of 1, 10, and 30 μmol/l. *p<0.05; **p<0.01 versus controls. (B) IL1β concentrations at 24 hours in culture media after incubation of RA synovial macrophages with A77 1726 at concentrations of 1, 10, and 30 μmol/l. No significant changes were found.
Interestingly, basal production of TNFα was found to be significantly higher than the basal production of IL1β (figs 1A and B).
Effects of A77 1726 treatment on intracytoplasmic cytokine production
The optimal kinetics of cytokine production by cultured macrophages under different conditions were determined by a method of staining for intracellular cytokines originally described by Andersson et al.23 The number of untreated cells positive for TNFα and IL1β was found to be higher at four hours (2% and 40%, respectively) than at 24 hours (2% and 10%, respectively), whereas the number of untreated cells, positive for IL6 was found to be similar at four and 24 hours (10%).
A progressive and significant time and dose dependent decrease in the number of TNFα and IL1β positive RA synovial macrophages treated with A77 1726 was seen in comparison with untreated cells (figs 2A and 2B, 5E and F).

(A) Intracytoplasmic expression of TNFα, IL1β and IL6 in RA synovial macrophages treated with different concentrations of A77 1726 after four hours. The percentage of TNFα and IL1β positive cells decreased significantly in a dose dependent manner. *p<0.05; **p<0.01; ***p<0.001 versus controls. IL6 was poorly expressed and the effects of A77 1726 were not significant. (B) Intracytoplasmic expression of TNFα, IL1β, and IL6 in RA synovial macrophage cells with different concentrations of A77 1726 after 24 hours. The percentage of IL1β and IL6 positive cells decreased significantly in a dose dependent manner. *p<0.05; ***p<0.01 versus controls. TNFα was poorly expressed at 24 hours and the effect of A77 1726 was not significant.
In particular, TNFα positive macrophages decreased significantly with A77 1726 at four hours (1–10 μmol/l and 30 μmol/l, p<0.05 and p<0.01, respectively) and further at 24 hours (10 μmol/l and 30 μmol/l, p<0.001).
Similarly, IL1β positive macrophages decreased significantly with A77 1726 at four hours (1, 10, and 30 μmol/l, p<0.05, p<0.01, and p<0.001, respectively) and further at 24 hours (1 μmol/l and 10–30 μmol/l, p<0.05, p<0.001, respectively).
IL6 positive macrophages were poorly expressed and the effects of A77 1726 were not significant at any time and dose.
Effects of A77 1726 on ICAM-1 expression
The expression of ICAM-1 is a marker of macrophage activation, which has a crucial role in cell/cell signalling. Two methods were used to detect the ICAM-1 expression: immunocytochemistry staining and western blot protein analysis.
At 24 hours a significant and dose dependent decrease of ICAM-1 expression after A77 1726 treatment with 10 and 30 μmol/l was found (p<0.01 and p<0.001, respectively) (figs 3A, 5C and D). These results were confirmed by the western blot analysis (fig 3B).

(A) ICAM-1 expression in RA synovial macrophages treated with different concentrations of A77 1726 after 24 hours as evaluated by immunocytochemistry and image analysis. The staining positivity was expressed as a percentage for cell number. **p<0.01; ***p<0.001. (B) ICAM-1 expression in RA synovial macrophages treated with different concentrations of A77 1726 after 24 hours as evaluated by western blot analysis.
Effects of A77 1726 on COX-2 production
Again two methods were used to detect the ICAM-1 expression: immunocytochemistry staining and western blot protein analysis.
At 24 hours a significant decrease of the intracytoplasmic staining for COX-2 was only seen in macrophages treated with A77 1726 30 μmol/l (p<0.001) (figs 4A, 5A and B). Western blot protein analysis for COX-2 confirmed these results (fig 4B). However, a progressive dose related decrease of COX-2 expression was detectable in the western blot analysis, supporting a dose dependent inhibition by A77 1726.

(A) COX-2 expression in RA synovial macrophages treated with different concentrations of A77 1726 after 24 hours as evaluated by immunocytochemistry and image analysis. The staining positivity was expressed as a percentage for cell number. ***p<0.001. (B) COX-2 expression in RA synovial macrophages treated with different concentrations of A77 1726 after 24 hours as evaluated by western blot analysis.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
COX-2 expression by cultured RA synovial macrophages with (A) and without (B) treatment with A77 1726 (30 μmol/l). ICAM-1 expression by cultured RA synovial macrophages with (C) and without (D) treatment with A77 1726 (30 μmol/l). Intracytoplasmic IL1 expression by cultured RA synovial macrophages with (E) and without (F) treatment with A77 1726 (30 μmol/l).
DISCUSSION
This study suggests that leflunomide (through its active metabolite A77 1726) may have anti-inflammatory effects, which are dose dependent, on activated human cultured macrophages from patients with RA. The observed anti-inflammatory activity seems to be related to the inhibition of cytokine production (TNFα and IL1β after four hours of A77 1726 treatment, IL6 after 24 hours), and also to the down regulation of ICAM-1 and COX-2 expression.
Previous studies, on cell-cell contact activation between T lymphocytes and monocytes, have shown up regulation of proinflammatory cytokines such as TNFα and IL1β and their inhibitors.24 These cytokines play a central part in the inflammatory process in RA synovial tissue and are mainly produced by activated monocytes/macrophages.20 Because leflunomide mainly inhibits, at the level of activated T lymphocytes, the enzyme dihydro-orotate dehydrogenase, which is the rate limiting step in de novo synthesis of pyrimidines, the cell cycle of other activated inflammatory cells might also be altered by leflunomide.8,25,26
Therefore, the observed reduced synthesis of proinflammatory cytokines produced by the action of A77 1726 on activated macrophages might be related to the induced reversible inhibition of de novo pyrimidine synthesis, sufficient to alter the progression of their cell cycle. These changes, both at the intracytoplasmic level and in cultured macrophages, were already evident at four hours, confirmed at 24 hours, and clearly dose dependent.
Concerning ICAM-1 expression, A77 1726 by depleting the pyrimidine pool seems to down regulate the glycosylation of adhesion molecules, further reducing cell-cell contact activation and homing of inflammatory cells during the inflammatory reaction.15,16
This study seems to suggest that reduced expression of ICAM-1 (30–55%) on cultured macrophages, mainly evident at 24 hours (A77 1726, 30 μmol/l), may well be related to the possibly altered de novo synthesis of these adhesion molecules. Therefore, a further anti-inflammatory activity exerted by leflunomide in RA seems to be the reduction of possible cell-cell contact activation by decreasing the intercellular adhesion molecule expression on synovial macrophages.
Finally, by taking into account the fact that synovial RA macrophages are indicated as the major producers of inducible COX-2 and of the inflammatory prostaglandins, leflunomide was shown, through its active metabolite A77 1726 at the highest concentration (30 μmol/l), to reduce the expression of COX-2 fourfold in comparison with untreated RA macrophages.27,28
COX-2, one of the major mediators of the inflammatory reaction in RA synovial tissue, is also strongly induced in activated monocytes/macrophages. Again, it is suggested that the reversible inhibition of de novo pyrimidine synthesis caused by leflunomide, which is sufficient to alter progression of the cell cycle in activated cells (that is, macrophages), might interfere also with COX-2 induction and contribute to the anti-inflammatory activity.29 However, direct effects have also been suggested.27
All these in vitro results seem to confirm the recent observation of a significantly reduced number of macrophages, together with decreased expression of TNFα, IL1β, and ICAM-1, in synovial tissue samples obtained from patients with RA after four months’ treatment with leflunomide.18
Recently, dynamic gadolinium enhanced magnetic resonance imaging (DRMRI) analysis of synovial tissue in patients with RA treated with leflunomide showed a significant reduction of the inflammatory reaction.30 Because the DRMRI correlates with the infiltrating leucocytes in the RA synovial tissue, it was confirmed that leflunomide exerts anti-inflammatory activity on monocytes/macrophages.18,30
Interestingly, very similar results for the effects on basic mechanisms of inflammation were seen with methotrexate, another antiproliferative drug exerting anti-inflammatory effects in RA.18,31–34
In conclusion, the mechanism of antiproliferative activity exerted by leflunomide on activated T lymphocytes seems to be the same mechanism (alteration of the cell cycle progression) which interferes with the functions of other activated cells—namely the monocytes/macrophages, which are strongly involved in the inflammatory reaction in RA synovial tissue.
The broad anti-inflammatory action exerted by leflunomide may indicate its possible use in short and long term treatment of RA.