Article Text

Abstract
Objective: To investigate whether interleukin 17 (IL17), derived specifically from T cells, can promote type II collagen release from cartilage. The ability of IL17 to synergise with other proinflammatory mediators to induce collagen release from cartilage, and what effect anti-inflammatory agents had on this process, was also assessed.
Methods: IL17 alone, or in combination with IL1, IL6, oncostatin M (OSM), or tumour necrosis factor α (TNFα), was added to bovine nasal cartilage explant cultures. Proteoglycan and collagen release were determined. Collagenolytic activity was determined by bioassay. Chondroprotective effects of IL4, IL13, transforming growth factor β1 (TGFβ1) and insulin-like growth factor-1 (IGF1) were assessed by inclusion in the explant cultures.
Results: IL17 alone stimulated a dose dependent release of proteoglycan and type II collagen from bovine nasal cartilage explants. Suboptimal doses of IL17 synergised potently with TNFα, IL1, OSM, and IL6 to promote collagen degradation. This collagen release was completely inhibited by tissue inhibitor of metalloproteinase-1 and BB-94 (a synthetic metalloproteinase inhibitor), and was significantly reduced by IL4, IL13, TGFβ1, and IGF1. In IL17 treated chondrocytes, mRNA expression for matrix metalloproteinase (MMP)-1, MMP-3, and MMP-13 was detected. Moreover, a synergistic induction of these MMPs was seen when IL17 was combined with other proinflammatory cytokines.
Conclusions: IL17 can, alone and synergistically in combination with other proinflammatory cytokines, promote chondrocyte mediated MMP dependent type II collagen release from cartilage. Because levels of all these proinflammatory cytokines are raised in rheumatoid synovial fluids, this study suggests that IL17 may act as a potent upstream mediator of cartilage collagen breakdown in inflammatory joint diseases.
- cartilage
- collagen
- interleukin 17
- synergy
- CNTF, ciliary neurotrophic factor
- CT, cardiotrophin
- GAGs, glycosaminoglycans
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- gp, glycoprotein
- IGF, insulin-like growth factor
- IL, interleukin
- LIF, leukaemia inhibitory factor
- LDH, lactate dehydrogenase
- MMP, matrix metalloproteinase
- OHPro, hydroxyproline
- OSM, oncostatin M
- RA, rheumatoid arthritis
- TGF, transforming growth factor
- TIMPs, tissue inhibitors of metalloproteinases
- TNF, tumour necrosis factor
Statistics from Altmetric.com
- CNTF, ciliary neurotrophic factor
- CT, cardiotrophin
- GAGs, glycosaminoglycans
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- gp, glycoprotein
- IGF, insulin-like growth factor
- IL, interleukin
- LIF, leukaemia inhibitory factor
- LDH, lactate dehydrogenase
- MMP, matrix metalloproteinase
- OHPro, hydroxyproline
- OSM, oncostatin M
- RA, rheumatoid arthritis
- TGF, transforming growth factor
- TIMPs, tissue inhibitors of metalloproteinases
- TNF, tumour necrosis factor
Rheumatoid arthritis (RA) is characterised by chronic inflammation of the synovium and destruction of articular cartilage. Synovial inflammation involves the infiltration and accumulation of activated macrophages and T cells, and proliferation of synovial fibroblasts. The most prominent cytokines in RA synovial fluid, such as interleukin-1 (IL1), tumour necrosis factor α (TNFα), oncostatin M (OSM), and IL6 are derived from macrophages and synovial fibroblasts.1–3 In contrast, levels of T cell derived cytokines are very low or absent.4
A novel human T cell derived cytokine, IL17, has been described that is produced by CD4+ activated memory (CD45+, RO+) T cells,5 a dominant subset of T cells present in RA synovial tissues6). Raised levels of IL17 have been reported in RA synovium7 and synovial fluids.8 IL17 is a pleiotropic cytokine that can stimulate proinflammatory cytokine production from a variety of cell types present within the joint, including macrophages, synovial fibroblasts, and chondrocytes.5,9–11 Taken together, these data suggest a potential role for IL17 in the initiation and perpetuation of inflammation in RA and other inflammatory arthritides.
Articular cartilage is a highly specialised tissue that allows smooth joint articulation. Cartilage is synthesised and maintained by chondrocytes, and is composed primarily of water, proteoglycan, and type II collagen. Entrapped within the type II collagen fibrillar network are proteoglycan molecules that pull water into the tissue. A rapid release of proteoglycan can be initiated from cartilage by stimulation with cytokines such as IL1 and TNFα, but this matrix component can be replaced relatively quickly.12 In contrast, collagen is much less readily released, but when collagen breakdown does occur, the structural integrity of the tissue is irreversibly lost.13 Degradation of type II collagen, therefore, represents a key control point in cartilage turnover.
A family of closely related enzymes, the matrix metalloproteinases (MMPs), collectively can degrade all the components of the extracellular matrix. Of these, the mammalian collagenases (MMP-1, MMP-8, and MMP-13) specifically cleave triple helical collagen to give characteristic three quarter and one quarter fragments,14 which can then undergo further proteolysis. Collagenases are known to be present within the rheumatoid joint and can be localised to rheumatoid tissues.15,16 MMPs are synthesised as zymogens that are activated by proteolytic removal of the propeptide region, and these active enzymes are inhibited by the tissue inhibitors of metalloproteinases (TIMPs).17 Regulation of these enzymes is critical to the turnover of the extracellular matrix.
Recent reports have indicated a role for IL17 in the turnover of connective tissues in RA. IL17 can inhibit the synthesis, as well as stimulate the breakdown, of type I collagen in RA synovium and bone explants.18 Moreover, the spontaneous release of type I collagen fragments from RA synovium and bone ex vivo was at least partially mediated by endogenous IL17.18 Animal studies have also suggested a role for IL17 in several models of arthritis.19,20 For cartilage, there are no reports to date on the effects of IL17 on type II collagen release, although IL17 has been shown to inhibit proteoglycan synthesis and stimulate proteoglycan loss in murine articular cartilage.21,22
This study seeks to study further the role of IL17 in cartilage destruction. Because collagen loss represents the irreversible step in cartilage destruction,13 we investigated the ability of IL17 to promote type II collagen release from cartilage and the role of MMPs in this process. MMP inhibitors and a number of anti-inflammatory agents were also assessed for their effect on IL17-induced cartilage collagen degradation. Moreover, we investigated the ability of IL17 to synergise with several other proinflammatory cytokines known to be present in the rheumatoid joint, including TNFα, IL1, OSM, and IL6.1–3
MATERIALS AND METHODS
Culture medium and test reagents
Cytokines, growth factors, and the soluble IL6 receptor (sIL6R) were all recombinant human. IL6, IL11, IL17, ciliary neurotrophic factor (CNTF), TNFα, transforming growth factor β1 (TGFβ1), insulin-like growth factor 1 (IGF1) and sIL6R were purchased from R&D Systems (Abingdon, UK). OSM and leukaemia inhibitory factor (LIF) were kindly provided by Professor J Heath (University of Birmingham, Edgbaston, UK). Cardiotrophin-1 (CT-1) was a generous gift from Dr D Pennica (Genentech, San Francisco, CA). IL1α and IL4 were produced in-house. All cytokines were stored at −70°C and diluted into culture medium immediately before use. Recombinant human TIMP-1 was produced in-house.23 BB-94, a broad spectrum metalloproteinase inhibitor, was generously provided by British Biotech Pharmaceuticals Ltd (Oxford, UK). Molecular biology reagents were purchased from Amersham Pharmacia Biotech (Little Chalfont, UK), and all other chemicals and biochemicals were commercially available analytical grade reagents as previously described.3,24 Control culture medium was Dulbecco's modification of Eagle's medium (Gibco, Paisley, UK) containing 25 mM HEPES supplemented with glutamine (2 mM), penicillin (100 IU/ml), streptomycin (100 μg/ml), and nystatin (20 IU/ml).
Cartilage degradation assay
Cartilage degradation assays were performed as previously described using bovine nasal septum cartilage in explant culture.3,24 Briefly, discs (2 mm in diameter) were cut from 2 mm slices of bovine nasal septum cartilage and washed twice in phosphate buffered saline (Sigma, Poole, UK) containing penicillin (100 IU/ml), streptomycin (100 μg/ml), and nystatin (20 IU/ml). Three discs per well of a 24 well plate were incubated in 600 μl of control culture medium for 24 hours. Fresh control medium (600 μl), with or without test reagents (four wells for each condition), was then added and the plate incubated at 37°C for seven days. Culture supernates were harvested and replenished with identical treatments as on day 1, and the experiment continued until day 14. Day 7 and 14 supernates were stored at −20°C until assay. The remaining cartilage was digested with papain (4.5 mg/ml) in 0.1 M phosphate buffer, pH 6.5, containing 5 mM EDTA and 5 mM cysteine hydrochloride, for 16 hours at 65°C. Digests were stored at −20°C until assay.
Viability of cartilage explants was assessed by screening for the production of lactate dehydrogenase (LDH) using the Cytotox 96 assay (Promega, Southampton, UK). This is always performed when new cytokine combinations are used with tissue explants. No increase in LDH levels with any of the cytokine combinations has been found (data not shown). Serum is excluded from cartilage explants as it can markedly alter the metabolism of cartilage in the absence of exogenous cytokine(s). Moreover, because this cartilage model is one of cartilage breakdown, we avoided using chondroprotective agents such as IGF125,26 which are known to be present in serum and able to replace it (see Rowan et al24 and references listed therein). The absence of serum has also been shown not to affect the viability of the tissue (data not shown), and previous studies have shown that cartilage in serum-free culture for 8–9 days can respond to serum and other growth factors.27
Proteoglycan, collagen, and collagenolytic activity assays
Culture supernates and cartilage digests were assayed for sulphated glycosaminoglycans (GAGs) (as a measure of proteoglycan) and hydroxyproline (OHPro) (as a measure of collagen) using modifications of the 1,9-dimethylmethylene blue dye binding28 and OHPro29 assays, respectively, as previously described.30 The release of proteoglycan and collagen was expressed as a percentage of the total. Collagenolytic activities present in the supernates from cartilage explants were determined by a 96 well plate modification of the diffuse fibril assay using 3H-acetylated collagen.31 Procollagenases were artificially activated by co-incubation with 0.7 mM 4-aminophenylmercuric acetate in order to obtain total (pro- + active) collagenolytic activity. One unit of collagenase activity degrades 1 μg of collagen per minute at 37°C. One unit of inhibitory activity inhibits two units of collagenolytic activity by 50%.
Cell culture, RNA isolation, and northern blot analysis
Primary bovine nasal chondrocytes were extracted from cartilage as previously described.32 Primary chondrocytes were cultured to confluence in control medium supplemented with 10% fetal calf serum. Cells were then serum-starved overnight before stimulation with test reagents in serum-free control medium for 24 hours as described previously.30 Total cellular RNA was isolated using the RNeasy kit (Qiagen, Crawley, UK) under RNAse free conditions according to the manufacturer's instructions. Equal amounts (20 μg/lane) of total RNA were fractionated on 1% agarose-0.4 M formaldehyde gels and stained with ethidium bromide to check for integrity. RNA was transferred to GeneScreen Plus membrane (NEN, Hounslow, UK) by capillary transfer. Blots were then sequentially hybridised with [α32P]dCTP-labelled cDNA probes and washed as previously described.30 Probe-specific mRNA was detected by exposure to Phosphor screen (Molecular Dynamics, Chesham, UK) and subsequent visualisation with a STORM 860 Phosphorimager (Molecular Dynamics). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used to assess equal loading of RNA.
Statistical analysis
The data were analysed with GraphPad Prism v2.01 (GraphPad Software Inc, San Diego, USA). Pairwise multiple comparisons were performed using the Bonferroni multiple comparison test. Values of ph0.05 were considered significant.
RESULTS
Effect of IL17 on the release of proteoglycan and collagen from cartilage explants
IL17 (0.4–50 ng/ml) induced a dose dependent breakdown of bovine nasal cartilage (fig 1). By day 7 of culture a significant and reproducible release of proteoglycan was observed with IL17 at concentrations between 2 and 50 ng/ml (fig 1A). Significant collagen release was induced by IL17 in a dose dependent manner by day 14 (fig 1B). The levels of collagen release at 50 ng/ml IL17 showed marked variation (ranging from 6 to 95% release) in six separate experiments using different cartilage samples, similar to that seen with IL1 alone.3,24–26,30,32 However, statistically significant collagen release was seen in almost all experiments compared with control (5/6 experiments; p≤0.05). When IL17 alone did not induce significant collagen release, it always promoted marked proteoglycan release (ph0.001, n=6; see below) and always synergised to promote significant collagen release (see below). IL17 alone also induced a significant increase in the collagenolytic activity detected in day 14 culture supernates (fig 1C). Although no collagenolytic activity was detected in the supernates of cartilage stimulated with 2 ng/ml IL17, a significant amount of procollagenase activity was present. The IL17-induced collagenolytic activity was completely inhibited by the exogenous addition of TIMP-1 (70 U/ml) into the bioassay (fig 1D).

Effect of IL17 on the release of proteoglycan and collagen, and the production of collagenolytic activity, from bovine nasal cartilage in explant culture. Three discs of cartilage per well in quadruplicate were cultured for 14 days in control medium ± IL17 (0.4–50 ng/ml). Supernates were harvested at day 7, replenished with identical treatments and harvested on day 14. The levels of (A) GAGs (as a measure of proteoglycan), and (B) OHPro (as a measure of collagen) released into the medium by day 7 and day 14, respectively, were determined as described in the “Materials and methods”. Results were expressed as a percentage of the total (mean (SD)). (C) Active and total collagenolytic activities (open and closed bars, respectively) present in day 14 media were determined by bioassay. (D) TIMP-1 (70 U/ml) was included in some bioassays of day 14 media to assess its effect on the collagenolytic activity present. Bonferroni's multiple comparison test was used to compare IL17 with control (A–D), and the same test was used to compare IL17 with control or IL17 + TIMP-1 in the bioassay (D), where ***p≤0.001; **p≤0.01
Effect of metalloproteinase inhibitors on IL17-induced collagen release
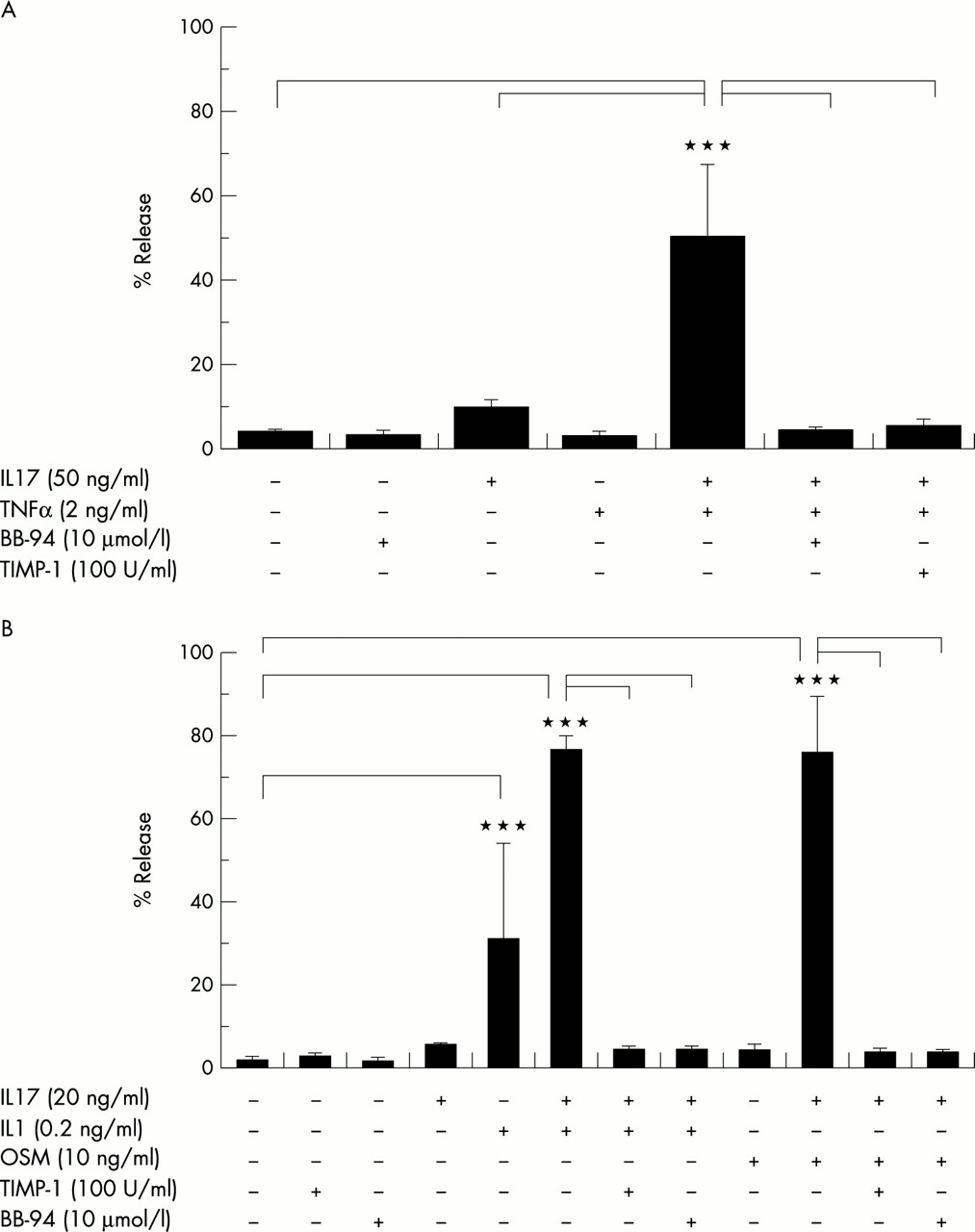
Bovine nasal cartilage explants were cultured with IL17 (100 ng/ml) in the presence or absence of TIMP-1 (100 U/ml) and the hydroxamate based metalloproteinase inhibitor BB-94 (10 μmol/l) for 14 days. IL17-induced collagen release from explant cultures was completely inhibited by co-incubation with these inhibitors (fig 2).

Effect of metalloproteinase inhibitors on IL17-induced collagen release from cartilage. Bovine nasal cartilage explants were cultured as described in fig 1 except that IL17 was at 100 ng/ml and either TIMP-1 (100 U/ml) or BB-94 (10 μmol/l) were also included. Day 14 culture supernates were assayed for OHPro as a measure of collagen, and the results expressed as a percentage release of the total (mean (SD)). Bonferroni's multiple comparison test was used to compare IL17 alone with control or IL17 + inhibitor, where ***p≤0.001.
Effects of anti-inflammatory agents on IL17-induced cartilage collagen release
Bovine nasal cartilage explants were incubated for 14 days with IL17 (50 ng/ml) in combination with IL4 (50 ng/ml), IL13 (50 ng/ml), TGFβ1 (20 ng/ml), or IGF1 (200 ng/ml). IL17-induced collagen release was inhibited by all these anti-inflammatory agents (p≤0.01); owing to the variable collagen release seen with IL17 alone, at least three separate cartilage samples were assessed (table 1).
Effect of anti-inflammatory agents on IL17-induced collagen release from cartilage
Effect of IL17 in combination with TNFα, IL1, or OSM on cartilage breakdown
To determine the effect of proinflammatory cytokines on IL17-induced cartilage breakdown, bovine nasal cartilage was incubated for 14 days with IL17 (0.01–50 ng/ml) ± TNFα (2 and 10 ng/ml), IL1α (0.1 ng/ml), or OSM (10 ng/ml). TNFα, IL1α, and OSM all synergised markedly with IL17 at suboptimal doses (that is, ≤10-fold less is required in combination than when alone) to induce proteoglycan release (fig 3). With IL17 alone, modest but significant collagen release was observed at doses between 10 and 50 ng/ml (fig 4). However, when IL17 at the suboptimal doses of 0.05 ng/ml and 0.5 ng/ml was combined with TNFα (10 and 2 ng/ml, respectively), a synergistic collagen release was seen (fig 4A). This synergy was even more marked with increasing concentrations of IL17. Similarly, IL17 at 0.08 ng/ml synergised with IL1α to induce significant collagen loss (fig 4B), and though less marked, a synergistic release of collagen was also observed with IL17 (10 ng/ml) in combination with OSM (fig 4B).
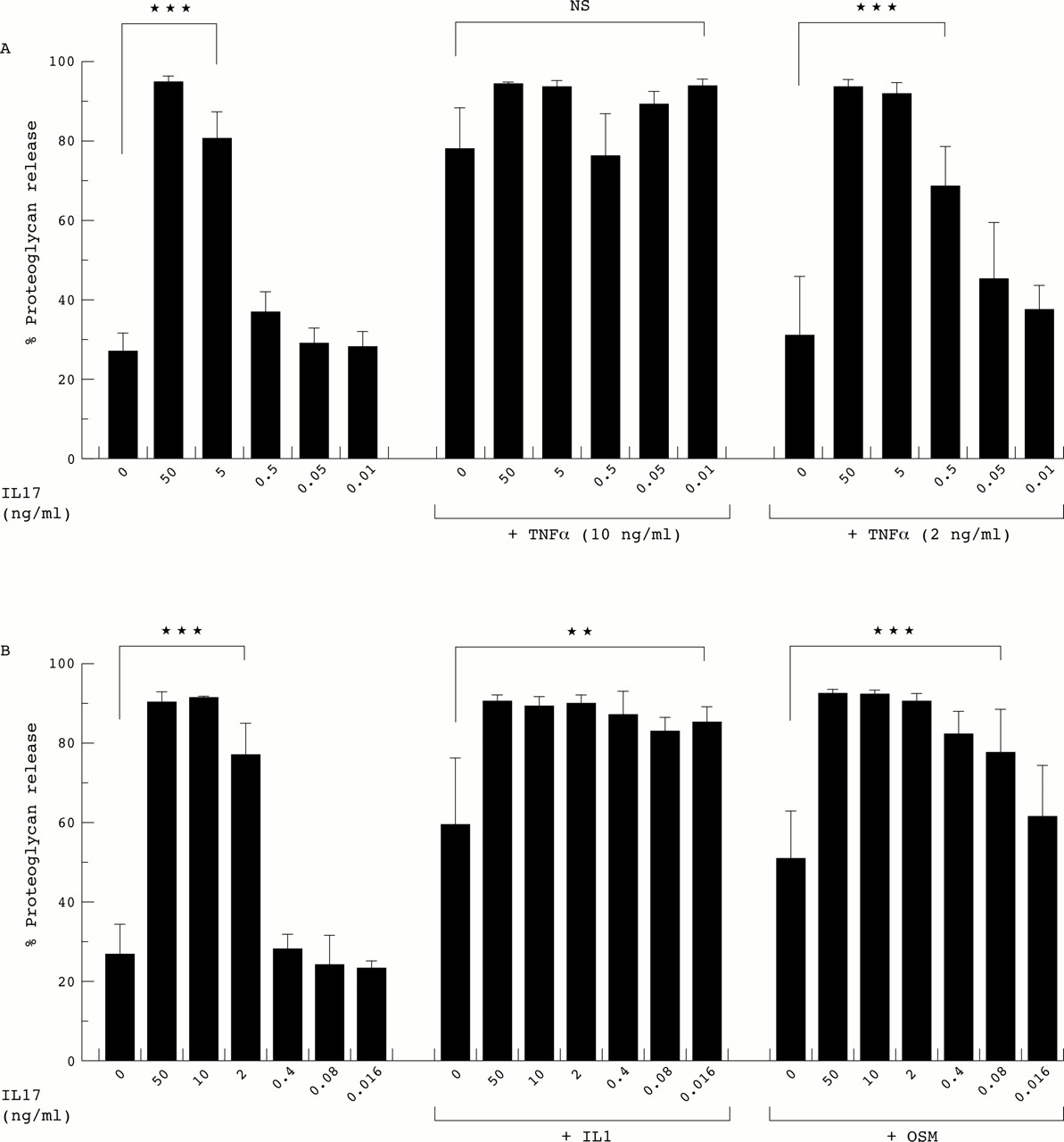
Effect of IL17 in combination with TNFα, IL1, or OSM on proteoglycan release from cartilage. Bovine nasal cartilage explants were cultured as described in fig 1 with IL17 (0.01–50 ng/ml) ± (A) TNFα (2 and 10 ng/ml), (B) IL1α (0.1 ng/ml) or OSM (10 ng/ml). Culture supernates (day 7) were assayed for the release of proteoglycan as described in “Materials and methods”, and the results expressed as a percentage release of the total (mean (SD)). Bonferroni's multiple comparison test was used to compare IL17 alone with control, and IL17 + TNFα or IL1 or OSM with TNFα or IL1 or OSM alone, where ***p≤0.001; **p≤0.01; NS, not significant. For the selected comparisons shown, the statistical significance relates to all the treatments within the lines.

Effect of IL17 in combination with TNFα, IL1, or OSM on the release of collagen from cartilage. Bovine nasal cartilage explants were cultured as described in fig 3 with IL17 (0.01–50 ng/ml) ± (A) TNFα (2 and 10 ng/ml), (B) IL1α (0.1 ng/ml) or OSM (10 ng/ml). Collagen release was determined as a measure of OHPro as described in “Materials and methods”, and the release by day 14 expressed as a percentage of the total (mean (SD)). Bonferroni's multiple comparison test was used as described in fig 3, where ***p≤0.001; *p≤0.05. For the selected comparisons shown, the statistical significance relates to all the treatments within the lines.
Effect of IL17 in combination with glycoprotein 130 binding cytokines on cartilage collagen release
OSM is a member of the glycoprotein 130 (gp130) binding cytokine family, which includes IL6, IL11, LIF, CNTF, and CT-1.33 To determine whether other family members might mimic OSM in synergising with IL17 to promote cartilage collagen breakdown, bovine nasal cartilage explants were incubated with IL17 in combination with the gp130 binding cytokines. Only OSM synergised with IL17 to induce collagen release (fig 5A), but the inclusion of sIL6R also enabled IL6 to promote synergistic cartilage collagen breakdown when in combination with IL17 (fig 5B).

Effect of IL17 in combination with gp130 binding cytokines on collagen release from cartilage. Bovine nasal cartilage explants were cultured as described in fig 1 with (A) IL17 (50 ng/ml) ± gp130 binding cytokines (OSM, 10 ng/ml; IL6, IL11, LIF, CNTF, CT-1, all at 50 ng/ml), or (B) IL17 (50 ng/ml) ± OSM (10 ng/ml) or IL6 (50 ng/ml) ± sIL6R (200 ng/ml). Collagen release was determined as a measure of OHPro as described in “Materials and methods”, and the release by day 14 expressed as a percentage of the total (mean (SD)). Bonferroni's multiple comparison test was used to compare IL17 alone with control, and IL17 + test reagent(s) with IL17 alone or IL17 + test reagent, where ***p≤0.001.
Effect of MMP inhibitors on the synergistic release of collagen from IL17-stimulated cartilage
Bovine nasal cartilage explants were cultured with IL17 ± TNFα (2 ng/ml) or IL1α (0.2 ng/ml) or OSM (10 ng/ml) in the presence or absence of TIMP-1 (100 U/ml) and the hydroxamate based metalloproteinase inhibitor BB-94 (10 μmol/l) for 14 days. The marked synergistic releases of collagen induced by these combinations were completely abrogated by the inclusion of TIMP-1 or BB94 (fig 6).

Effect of metalloproteinase inhibitors on the synergistic release of collagen from cartilage stimulated with IL17 in combination with TNFα, IL1, or OSM. Bovine nasal cartilage explants were cultured as described in fig 3 except that TIMP-1 (100 U/ml) or BB-94 (10 μmol/l) were included. (A) IL17 (50 ng/ml) was combined with TNFα (2 ng/ml), and (B) IL17 (20 ng/ml) was combined with IL1α (0.2 ng/ml) or OSM (10 ng/ml). Collagen release was determined as a measure of OHPro as described in “Materials and methods”, and the release by day 14 expressed as a percentage of the total (mean (SD)). Bonferroni's multiple comparison test was used to compare each cytokine combination with the same combination + inhibitor or control, where ***p≤0.001.
Effect of IL17 alone and in combination with proinflammatory cytokines on MMP expression in chondrocytes
Primary chondrocytes from bovine nasal cartilage were stimulated for 24 hours with IL17 in combination with TNFα, IL1α, OSM or IL6±sIL6R. Northern blot analyses showed that IL17 alone induced the expression of MMP-1, MMP-3, and MMP-13 mRNA over control levels at doses between 25 and 100 ng/ml (figs 7 and 8). MMP-13 expression was the most consistent and marked after stimulation with IL17 alone. TNFα (10 ng/ml) and IL1α (0.2 ng/ml) both synergised potently with IL17 (5–100 ng/ml) to markedly induce MMP-1, MMP-3, and MMP-13 expression; this induction was maximal even at 5 ng/ml IL17 (fig 7). IL17 (2–50 ng/ml) also synergised with OSM (10 ng/ml) to induce expression of these MMPs; maximal effects were achieved with IL17 at 50 ng/ml (fig 8A). This synergistic induction of MMPs was also seen with IL17 (50 ng/ml) in combination with IL6 (50 ng/ml) in the presence of sIL6R (200 ng/ml) (fig 8B). TIMP-1 mRNA levels were not significantly modulated by IL17 alone (figs 7 and 8). Only the combination of TNFα and IL17 (5–100 ng/ml) reduced TIMP-1 expression (fig 7). IL6 + sIL6R induced TIMP-1 expression when in combination with IL17 (50 ng/ml) (fig 8B). A minor induction of TIMP-1 mRNA was also seen with OSM in combination with IL17 at 50 ng/ml (but not at 2 and 10 ng/ml); OSM alone stimulated significant TIMP-1 expression (fig 8A).

Effects of IL17 in combination with TNFα or IL1α on MMP and TIMP mRNA expression in chondrocytes. Total cellular RNA (20 μg) from primary bovine nasal cartilage chondrocytes was harvested after stimulation for 24 hours with IL17 (5–100 ng/ml) ± TNFα (10 ng/ml) or IL1α (0.2 ng/ml) in serum free medium. GAPDH was used to assess RNA loading.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of IL17 in combination with OSM or IL6/sIL6R on MMP and TIMP mRNA expression in chondrocytes. Total cellular RNA (20 μg) from primary bovine nasal cartilage chondrocytes was harvested after stimulation for 24 hours with (A) IL17 (2–50 ng/ml) ± OSM (10 ng/ml), or (B) IL17 (50 ng/ml) ± IL6 (50 ng/ml) ± sIL6R (200 ng/ml) in serum-free medium. GAPDH was used to assess RNA loading.
DISCUSSION
Infiltration of T cells into the synovial joint is considered to be an early and important event in the pathogenesis of RA, but the role of these cells in cartilage destruction is not clearly understood. Recently, the T cell derived cytokine IL17 was shown to inhibit cartilage proteoglycan synthesis ex vivo, as well as induce its degradation both in vivo and ex vivo.18,21,22 This study confirms that IL17 can promote marked proteoglycan loss from cartilage, and for the first time demonstrates that IL17 can induce chondrocyte mediated cartilage collagen breakdown. This is particularly relevant because it has previously been shown that, although proteoglycan loss is reversible,12 the release of type II collagen from cartilage results in irreversible tissue damage.13 Collagen degradation, therefore, represents a key control point in cartilage turnover.
The collagen release induced by IL17 alone was less marked than that for proteoglycan release as is typically seen with other proinflammatory mediators.3,24,26 However, at suboptimal doses IL17 synergised potently with the proinflammatory cytokines TNFα and IL1 to promote cartilage breakdown. In addition, IL17 synergised with the gp130 binding cytokines OSM and IL6 (when in the presence of sIL6R). Raised levels of IL17 in RA synovial fluids have been reported,8 and these mean levels are in excess of the suboptimal IL17 doses used in this study. Furthermore, levels of TNFα, IL1, OSM, IL6, and sIL6R have also been shown to be raised in RA synovial fluids.1–3,34 Thus, given its ability to synergistically promote cartilage collagen breakdown ex vivo, the presence of IL17 within the proinflammatory milieu of the rheumatoid joint may contribute significantly to the destruction of cartilage in vivo. Anti-TNFα treatment has proved to be efficacious in the treatment of RA.35 Our data indicate that IL17 may also be an effective therapeutic target in disease, and given the redundancy and overlap that exists amongst cytokines, a combinatorial anticytokine approach including anti-IL17 and anti-TNFα, may be required. Indeed, blockade of endogenous IL17 has been shown to further inhibit the release of type I collagen from human synovium and bone after TNFα blockade.36
In addition to the direct effects of IL17 on cartilage, IL17 can induce the production of a number of cytokines, such as IL1, IL6, and TNFα, from various cell types present in the joint.9–11 Such production may then result in synergistic interactions between this T cell derived cytokine and these other proinflammatory mediators, leading to cartilage destruction. In this study we have also shown that IL17 potently synergises with several proinflammatory cytokines to promote the release of type II collagen from cartilage. Furthermore, IL17 can promote the chemoattraction of neutrophils through the secondary cytokine production of IL8.37 Raised levels of IL17 may, therefore, also contribute to the accumulation of neutrophils within the rheumatoid joint. This potential role for IL17 may also be significant because neutrophils can release OSM from intracellular stores upon activation.38
IL17-induced cartilage collagen loss was shown to be accompanied by an increase in collagenolytic activity that was inhibited by TIMP-1. Moreover, the collagen release induced by IL17, both alone and in combination with TNFα, IL1, and OSM, was abrogated by TIMP-1 and the broad spectrum metalloproteinase inhibitor BB-94. These findings clearly indicate that IL17-induced collagen release is MMP dependent. We have shown that IL17 can up regulate the expression of the collagenases MMP-1 and MMP-13 in chondrocytes, implicating them in this cytokine mediated catabolic process. MMP-13 expression was the more marked and consistently induced by IL17 alone, perhaps indicating a direct role for this collagenase in IL17-induced collagen degradation. This may be especially relevant given the preference of MMP-13 for type II collagen and its reported ability to cleave this macromolecule five to 10 times more efficiently than MMP-1.39 This is the first report of collagenase induction by IL17 in chondrocytes, although induction of the procollagenase activator MMP-3 has been previously shown11 and is confirmed in this study. This induction of MMP-3 may contribute significantly to IL17-induced cartilage collagen breakdown through the activation of proMMP-1 and proMMP-13.
The lack of an effect of IL17 alone on TIMP-1 mRNA expression in chondrocytes has also been shown in RA synoviocytes.40 Inhibition of TIMP-1 mRNA expression was seen with the combination of TNFα and IL17, a situation that would favour an increase in the net collagenolytic activity induced by this cytokine combination. IL17 also synergised with the gp130 binding cytokines OSM and IL6/sIL6R to induce both MMP and TIMP-1 expression in chondrocytes. A similar induction of TIMP-1 occurs after IL1 + OSM or IL6/sIL6R, but this is transient.24 Time course studies would confirm whether this transient effect is also found with IL17 in combination with OSM and IL6/sIL6R.
We have also clearly shown that anti-inflammatory molecules can influence the catabolic effects of IL17. Attenuation of the IL17-induced inhibition of proteoglycan synthesis by IL4 has been previously demonstrated.21 In addition, we have previously shown that IL1+OSM-induced cartilage collagen breakdown is inhibited by IL4, IL13, TGFβ1 and IGF1,25,30,41 and such observations also extend to TNFα-induced collagen release for TGFβ1 and IGF1.26 This study demonstrates that all these anti-inflammatory agents inhibited IL17-induced cartilage collagen breakdown. The inhibitory effects of TGFβ1 and IGF1 on IL1/OSM or TNFα appear to be through the inhibition of MMP expression,25,26,41 whereas the effects of IL4 and IL13 appear to be through the blockade of procollagenase activation.30 Similar mechanisms seem to be responsible for the inhibitory actions of these agents on IL17-induced collagen catabolism (data not shown).
In conclusion, this study has demonstrated several novel properties of the T cell derived cytokine, IL17, in that it can promote the release of type II collagen from cartilage. This process is chondrocyte mediated and MMP dependent. Moreover, IL17 can synergise with a number of other proinflammatory cytokines associated with RA through a synergistic induction of collagenase (MMP-1 and MMP-13) enzymes. The infiltration and activation of T cells into RA synovium is considered an early event in the disease process. Thus, we present further evidence to suggest a role for IL17 in the pathogenesis of RA, and propose that IL17 may act as a potent upstream mediator in inflammatory joint disease by initiating a proinflammatory cascade as well as directly promoting cartilage destruction.
Acknowledgments
We are indebted to British Biotech Pharmaceuticals Ltd for their generous supply of BB-94. We are grateful to Dr S Young-Min for assistance in the statistical analyses of the data. Aspects of this work were funded by the Arthritis Research Campaign, Glaxo-SmithKline, and the Wellcome Trust.