Article Text

Abstract
OBJECTIVE To investigate the effect of insulin-like growth factor 1 (IGF1) on the release of collagen, and the production and expression of matrix metalloproteinases (MMPs) induced by the proinflammatory cytokine interleukin 1α (IL1α) in combination with oncostatin M (OSM) from bovine nasal cartilage and primary human articular chondrocytes.
METHODS Human articular chondrocytes and bovine nasal cartilage were cultured with and without IGF1 in the presence of IL1α or IL1α + OSM. The release of collagen was measured by an assay for hydroxyproline. Collagenase activity was determined with the diffuse fibril assay using3H acetylated collagen. The expression of MMP-1, MMP-3, MMP-8, MMP-13, and tissue inhibitor of metalloproteinase 1 (TIMP-1) mRNA was analysed by northern blot.
RESULTS IGF1 can partially inhibit the release of collagen induced by IL1α or IL1α + OSM from bovine nasal cartilage. This was accompanied by a reduced secretion and activation of collagenase by bovine nasal cartilage. IGF1 can also down regulate IL1α or IL1α + OSM induced MMP-1, MMP-3, MMP-8, and MMP-13 mRNA expression in human articular chondrocytes and bovine chondrocytes. It had no significant effect on the production and expression of TIMP-1 mRNA in chondrocytes.
CONCLUSION This study shows for the first time that IGF1 can partially block the release of collagen from cartilage and suggests that down regulation of collagenases by IGF1 may be an important mechanism in preventing cartilage resorption initiated by proinflammatory cytokines.
- collagen
- metalloproteinases
- cytokines
- insulin-like growth factor 1
Statistics from Altmetric.com
Cartilage is a highly organised extracellular matrix in which proteoglycan (PG) and collagen fibrils are the two major structural components. PGs are complex, highly charged macromolecules with a high turnover rate, which pull water into the tissue and allow it to resist compression. The intact collagen network provides a permanent framework that gives tensile strength, and allows PG to be held in the matrix. Collagen has a low turnover rate, and when degradation of collagen does occur the structural integrity of the tissue is irreversibly lost.1 ,2 The degradation of PG and collagen is a prominent feature of the cartilage loss seen in both rheumatoid arthritis (RA) and osteoarthritis (OA), and ultimately leads to joint destruction.
Chondrocytes within cartilage play an important part in maintaining the surrounding matrix by modulating the synthesis and degradation of the various components in both normal and pathological conditions. During degenerative joint diseases, such as RA and OA, chondrocytes elicit a catabolic response which exceeds anabolism of new matrix molecules.1 The expression and production of proteolytic enzymes is increased and a major role is indicated for matrix metalloproteinases (MMPs), which can degrade matrix components.3 ,4
Current knowledge points to the involvement of MMPs in the pathological destruction of cartilage in arthritis. Collagens can be specifically cleaved by the collagenases. Three mammalian collagenases (MMP-1, MMP-8, and MMP-13) can directly cleave the Gly975-Leu976 bond on the α1(II) chain, generating characteristic three quarter and one quarter length fragments.5 ,6 After this cleavage the collagen fragments become susceptible to further degradation by other enzymes, such as stromelysin-1 (MMP-3) and gelatinases A and B (MMP-2 and MMP-9). MMP-3 is another potent enzyme in the turnover of cartilage components, in that it can cleave the N-telopeptide of type II collagen, which may lead to depolymerisation of type II collagen.7 ,8 MMP-3 is also an activator of other MMPs, such as MMP-1 and MMP-9.9 ,10 All these enzymes are expressed by chondrocytes.3 ,11 These potent enzymes are controlled at key points that include the stimulation of synthesis and secretion by cytokines and growth factors, such as interleukin 1 (IL1) and tumour necrosis factor α, and the activation of proenzyme forms.3 ,12 Tissue inhibitors of metalloproteinases (TIMPs), which are also expressed by chondrocytes, suppress the biological activity of collagenases.13 ,14
Insulin-like growth factor (IGF) is a polypeptide with structural and functional homology to proinsulin.15 Human IGF1 and IGF2 play a part in growth and development of many tissues during fetal and adult life and are thus implicated in tissue hypertrophy and repair. IGF1 is known to enhance the synthesis of matrix in normal cartilage in vivo and in vitro and is recognised as one of the most important growth factors affecting the metabolism of matrix molecules found in cartilage. This growth factor enhances the synthesis of PG and collagen by chondrocytes in cartilage and maintains expression of cartilage type II collagen in the absence of serum without increasing the synthesis of type I collagen.16-18 IGF1 also blocks IL1 stimulated PG degradation and reverses the suppression of a cartilage-specific phenotype induced by IL1.19-22 Thus it is also an important regulator in repair processes during joint disease.23 In bone IGF1 has been found to stimulate collagen synthesis and inhibit collagenase mRNA expression in bone cells and osteoblasts.24-26 The expression and synthesis of both IGF and IGF binding proteins (IGFBPs) are increased in arthritic chondrocytes.27 However, the precise mechanism by which IGF1 blocks cartilage breakdown induced by proinflammatory cytokines is not clear. Little information is available about the effect of this growth factor on the expression and production of MMPs induced by proinflammatory cytokines, such as IL1α, in cartilage.
We have recently shown that the combination of IL1α and oncostatin M (OSM) promotes a dramatic release of PG and collagen fragments from bovine, porcine, and human cartilage explants in culture. This synergistic release of collagen is accompanied by a marked up regulation of procollagenases, and their subsequent activation as well as a reduction in TIMP-1.28 ,29 Cytokines such as IL4 and transforming growth factor β1 (TGFβ1) can block these effects.30 ,31
In this study we investigated whether, and to what degree, IGF1 influences the release of collagen fragments from bovine nasal cartilage stimulated with IL1α in combination with OSM, and determined the possible mechanisms involved in the regulation of collagenases.
Material and methods
Chemicals were obtained from the following suppliers: Human recombinant IL1α was a generous gift from Glaxo Group Research Ltd (Greenford, UK). Human recombinant OSM was generously supplied by Professor John Heath, Department of Biochemistry, Birmingham, UK. Human recombinant IGF1 was purchased from R&D Systems (Abingdon, UK). All other chemical and biochemicals were commercially available analytical grade reagents obtained from Fisons (Loughborough, UK) or Merck (Poole, UK) or have been previously described.28 ,31
CARTILAGE DEGRADATION ASSAY
Control culture medium was Dulbecco's modification of Eagle's medium (DMEM) containing 25 mM HEPES (Gibco, Paisley, UK) supplemented with glutamine (2 mM), streptomycin (100 μg/ml), penicillin (100 U/ml), and amphotericin B (2.5 μg/ml). Bovine nasal septum cartilage was held at 4°C overnight after slaughter. Discs were punched from 2 mm slices to give pieces 2 mm in diameter, which were washed twice in Dulbecco's phosphate buffered saline (DPBS). Three discs per well of a 24 well plate were incubated at 37°C in control medium (600 μl) for 24 hours. Fresh control medium (600 μl), with or without test reagents (four wells for each condition), was then added and the plate incubated at 37°C for seven days. The supernatants were harvested and replaced with fresh medium containing identical test reagents to day 1. The experiment was continued for a further seven days. Day 7 and 14 supernatants and the remaining cartilage discs were stored at −20°C until assayed.29The viability of cartilage explants was assessed by screening for the production of lactate dehydrogenase (LDH) using the Cytotox 96 assay (Promega, Southampton, UK). This was always performed when new cytokine combinations were used with explants. No increase in LDH levels with any of the cytokine combinations was seen (data not shown). Serum is excluded from cartilage explants because it can markedly alter the metabolism of cartilage in the absence of exogenous cytokine(s).32 The absence of serum does not affect the viability of cartilage under these conditions and previous studies have shown that cartilage in serum-free culture for eight to nine days can respond to serum and other growth factors,33 and IGF1, which is present in serum, can also replace it.34 As this study investigated the effect of IGF1 on cartilage, serum was excluded from the explant culture system.
PROTEOGLYCAN AND COLLAGEN ASSAY
Media samples and papain digests of the remaining cartilage were assayed for sulphated glycosaminoglycan (as a measure of PG release) using the 1,9-dimethylmethylene blue dye binding assay.35Hydroxyproline release (as a measure of collagen degradation) was assayed by a modification of the method as described.29Mean (SD) total hydroxyproline measured in these experiments was 496.7 (112.0) ng/well.
COLLAGENASE AND TIMP ACTIVITY
Collagenase activity was determined with the diffuse fibril assay using 3H acetylated collagen.36 Aminophenyl mercuric acetate (APMA) was added at 0.7 mmol/l to activate procollagenases. Assays without APMA gave an estimate of active collagenase, whereas inclusion of APMA allowed total collagenase (active + procollagenase) levels to be measured. Inhibitory activity was assayed by the addition of samples to a known amount of active collagenase in the diffuse fibril assay. This assay measures free TIMP activity but does not detect TIMP complexed to MMP. One unit of collagenase activity degrades 1 μg of collagen per minute at 37°C and one unit of TIMP inhibits two units of collagenase by 50%.
CHONDROCYTE CULTURE
Human articular cartilage samples were collected from three patients with OA (age 65 (6)) who were undergoing total knee joint replacement surgery in a local hospital. These samples were predominately taken from areas of cartilage that appeared macroscopically normal. Both bovine and human cartilage were cut into small pieces, and treated by sequential enzyme digestion. The cartilage was first incubated at 37°C for 15 minutes with hyaluronidase at 1 mg/ml in DPBS. After three washes with DPBS, cartilage was incubated with 0.25% (w/v) trypsin in DPBS for 30 minutes at 37°C. The cartilage pieces were then incubated overnight with 3 mg/ml bacterial collagenase in DPBS at 37°C. The chondrocytes were collected by centrifugation at 1100 rpm for five minutes. After washing three times with DPBS, chondrocytes were incubated in DMEM containing 10% (v/v) fetal calf serum at 1 × 106 cells/T-25 cm2tissue culture flask in 3 ml of culture medium. When the cells reached 80–90% confluence the medium was removed and the cells washed twice with DPBS, and then incubated in 3 ml serum-free DMEM with test cytokines and growth factor for the indicated time period.
NORTHERN BLOT ANALYSIS
Total cellular RNA was isolated and purified using the RNeasy kit (Qiagen, Crawley, UK). Equal amounts (10–20 μg) of RNA were fractionated on 1% agarose, 0.4 M formaldehyde gels and stained with ethidium bromide to check for integrity. RNA was transferred to a GeneScreen Plus membrane, prehybridised for two to three hours in 50% formamide, 1 M NaCl, 1 × Denhardt's solution, 1% sodium dodecyl sulphate (SDS) and 100 μg/ml denatured salmon sperm DNA. Blots were probed for 18 hours at 42°C with full length human MMP-1, MMP-3, MMP-8, (from A Galloway, British BioTechnology, Oxford, UK), MMP-13 (from Dr V Κnäuper, University of East Anglia, Norwich, UK), TIMP-1, and 1.3 kb rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH; from Dr C Redfern, University of Newcastle, UK) cDNA probes labelled by random priming with α32P dCTP. Blots were washed twice in 2 × SSC for 15 minutes at room temperature and twice for 30 minutes at 65°C in 2 × SSC, 0.5% SDS. 32P labelled cDNA-mRNA hybrids were visualised by autoradiography. The mRNA levels were measured by scanning densitometry of the bands using ImageMaster ID software (Amersham Pharmacia Biotech, Little Chalfont, UK). MMP-1, MMP-3, MMP-8, MMP-13, and TIMP-1 mRNA levels were normalised to GAPDH in the same RNA samples.31
STATISTICS
Student's t test was used to compare each group with control.
Results
EFFECTS OF IGF1 ON IL1α OR IL1α + OSM STIMULATED PG RELEASE
PG is readily released from cartilage when stimulated with a variety of cytokines and other agents.19-22 ,28 ,37 In this study we measured the release of PG at day 2 and confirmed the results of previous studies showing that IGF1 at 100 ng/ml can block the PG release induced by IL1α and IL1α + OSM by approximately 20–35%, although increasing IGF1 to 1000 ng/ml caused no further decrease in the amount of PG released (data not shown).19 ,22
LACTATE DEHYDROGENASE (LDH) ASSAY
Normally, the viability of cartilage explants was assessed by screening for the production of LDH using the Cytotox 96 assay. Previously, the maximum concentrations of IGF1 used in cartilage explant cultures were in the range 500–700 ng/ml.23 ,36In this experiment the maximum concentration of IGF1 used was 1000 ng/ml. An LDH assay was performed with cultured bovine nasal cartilage media, controls, or treated with IL1α or IL1α + OSM +/− IGF1 (1000 ng/ml). No increase in LDH levels with any of the cytokine combinations was found (data not shown).
EFFECT OF IGF1 ON IL1α OR IL1α + OSM INDUCED COLLAGEN RELEASE
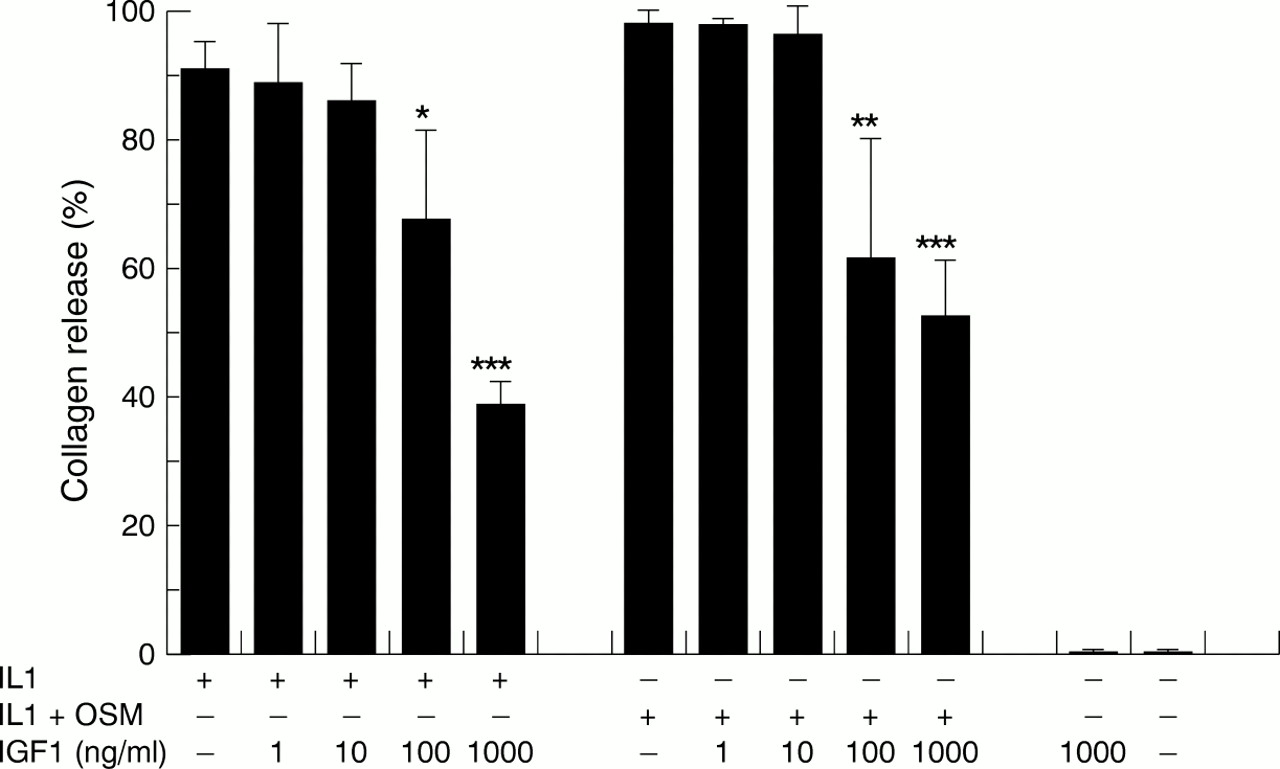
Previous studies have shown that OSM and IL1α together markedly stimulated the release of collagen fragments from bovine nasal cartilage in culture by day 14.28 ,29 In this study we used this combination of cytokines to stimulate, rapidly and reproducibly, the release of collagen from cartilage. To test the ability of IGF1 to inhibit IL1α or IL1α + OSM stimulated collagen release, bovine cartilage was cultured in the presence of IL1α or IL1α + OSM with various concentrations of IGF1 for 14 days. By day 14, over 90% collagen release was seen in cytokine treated cartilage samples; IGF1 produced a dose dependent inhibition of the IL1α and IL1α + OSM stimulated collagen release (fig 1). A statistically significant inhibition of IL1α or IL1α + OSM induced cartilage collagen release was seen at 100 ng/ml IGF1. The IL1α or IL1α + OSM induced collagen release was blocked by approximately 45%.
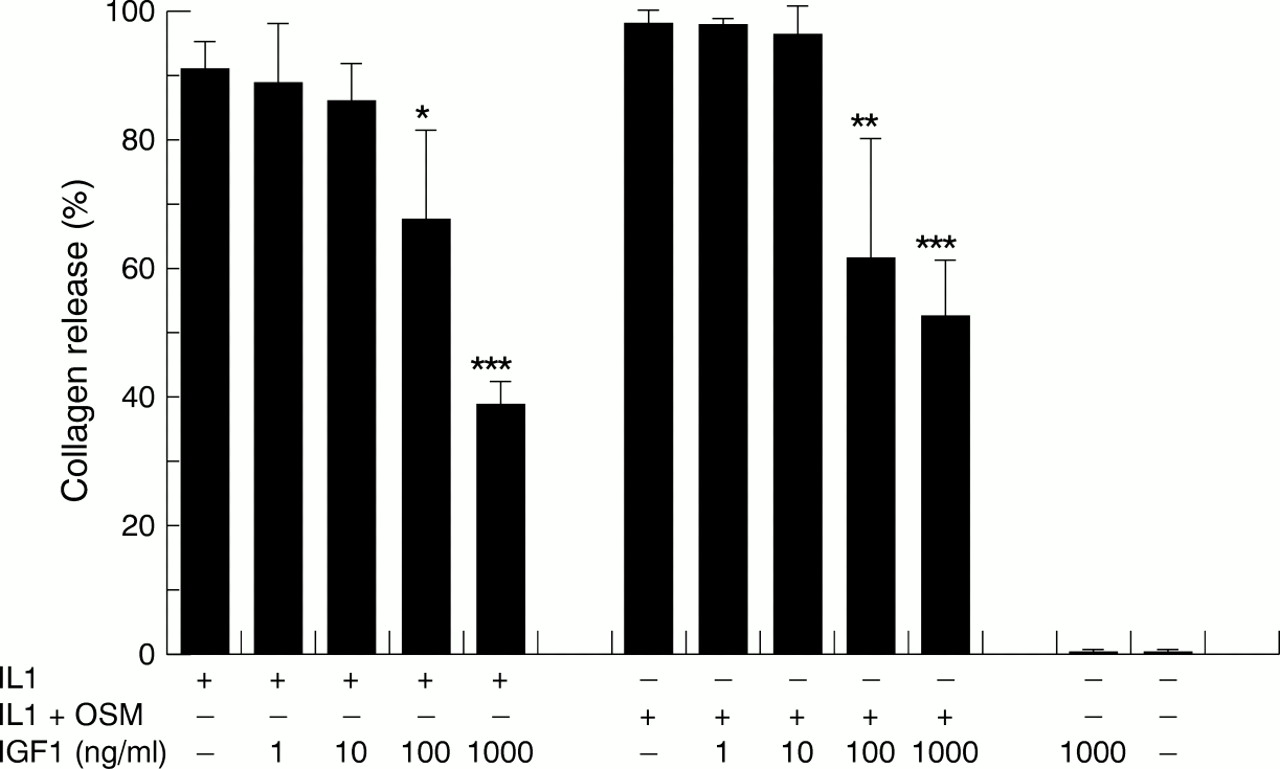
The effects of insulin-like growth factor 1 (IGF1) on the release of collagen fragments induced by interleukin 1α (IL1α) and IL1α + oncostatin M (OSM). Bovine nasal cartilage was cultured in serum-free medium in the presence of either medium alone, or medium containing IL1α (1 ng/ml), IL1α (0.2 ng/ml) + OSM (2 ng/ml) with IGF1 (1–1000 ng/ml) for 14 days. Media were collected at days 7 and 14. The levels of collagen fragments released into the media on days 7 and 14 were determined by the measurement of hydroxyproline as described in “Material and methods”. Results shown are for the cumulative collagen release for 14 days of culture and expressed as a percentage of the total (mean (SD)). All assays were done in quadruplicate. The experiments were performed in duplicate. Significance was analysed with respect to the cytokine compared with the cytokine + IGF1, or IGF1 compared with control. *p<0.05, **p<0.01, ***p<0.001 using t test.
EFFECT OF IGF1 ON LEVELS OF ACTIVE AND PROCOLLAGENASE IN THE MEDIA OF IL1α OR IL1α + OSM TREATED CARTILAGE
The media collected from IL1α or IL1α + OSM with or without IGF1 treated cartilage samples were assayed for the activity of collagenolytic enzymes as determined by the diffuse fibril assay using3H acetylated collagen.36 APMA was added to activate procollagenases. IL1α or IL1α + OSM treated cartilage samples produced high levels of active and procollagenase at day 14. IGF1 significantly reduced both active and procollagenase activity induced by IL1α or IL1α + OSM, in a dose dependent way (fig 2). The pattern of down regulation of collagenase activity correlated with the reduction in collagen release (compare figs 1 and 2).

Effects of insulin-like growth factor 1 (IGF1) on the levels of collagenolytic activity induced by interleukin 1α (IL1α) or IL1α + oncostatin M (OSM). Bovine nasal cartilage was treated as described for fig 1. The levels of procollagenase and active collagenase activity in media removed from cultured cartilage at day 14 after stimulation with (A) IL1α and (B) IL1α + OSM, with or without IGF1, were measured as described in “Material and methods”.
EFFECT OF IGF1 ON TIMP INHIBITORY ACTIVITY IN THE MEDIA OF IL1α OR IL1α + OSM TREATED CARTILAGE
Cytokines such as TGFβ1 and OSM induce TIMP-1 synthesis and secretion in human chondrocytes.38 ,39 We decided to investigate the effect of IGF1 on TIMP-1 production, which is down regulated by IL1α or IL1α + OSM. The media collected from cartilage treated with IL1α or IL1α + OSM in the presence of various concentrations of IGF1 were assayed for TIMP activity. These experiments showed no free TIMP activity in any of the media samples (data not shown). Furthermore, IGF1 alone did not alter the measurable level of TIMP activity compared with control (data not shown).
EFFECTS OF IGF1 ON THE EXPRESSION OF MMP AND TIMP mRNA IN BOVINE NASAL CHONDROCYTES
Bovine nasal chondrocytes were treated with IL1α, OSM, and IL1α + OSM with 100 ng/ml IGF1 in serum-free medium. After 24, 48, and 72 hours, total cellular RNA was purified and transferred to a GeneScreen membrane. Blots were probed with cDNA probes for MMP-1, MMP-3, MMP-8, MMP-13, and TIMP-1. The blots showed that IL1α stimulated MMP-1, MMP-3, and MMP-13 mRNA expression in a time dependent fashion (fig 3). IGF1 can decrease IL1α, OSM, and IL1α + OSM induced MMP-1, MMP-3, and MMP-13 mRNA expression, and also down regulate the basal MMP expression. There was no detectable MMP-8 mRNA expression in bovine nasal chondrocytes after stimulation with IL1α or IL1α + OSM, with or without IGF1 (data not shown). This may be because the human MMP-8 probe does not cross react with bovine MMP-8 mRNA or bovine chondrocytes express MMP-8 mRNA at levels below northern blot detection. IL1α + OSM and OSM up regulated TIMP-1 expression. IGF1 had no significant effect on the expression of TIMP-1 mRNA when added alone or in combination with cytokines (fig3).

Effects of insulin-like growth factor 1 (IGF1) on the expression of metalloproteinase (MMP) and tissue inhibitor of metalloproteinase (TIMP) mRNA in bovine chondrocytes. Bovine nasal chondrocytes were incubated with interleukin 1α (IL1α; 1 ng/ml), oncostatin M (OSM; 10 ng/ml) and IL1α (0.2 ng/ml) + OSM (2 ng/ml) with IGF1 (200 ng/ml) in serum free medium for the times indicated. The expression of MMP-1, MMP-3, MMP-8, MMP-13, and TIMP-1 mRNA was analysed by northern blot. Lane 1 = IL1α ; lane 2 = IL1α + IGF1; lane 3 = IL1α + OSM; lane 4 = IL1α + OSM + IGF1; lane 5 = OSM; lane 6 = OSM + IGF1; lane 7 = IGF1; lane 8 = control.
EFFECTS OF IGF1 ON CYTOKINE INDUCED MMP-1, MMP-3, MMP-8, AND MMP-13 mRNA EXPRESSION IN HUMAN ARTICULAR CHONDROCYTES
Human articular chondrocytes were treated with IL1α, OSM, and IL1α + OSM with or without IGF1 (200 ng/ml) in serum-free medium for 48 hours. Total cellular RNA was purified and transferred to a GeneScreen membrane. Blots were probed with cDNA probes for MMP-1, MMP-3, MMP-8, and MMP-13. The blots showed that IGF1 can decrease IL1α, OSM, and IL1α + OSM induced expression of MMP-1, MMP-3, MMP-8, and MMP-13 mRNA (fig 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of insulin-like growth factor 1 (IGF1) on the expression of metalloproteinase (MMP) mRNA in human articular chondrocytes. Human articular chondrocytes were incubated with interleukin 1α (IL1α; 1 ng/ml), oncostatin M (OSM; 10 ng/ml), and IL1α (1 ng/ml) + OSM (10 ng/ml) with IGF1 (200 ng/ml) in serum-free medium for 24 hours. The expression of MMP-1, MMP-3, MMP-8, and MMP-13 mRNA was analysed by northern blot.
Discussion
The major focus of this study was to investigate agents that prevent collagen breakdown. We demonstrate, for the first time, that IGF1 can partially block collagen fragment release and decrease collagenase activity induced by IL1α and IL1α + OSM from bovine nasal cartilage. Previously, IGF1 has been shown to enhance collagen and PG synthesis by chondrocytes and osteoblasts, and to inhibit both basal and IL1 induced degradation of PG in cartilage.16-22 Furthermore, IGF1 can successfully maintain cartilage structure and biochemistry and enhance the repair of damaged cartilage.23 ,33 ,40-42 No studies on IGF1 blocking collagen breakdown induced by cytokines in cartilage have been reported previously. In this study we show that there is a similar pattern attenuating IL1α and IL1α + OSM induced collagen release and a down regulation of active and procollagenases by IGF1 in cultured bovine cartilage. This suggests that down regulation of collagenases by IGF1 is an important mechanism in blocking the action of proinflammatory cytokines which play a part in cartilage resorption, especially collagen degradation. The protection of cartilage by IGF1 was considered to involve the prevention of production and release of collagenases or the increase in a protease activator inhibitor,25 ,26 ,43 but these studies are not conclusive.
MMPs are known to degrade various components of the cartilage matrix and several have been demonstrated in RA and OA tissues.11 ,12 ,44 To date, most of the studies on MMP expression and production in arthritic joint tissues and cells have been observations on the effect of cytokine or growth factor alone on chondrocytes.3 ,45 Only a few studies have examined the interactions between catabolic cytokines and anabolic growth factors on the expression of MMPs in chondrocytes. TGFβ1 is known to inhibit partially IL1 stimulated cartilage degradation in vitro46and was shown to down regulate MMP mRNA levels in osteoarthritic and normal chondrocytes.47 Interferon γ can also block the production of IL1 stimulated collagenases in human articular chondrocytes.48 Our observations, using primary human and bovine chondrocytes, show that IGF1 can down regulate MMP-1, MMP-3, and MMP-13 mRNA expression induced by IL1α and IL1α + OSM. Furthermore, we found that IL1α and IL1α + OSM could stimulate the expression of MMP-8 in human articular chondrocytes, though the level of MMP-8 expression was relatively low compared with the expression of other MMPs. MMP-8 levels were also down regulated by IGF1 in human chondrocytes. This is consistent with the observations of Cole and coworkers when they first detected that MMP-8 mRNA was expressed in human chondrocytes. They found that although the amount of mRNA expression by either isolated human articular chondrocytes or chondrocytes within cartilage was very low, it was significantly up regulated by IL1.11 In bovine nasal chondrocytes there was no detectable expression of MMP-8 mRNA after stimulation by cytokines. This might be because the human MMP-8 probe failed to cross react with bovine MMP-8, or perhaps the level of MMP-8 mRNA expression by bovine nasal chondrocytes was below the limit of detection. IGF1 has been previously shown to inhibit the expression of collagenase mRNAs, such as MMP-1 and MMP-13, in osteoblasts.25 ,26 This study is the first report that IGF1 can down regulate the expression and production of collagenases in chondrocytes stimulated by proinflammatory cytokines.
OSM is a multifunctional cytokine produced by activated T lymphocytes and monocytes,49 as well as rheumatoid synovial macrophages.29 It is structurally and functionally related to the IL6 family of cytokines, which includes IL6, IL11, leukaemia inhibitory factor, OSM, ciliary neurotrophic factor, and cardiotrophin 1. The IL6 family of cytokines, except cardiotrophin 1, can inhibit PG synthesis in articular cartilage.37 OSM was initially proposed as a chondroprotective agent when it was found to up regulate TIMP-1 production in human articular chondrocytes, synovial fibroblasts, and synovial lining cells.39 Later studies have shown that it stimulates PG release and also inhibits PG synthesis in cartilage explants.37 A raised level of OSM in rheumatoid joints correlates significantly with synovial fluid white cell count.29 ,50 More recently, studies in our laboratory have shown that the combination of IL1α with OSM initiated a dramatic release of collagen from bovine, porcine, and human cartilage which correlated with increased levels of active collagenase and procollagenase and decreased levels of TIMP activity.29 In this study we combined OSM (2 ng/ml) with IL1α (0.2 ng/ml), and after 14 days' incubation over 90% collagen was released from the cartilage. At these concentrations, neither agent alone promoted collagen release (data not shown). The potency of the low concentration combination of IL1α + OSM for collagen release is similar to that with 1 ng/ml IL1α alone. Furthermore, there are similar levels of collagenase in the media from IL1α + OSM (0.2/2 ng/ml) stimulated cartilage as from IL1α (1 ng/ml) alone at day 14. OSM alone, even at 10 ng/ml in cultured bovine cartilage, did not significantly induce collagen release. Northern blot data indicated that OSM (at 10 ng/ml) up regulated the expression of MMP mRNAs, but the potency was lower than that of IL1α or IL1α + OSM. IGF1 also down regulated this expression. This agrees with the finding that when SW 1353 chondrosarcoma cells were treated with OSM alone very low MMP-1 and MMP-13 levels were produced. However, the release of the procollagenases MMP-1 and MMP-13 was potentiated with OSM in combination with IL1.51 Clearly, the increased release of collagen from bovine nasal cartilage is due to increasing levels of active collagenases, but the precise mechanisms involved in the synergy are not yet known. The mechanism may be associated with OSM up regulating enzymes that are responsible for the initiation of activation cascades, and/or the stimulation of other catabolic cytokines.
We noted that IL1α + OSM or OSM up regulated TIMP-1 mRNA expression, and that IGF1 alone or in combination with IL1α or IL1α + OSM did not significantly modify TIMP-1 production in chondrocytes. This is consistent with the recent report by Canalis et al, where it was found that neither IGF1 nor IGF-2 modified the expression of TIMP-1, TIMP-2, or TIMP-3 mRNA in osteoblasts.25
It is known that IGF1 bioavailability is modulated by specific IGFBPs, six of which have been identified (IGFBP1–6). IGFBPs can bind to this growth factor in order to sequester it so as to prevent its binding to IGF receptors19 and IGFBP was found to inhibit IGF1 stimulated PG synthesis.52 Raised levels of IGFBP have been detected in RA and OA synovial fluids, and the production of IGFBPs by chondrocytes is increased in these disease conditions. IL1 induces secretion of IGFBPs and down regulates IGF mRNA levels in chondrocytes, which indicates that IGFBPs might accelerate cartilage destruction by blocking the protective action of IGF1.19Studies also found that IGFBPs can be degraded by MMP-1, MMP-2, and MMP-353 ,54 and it is considered that degradation of IGFBPs by MMPs is an important step in regulating IGF bioactivity. This study clearly shows that IGF1 has a protective effect on cartilage stimulated to resorb, but previous studies have shown that human OA chondrocytes do not always respond to IGF, possibly because increased levels of IGFBP are present.55 Schoutenet al showed that increased circulating levels of IGF1 were associated with osteophyte formation but not with preventing the erosion of cartilage in patients with OA of the knee.56 Thus the interactions between IGF, IGFBPs, and proteinases are complex in both health and disease and need further study.
IGF1 can block collagen release and decrease collagenase activity induced by IL1α and IL1α + OSM in bovine nasal cartilage. This is further confirmed from collagenase mRNA levels by northern blot analyses. These observations represent new biological effects for this growth factor and suggests that down regulation of collagenases by IGF1 may be an important mechanism in preventing cartilage resorption initiated by proinflammatory cytokines.
Acknowledgments
This work was supported by the Arthritis Research Campaign and the Newcastle University Hospitals Special Trustees, UK.