Article Text

Abstract
Background: It has previously been shown that many osteoclast precursors are included in the granulation tissue within the pseudocapsule obtained at revision arthroplasty from hips with osteolysis. In vitro culture of only cells isolated from the granulation tissue has been previously shown to generate many mature osteoclasts.
Objective: To investigate the presence or otherwise of supporting cells, similar to stromal cells, which differentiate osteoclasts within the granulation tissue.
Methods: Cells isolated from the granulation tissue were cultured alone, and after four weeks fibroblast-like cells (granulation fibroblasts) remained. Rat non-adherent bone marrow cells (NA-BMCs) were co-cultured with the granulation fibroblasts with or without 1α,25(OH)2D3 (10−8 M) or heat treated ROS 17/2.8 cell conditioned medium (ht ROSCM), or both. Multinucleated cells (MNCs), which formed, were assessed by biochemical and functional characterisation of osteoclasts. Receptor activator of NFκB ligand (RANKL) was investigated by immunohistochemistry.
Results: Co-culture of NA-BMCs and granulation fibroblasts caused the formation of tartrate resistant acid phosphatase (TRAP) positive MNCs, which had the calcitonin receptor (CTR), the Kat-1 antigen, which is specific to the surface of rat osteoclasts, and the ability to form pits in the presence of both 1α,25(OH)2D3 and ht ROSCM or in the presence of just ht ROSCM. RANKL was detected in fibroblast-like cells in the granulation tissue.
Conclusion: These data suggest that granulation fibroblasts support osteoclast differentiation, as do osteoblasts/stromal cells, and may play a part in aseptic loosening.
- aseptic loosening
- osteolysis
- osteoclasts
- fibroblasts
- CTR, calcitonin receptor
- FBS, fetal buffered saline
- ht ROSCM, heat treated ROS 17/2.8 cell conditioned medium
- mAb, monoclonal antibody
- α-MEM, α-minimum essential medium
- MNCs, multinucleated cells
- NA-BMCs, non-adherent bone marrow cells
- ODF, osteoclast differentiation factor, OPGL, osteoprotegerin ligand
- PBS, phosphate buffered saline
- PCR, polymerase chain reaction
- RANKL, receptor activator of NFκB ligand
- RT, reverse transcriptase
- [125I]sCT
- [125I]salmon calcitonin
- SEM, scanning electron microscopy
- TRAP, tartrate resistant acid phosphatase
Statistics from Altmetric.com
- CTR, calcitonin receptor
- FBS, fetal buffered saline
- ht ROSCM, heat treated ROS 17/2.8 cell conditioned medium
- mAb, monoclonal antibody
- α-MEM, α-minimum essential medium
- MNCs, multinucleated cells
- NA-BMCs, non-adherent bone marrow cells
- ODF, osteoclast differentiation factor, OPGL, osteoprotegerin ligand
- PBS, phosphate buffered saline
- PCR, polymerase chain reaction
- RANKL, receptor activator of NFκB ligand
- RT, reverse transcriptase
- [125I]sCT
- [125I]salmon calcitonin
- SEM, scanning electron microscopy
- TRAP, tartrate resistant acid phosphatase
Aseptic loosening is a major cause of late failure of total joint arthroplasty. Its causes comprise mechanical factors, including micromotion of the prosthesis and stress shielding, and biological factors that involve wear particles. Generation of wear particles has emerged as the major problem in total joint arthroplasty, as was first recognised by Charnley.1 He linked massive granuloma formation in response to Teflon particles to early clinical failures of total joint arthroplasty. Since that time, clinical reports have implicated the biological response to wear particles as a causative factor in aseptic loosening.
The biological response of varying cell types to wear particles has been investigated. Charnley noted that macrophages are present at the interface between the bone and cement.1 Willert demonstrated foreign body reaction to particles of plastic, metal, and acrylic that results in the formation of granulation tissue containing macrophages and giant cells.2 Subsequently, histomorphological analysis of the interface membrane from revision hips has shown that the cell types are diverse, including macrophages, fibroblasts, T cells, and multinucleated giant cells.3–8 This inflammatory response is thought to be associated with foreign body macrophages in reaction to wear particles. It is well recognised that macrophages that have phagocytosed wear particles then produce humoral factors—for example, cytokines, growth factors, and prostaglandins, which are known to activate osteoblasts to stimulate osteoclastic bone resorption in vitro.9–18 Recently, osteoclasts have been thought to be the main cells in bone resorption of aseptic loosening.19–29 In general, cell to cell contact between osteoblasts/stromal cells and osteoclast precursors is a necessary step for osteoclast differentiation.30 Moreover, osteoblasts/stromal cells support osteoclast activation in vitro.31 In the granulation tissue from revision hips with aseptic loosening, the mechanisms of osteoclast differentiation are controversial. Quinn et al have shown that under co-culture with mouse osteoblastic cells, mouse inflammatory foreign body macrophages which are derived from granulomas formed in response to subcutaneously implanted polymethylmethacrylate particles can differentiate into osteoclast like cells.22,32 Sabokbar et al have shown that human macrophages that are isolated from the pseudomembrane obtained at revision total hip arthroplasty differentiated into osteoclast-like cells under co-culture with mouse osteoblastic cells.25 These data indicate that wear particles associated with macrophages may contribute to osteolysis in the failed arthroplasty by differentiating into mature osteoclasts under co-culture with osteoblastic cell lines. However, the types of cells which can support osteoclast differentiation in the granulation tissue from revision hips with aseptic loosening are still unknown.
We have previously shown that many osteoclast precursors are contained within the granulation tissue inside the pseudocapsule from revision hips with osteolysis.33 After culturing the isolated cells alone in vitro, osteoclast-like cells appeared in the culture wells. This seems to suggest that the cells which support osteoclast differentiation exist within the granulation tissue. The purpose of this study was to investigate the presence of cells like osteoblasts/stromal cells that can support osteoclast differentiation in granulation tissue which does not contain bone inside the pseudocapsule.
PATIENTS AND METHODS
Patients and samples
The inner granulation tissue of the pseudocapsule was harvested, far from the bone tissue, from seven patients undergoing surgery in revision hips with osteolysis (three men, four women, average age 73). Radiographs of all samples were taken before the subsequent procedure using Softex (Osaka Softex Sales Co, Osaka, Japan) to confirm that the granulation tissue did not include bone tissue. Skin scars after surgery were obtained as control samples from two patients (one man, one woman average age 74.5).
Cell culture
Preparation of cells from the pseudocapsule
The granulation tissue was cut into small pieces, and these were digested in phosphate buffered saline (PBS) containing 2 mg/ml collagenase for two hours at 37°C. The isolated cells were suspended in α-minimum essential medium (α-MEM) (Gibco Grand Island, NY) containing 10% fetal buffered saline (FBS; Gibco Grand Island, NY). To exclude osteoclast progenitor cells from the isolated cells, the cells were cultured for four passages. They were then fixed and stained for tartrate resistant acid phosphatase (TRAP) to confirm that there were no osteoclast progenitor cells. Then, the cells were stored in α-MEM containing 10% dimethyl sulphoxide at −80°C as granulation fibroblasts for further experiments.
Preparation of non-adherent bone marrow cells
Bone marrow cells were harvested from male Sprague-Drawley rats (4–7 weeks old) (SEAC Yoshitomi, Fukuoka, Japan). To deplete stromal cells from the bone marrow cells, the cells were passed through a Sephadex G-10 column, as described previously.34,35 The cells that were passed through the column were harvested as non-adherent bone marrow cells (NA-BMCs).
Co-culture system
The rat osteosarcoma cell line ROS 17/2.8 was kindly provided by Dr L Bonewald (University of Texas Health Science Centre, San Antonio). Conditioned medium of ROS 17/2.8 cells was concentrated 10 times by a Cetriprep-10 filter (AMICON Corp, Danvers, MA). The conditioned medium was treated at 95°C for five minutes, and heat treated ROS 17/2.8 cell conditioned medium (ht ROSCM) was added to the cultures at a concentration of 10% (vol/vol) to obtain a rich population of preosteoclasts according to the previous study.36 After the NA-BMCs were cultured in 24 well culture plates (1×106/well) in α-MEM containing 15% FBS with 1α,25-dihydroxyvitamin D3 (1α,25(OH)2D3) (10−8 M) (Biomol Plymouth Meeting, PA) and ht ROSCM for four days, TRAP positive mononuclear cells were observed. Then, the granulation fibroblasts (5×104/well) in α-MEM containing 15% FBS were seeded to the culture well of the previously seeded NA-BMCs in the supplementation of 1α,25(OH)2D3 (10−8 M) or ht ROSCM, or both.
Characterisation of cultured cells
TRAP staining
Cultured cells were co-cultured for 2–14 days, then fixed, and the TRAP in the cells was stained (Sigma Chemicals, St Louis, MO). Cells containing three or more nuclei were counted as multinucleated cells (MNCs). TRAP positive MNCs were assessed as osteoclast-like cells.
Calcitonin receptor autoradiography
The calcitonin receptor (CTR) is known to be a specific marker present on preosteoclasts and mature osteoclasts. The appearance of CTR in cultured cells was assessed by autoradiography using [125I]salmon calcitonin ([125I]sCT) (Amersham, Buckinghamshire, UK). The cells were incubated for one hour at room temperature with 1 μCi/ml (1.77 ng/ml) [125I]sCT in the absence or presence of excess unlabelled sCT (500-fold). The cells were rinsed three times with serum-free α-MEM containing 0.1% bovine serum albumin, then fixed with 0.1 M cacodylate buffer (pH 7.3) containing 2% formaldehyde and 2% glutaraldehyde for 10 minutes. The cells were washed with 0.1 M cacodylate buffer once and processed for TRAP staining. The bottom of each well was cut out from the culture plates, dipped in NR-M2 emulsion (Konica Co, Tokyo, Japan), and air dried, followed by exposure for seven days at 4°C. The autoradiographs were developed with Konicadol (Konica) and fixed with Konifix (Konica).
Immunostaining using anti-Kat-1 antigen monoclonal antibody (mAb Kat-1)
Kat-1 antigen is a unique cell surface protein for rat osteoclasts.37 Cultured cells were stained with mAb Kat-1 at 4°C for 45 minutes. The cells were fixed with 2% paraformaldehyde in PBS and blocked with 3% goat serum in PBS, then allowed to react with biotinylated antimouse IgM antibodies for 30 minutes. The cells were processed for the avidin-biotin complex procedure using an ABC-AP kit (Vector Laboratories, Burlingame, CA). The endogenous alkaline phosphatase activity was blocked with 1 mM levamisole. Then, the cells were fixed with 5% glutaraldehyde in PBS.
Detection of pit formation using scanning electron microscopy
Bone resorption is a distinctive function of mature osteoclasts. Dentin slices with a thickness of 10–15 μm were made from ivory and stored in 70% ethanol. The cells attached on dentin slices were cultured for seven days, then fixed and processed for TRAP staining. The cells were removed from the slice by dipping the slices in 0.25 M NH4OH followed by sonication. The slices were air dried and mounted on scanning electron microscopy (SEM) stubs. They were platinum-sputtered and examined by SEM.
Reverse-transcription (RT)-PCR
The granulation fibroblasts were cultured in α-MEM containing 15% FBS with or without 1α,25(OH)2D3 (10-8 M) and/or ht ROSCM for four days. Total RNA was isolated according to the single step guanidinium thiocyanate-phenol-chloroform extraction procedure using Isogen. For a reverse transcriptase (RT) and polymerase chain reaction (PCR), a Perkin Elmer/Cetus DNA thermal cycler was used. After denaturation at 65°C for 10 minutes, cDNA was produced by reverse transcription at 42°C for one hour (5 μg of total RNA in 20 μl of 50 mM Tris-HCl, 8 mM MgCl2, 30 mM KCl, 1 mM of dNTP mixture, 25 U RNase inhibitor, and 25 U reverse transcriptase).
PCR was performed as follows. Briefly, the reaction mixture contained 20 pM of each primer, 100 mM Tris-HCl, 15 mM MgCl2, 500 mM KCl, 2.5 mM of dNTP mixture, 0.5 U Taq DNA polymerase, and sterile distilled water up to 50 μl. The oligonucleotide primers used were CTR (5′ to 3′: forward GCAATGCTTTCACTCCTGAGAAAC, reverse CAGTAAACACAC AGCCACGACAATGAG),38 and ribosomal RNA (5′ to 3′: forward TTACCAAAAGTGGCCCACTA, reverse GAAAGATG GTGAACTATGCC).39 PCR amplification was performed with cycles of denaturation at 94°C for one minute, annealing at 56°C (CTR) or 66°C (rRNA) for one minute, and extension at 72°C for one minute. CTR has two isoforms by alternative splicing. The primers for CTR detected both of them. The amplified fragments were 346 bp, 412 bp (CTR), and 345 bp (rRNA).
Immunostaining using antibody anti-receptor activator of NFκB ligand (RANKL)/osteoclast differentiation factor (ODF)/osteoprotegerin ligand (OPGL)
The granulation tissues were fixed with 4% paraformaldehyde in PBS overnight at 4°C and embedded in paraffin. An immunohistochemical study was carried out using Vectastain ABC-PO kit (Vector Laboratories, Burlingame, CA). Briefly, endogenous peroxidase activity was removed by a 60 minute incubation in methanol containing 0.3% H2O2. Non-specific immunoglobulin binding was blocked by a 15 minute incubation of sections in PBS containing 5% normal rabbit serum. Goat antihuman RANKL polyclonal antibody (Santa Cruz Biotechnology Inc, Santa Cruz, CA) was diluted at a concentration of 1 μg/ml and incubated overnight at 4°C. After extensive washing, the sections were incubated with biotinylated rabbit antigoat IgG and avidin-enzyme complex. Antibody binding was visualised by incubating the slides in 3,3′-diaminobenzidine and hydrogen peroxide.
Statistics
Two way analysis of variance followed by Tukey's test was used to determine the effect of supplementation (presence of just ht ROSCM v presence of 1α,25(OH)2D3 plus ht ROSCM) and number of days (0, 2, 4, 7, 10, 14) on the number of TRAP positive MNCs over the time course (SigmaStat, version 2.0, SPSS Inc, Chicago, IL).
RESULTS
Preparation of the granulation fibroblasts
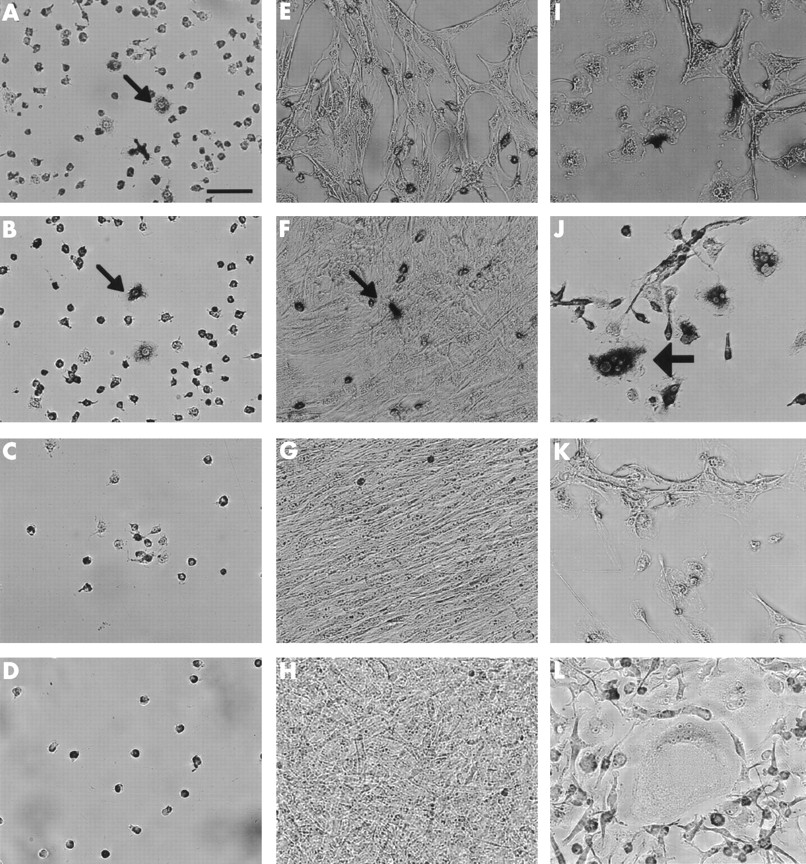
Cells isolated from the inner granulation tissue were composed of monocytes, macrophage-like cells, and fibroblast-like cells. After one week of culture, the isolated cells reached confluence, and TRAP positive MNCs were visible (fig 1A). When the isolated cells were cultured on a dentin slice for one week, the TRAP positive MNCs formed pits on the dentin slice. In the same culture conditions, the isolated cells came to be composed of only fibroblast-like cells (granulation fibroblasts) after four passages of the culture, and no TRAP positive MNCs or mononuclear cells were seen (fig 1B). To confirm that osteoclast precursors had disappeared from the culture, the expression of CTR mRNA was examined in the cultured cells using RT-PCR. No transcript for CTR mRNA was found (fig 2). Variable supplementation in the culture did not influence the results.

Preparation of granulation fibroblasts. Cells isolated from the granulation tissue staining for TRAP (A) after culture for one week and (B) after culture for four weeks. Bar = 10 μm. TRAP positive MNCs (arrow) appeared in the culture of the isolated cells from the granulation tissue after culture for one week (A). After culture for four weeks, neither TRAP positive mononuclear cells nor MNCs were seen, only the granulation fibroblasts remaining in the culture (B).

Expression of CTR mRNA in granulation fibroblasts. Expression of CTR mRNA by RT-PCR (50 cycle). Lane 1, giant cell tumour tissue (positive control); lane 2, granulation tissue; lane 3–6, granulation fibroblast culture (lane 3, with no supplementation; lane 4, with 1α,25 (OH)2D3; lane 5, with ht ROSCM; lane 6, with 1α,25 (OH)2D3 and ht ROSCM). Giant cell tumour tissue and granulation tissue showed two signals (346 bp and 412 bp) of CTR, but the culture of granulation fibroblasts alone did not show any signal under any of the conditions.
Osteoclast differentiation supporting the activity of granulation fibroblasts
To examine the supporting activity for osteoclast differentiation of the granulation fibroblasts, they were co-cultured with rat NA-BMCs as a source of osteoclast progenitor cells. In the co-cultures supplemented with ht ROSCM, TRAP positive MNCs appeared on day 2, and the number peaked on day 4 (figs 3 and 4). There was no significant difference in the number of TRAP positive MNCs between co-cultures supplemented with 1α,25(OH)2D3 and ht ROSCM and co-cultures supplemented with just ht ROSCM. In the co-cultures supplemented with just 1α,25(OH)2D3 , or with no supplementation at all, no TRAP positive MNCs were formed. Furthermore, no TRAP positive MNCs appeared in the co-cultures of rat NA-BMCs and human skin fibroblasts, or in the cultures of NA-BMCs alone, or of the granulation fibroblasts alone.

Stromal cell-like activity of granulation fibroblasts. Three different groups of cells were cultured; NA-BMCs alone (A–D), NA-BMCs and skin fibroblasts (E–H), NA-BMCs and granulation fibroblasts (I–L). This figure shows cultured cells stained by TRAP on day 2 (A, E, I), day 4 (B, F, J), day 7 (C, G, K), or day 14 (D, H, L). Bar = 10 μm. In the culture of NA-BMCs alone and NA-BMCs and skin fibroblasts, TRAP positive mononuclear cells (small arrows) survived until day 4 (A, B, F). Only in the culture of NA-BMCs and granulation fibroblasts, could we observe TRAP positive MNCs (large arrows) (J).
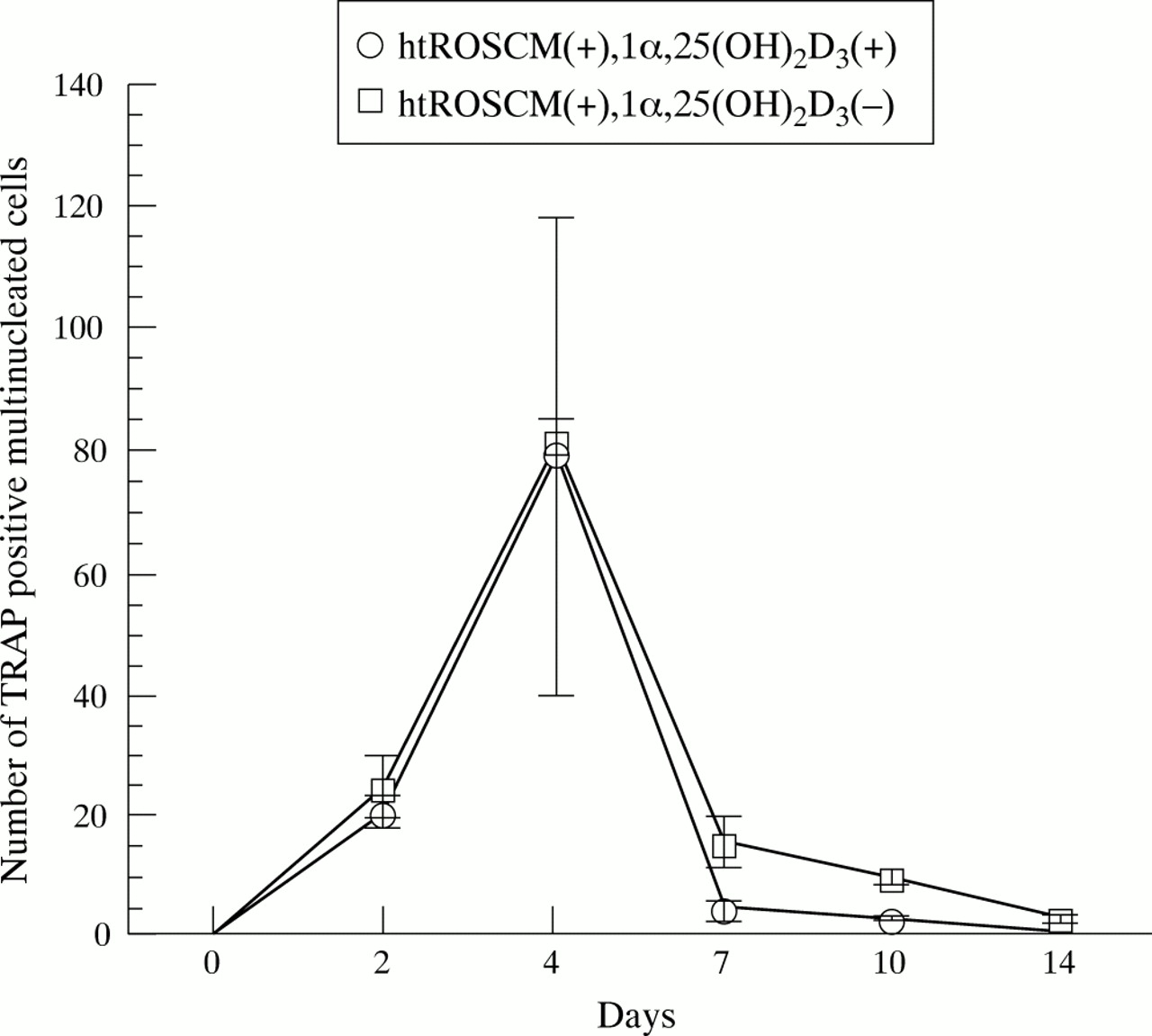
Effect of 1α,25(OH)2D3 supplementation on the number of TRAP positive MNCs. In the co-culture of NA-BMCs and granulation fibroblasts in the presence of 1α,25(OH)2D3 and ht ROSCM or in the presence of just ht ROSCM, TRAP positive MNCs appeared at two days and the number of TRAP positive MNCs increased until four days, reaching a maximum, before decreasing with time until the end of the co-culture. There was no significant difference in the number of TRAP positive MNCs (in the presence of just ht ROSCM compared with the presence of 1α,25(OH)2D3 and ht ROSCM).
Functional and biochemical characterisation of the formed MNCs
Radiolabelled CT became bound to the TRAP positive MNCs in the co-culture of rat NA-BMCs and the granulation fibroblasts supplemented with ht ROSCM (fig 5A). This binding was inhibited by the addition of excess unlabelled CT, indicating that CTR existed on the TRAP positive MNCs (fig 5B). To confirm the origin of the MNCs formed, the reactivity of the cells to monoclonal antibodies against Kat-1 was investigated. The MNCs formed were positive for Kat-1 by immunohistochemistry, indicating that they were from rat bone marrow cells (fig 6A) On the contrary, there were no positive cells in either the cell culture from human giant cell tumour or from the inner granulation tissue (fig 6B). When the granulation fibroblasts and rat NA-BMCs were co-cultured on a dentin slice supplemented with ht ROSCM, deep pits were observed under an SEM (fig 7A). No pits were observed in the cultures of NA-BMCs alone or the granulation fibroblasts alone (fig 7B).
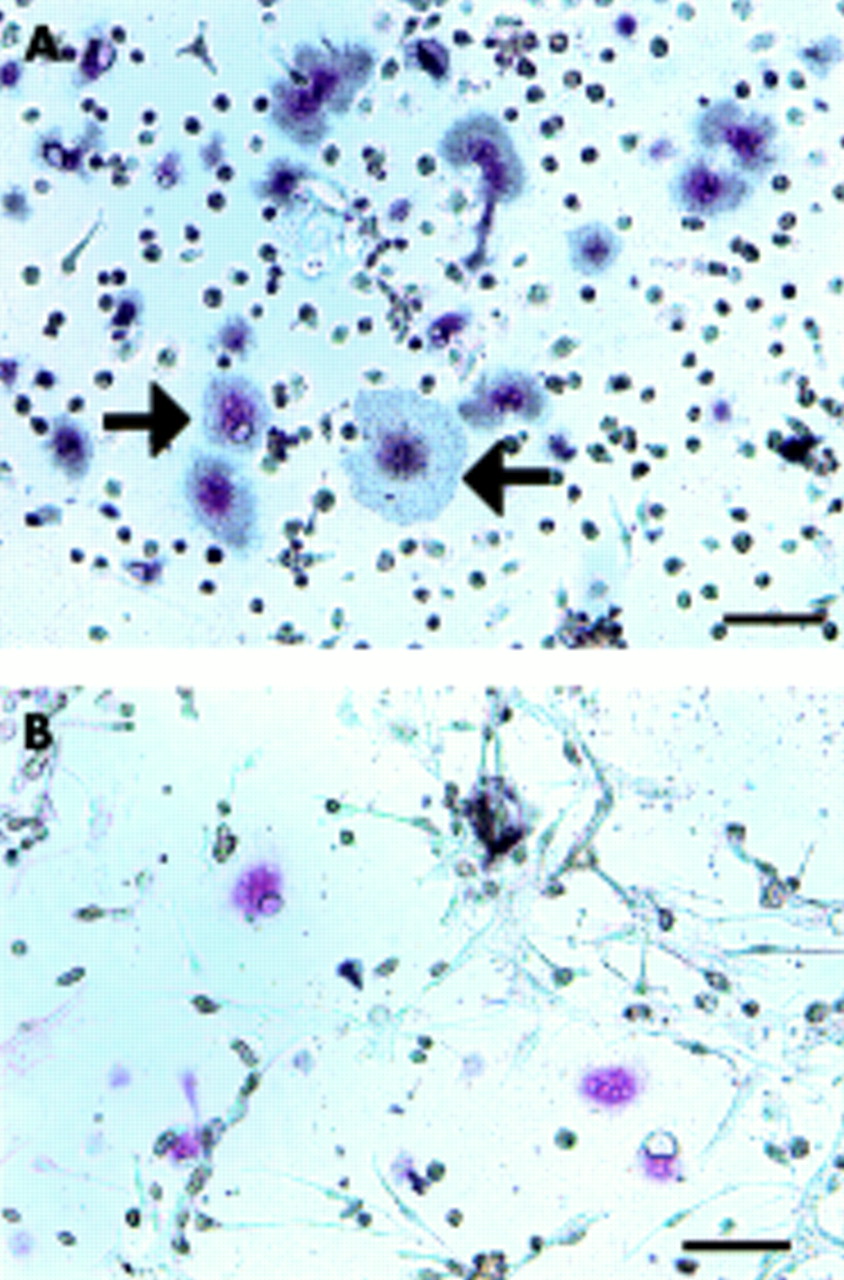
Presence of CTR on the TRAP positive MNCs formed. The cells cultured with 1α,25(OH)2D3 and ht ROSCM for four days were incubated with [125I]sCT for one hour, either without (A) or with (B) an excess amount of unlabelled sCT, followed by slight staining for TRAP. Bar = 10 μm. Grains were seen over TRAP positive MNCs (black arrow) and TRAP positive mononuclear cells (A). No grains were detected over TRAP positive MNCs or TRAP positive mononuclear cells in the same culture system when the cultured cells were incubated with an excess amount of unlabelled sCT (B).

Immunostaining by mAb Kat-1. The cultured cells (A) and human osteoclast-like cells from the giant cell tumour (B) which had been cultured for four days were stained with mAb Kat-1 followed by detection, using an ABC-AP kit. Bar = 10 μm. The MNCs formed were stained with mAb Kat-1 (A). Human osteoclast-like cells were unreactive with mAb Kat-1 (B).

Pit formation by SEM. The cultured cells (A) and NA-BMCs alone (B) were cultured on a dentine slice for seven days. Pits were formed by the cultured cells (A). In the culture of NA-BMCs, pits were not seen (B).
Localisation of RANKL/ODF/OPGL in the granulation tissue
RANKL was detected in vascular cells and fibroblast-like cells of the granulation tissues (fig 8). However, there were some fibroblast-like cells with no staining of RANKL.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Localisation of RANKL in the granulation tissues. The granulation tissues inside the pseudocapsule were immunostained by anti-RANKL antibody (A) and control (B). Bar = 10 μm. RANKL was localised at vascular cells (arrow) and fibroblast like cells (arrow heads).
DISCUSSION
The granulation fibroblasts supported osteoclast precursors in rat NA-BMCs to differentiate TRAP positive MNCs. The MNCs had CTR and Kat-1, which is a unique cell surface protein for rat osteoclasts, on the cell surface, and had resorption activity on a dentine slice. These findings indicated that the MNCs formed were mature rat osteoclasts. This means that the granulation fibroblasts had the supporting activity of osteoclast differentiation.
For two reasons, we have been using the inner granulation tissue of the pseudocapsule as a cell source, rather than the granulation tissue at the osteolysis lesion between failed components and bone tissue. Firstly, the inner granulation tissue is supposed to be rich in wear particles because it is close to the joint at which wear particles are mostly generated. Therefore, many cells which are influenced by wear particles either directly or indirectly are expected to exist within the inner granulation tissue. Secondly, the inner granulation tissue of the pseudocapsule has a very low possibility of becoming contaminated with bone or bone marrow, which may include osteoblasts or bone marrow stromal cells.
The inner granulation tissue was harvested far from the bone tissue to exclude contamination of cells from bone tissue. This was confirmed by radiographs of the tissue before any subsequent treatment. Furthermore, in the culture of NA-BMCs alone, no TRAP positive MNCs were formed. These data support the fact that there was no contamination of osteoblasts within the granulation tissue and no contamination of stromal cells in the NA-BMCs. Therefore, the cells which showed stromal cell-like activity originated from within the inner granulation tissue.
In the culture of the granulation fibroblasts alone, no TRAP positive MNCs or mononuclear cells were observed. Furthermore, no transcript for CTR mRNA was detected in the culture by RT-PCR, indicating that osteoclasts or immediate osteoclast precursors were absent. Furthermore, the formed MNCs had Kat-1, which is a cell surface protein of rat osteoclast lineage cells. These data support the notion that the osteoclast-like cells formed were originally from rat NA-BMCs.
In general, bone resorbing factors, such as 1α,25(OH)2D3, are necessary for in vitro osteoclast differentiation, in addition to osteoblasts or other specific bone derived stromal cells. This paper showed that cells which had osteoclast differentiation supporting activity were included among the isolated cells from the inner granulation tissue of hips which had undergone total hip arthroplasty but failed with osteolysis. However, 1α,25(OH)2D3 was not necessary to induce TRAP positive MNCs in our co-culture system. Kukita et al have shown that rat osteoblasts activated by 1α,25(OH)2D3 are required for a multinucleation step of rat osteoclast differentiation.36 Rat osteoclast progenitor cells can be differentiated into TRAP positive mononuclear cells without supplementation of 1α,25(OH)2D3 (presence of just ht ROSCM). One of the reasons why TRAP positive MNCs formed without 1α,25(OH)2D3 in our co-culture system is that the species of supporting cells for osteoclast differentiation may differ between rat osteoblasts and human granulation fibroblasts. Another possible reason is that the supporting mechanism by granulation fibroblasts may differ from that by osteoblasts/stromal cells.
Current studies show that osteoblasts or stromal cells derived from bone marrow express ODF or OPGL.40,41 This mediates an essential signal to osteoclast progenitor cells for their differentiation into osteoclasts, whereas osteoclast inhibitory factor or osteoprotegerin inhibits osteoclast differentiation by interrupting cell to cell signalling between osteoclast precursors and osteoblasts or stromal cells.42,43 Recent study suggested that macrophages isolated from the pseudomembrane surrounding loose arthroplasty components can differentiate into osteoclastic bone resorbing cells in the presence of RANKL/ODF/OPGL, and osteoclast inhibitory factor/giant cell tumour inhibits this process in a dose dependent fashion.44 Our data suggest that RANKL/ODF/OPGL is expressed in the granulation tissues and may play a part in osteoclast differentiation.
Information on the role of fibroblasts in the revision of hips which have undergone total hip arthroplasty, but failed with osteolysis, is limited.45–47 Most such studies have dealt with cytokines, metalloproteinases, and collagens which the fibroblasts from interfacial membrane produced, or with the growth of the interfacial membrane. As far as we know this study is the first report which shows that the granulation fibroblasts from the granulation tissue inside the pseudocapsule can support osteoclast differentiation.
Previously, we have shown that the inner granulation tissues also include osteoclast progenitor cells. This suggests that the granulation tissue may provide osteoclasts which induce local bone resorption, or osteolysis, near the joints. Osteolysis near the joints is a critical step of the component loosening. This would cause bone defect around the proximal portion of the femoral component, resulting in enlargement of the gap around the femoral component. It would enable wear particles from the joint to reach deep within the bone tissue around the component, resulting in expansion of the osteolysis around the femoral component. The granulation fibroblasts and the osteoclast precursors within the inner granulation tissue may also play a part in osteolysis around the sockets.
This study also suggests that osteoclasts have a role in osteolysis, which causes aseptic loosening, and supporting cells are necessary for osteoclast differentiation. The supporting activity of granulation fibroblasts would be a potential target for prohibiting the progression of osteolysis.
Acknowledgments
The English used in this manuscript was revised by Miss K Miller (Royal English Language Centre, Fukuoka, Japan).