Article Text

Abstract
OBJECTIVE To determine the prevalence of myositis specificautoantibodies (MSAs) and several myositisassociated autoantibodies (MAAs) in a large group of patients with myositis.
METHODS A total of 417 patients with myositis from 11 European countries (198 patients with polymyositis (PM), 181 with dermatomyositis (DM), and 38 with inclusion body myositis (IBM)) were serologically analysed by immunoblot, enzyme linked immunosorbent assay (ELISA) and/or immunoprecipitation.
RESULTS Autoantibodies were found in 232 sera (56%), including 157 samples (38%) which contained MSAs. The most commonly detected MSA was anti-Jo-1 (18%). Other anti-synthetase, anti-Mi-2, and anti-SRP autoantibodies were found in 3%, 14%, and 5% of the sera, respectively. A relatively high number of anti-Mi-2 positive PM sera were found (9% of PM sera). The most commonly detected MAA was anti-Ro52 (25%). Anti-PM/Scl-100, anti-PM/Scl-75, anti-Mas, anti-Ro60, anti-La, and anti-U1 snRNP autoantibodies were present in 6%, 3%, 2%, 4%, 5%, and 6% of the sera, respectively. Remarkable associations were noticed between anti-Ro52 and anti-Jo-1 autoantibodies and, in a few sera, also between anti-Jo-1 and anti-SRP or anti-Mi-2 autoantibodies.
CONCLUSIONS The incidence of most of the tested autoantibody activities in this large group of European patients is in agreement with similar studies of Japanese and American patients. The relatively high number of PM sera with anti-Mi-2 reactivity may be explained by the use of multiple recombinant fragments spanning the complete antigen. Furthermore, our data show that some sera may contain more than one type of MSA and confirm the strong association of anti-Ro52 with anti-Jo-1 reactivity.
- autoimmunity
- polymyositis
- dermatomyositis
- autoantibodies
- autoantigens
Statistics from Altmetric.com
The idiopathic inflammatory myopathies (IIM) are a heterogeneous group of systemic diseases characterised by a progressive proximal muscle weakness, raised serum creatine kinase, characteristic electromyographic abnormalities, and inflammatory infiltrates in skeletal muscles.1 The three major categories of these disorders are polymyositis (PM), dermatomyositis (DM), and inclusion body myositis (IBM).1 The initial events leading to the onset of these diseases are still unknown. Both PM and DM are commonly regarded as autoimmune disorders. The immunopathogenesis of IBM is unclear as evidence has been presented for a similar cell mediated autoimmune response in PM and IBM, whereas others suggested that the immune response in IBM is secondary to degenerative changes in skeletal muscle tissue (reviewed by Askanas and Engel2).
Various autoantibodies directed to defined nuclear and cytoplasmic antigens are found in up to 55% of patients with PM or DM.3-5 Most of these autoantibodies, referred to as myositis associated autoantibodies (MAAs), are not specific for IIM, because they are commonly encountered in other rheumatic disorders without clinical signs of myositis. Antigens recognised by MAAs include among others, the PM/Scl autoantigens, the Mas autoantigen, and components of the U1 small nuclear RNP (snRNP) or the cytoplasmic Ro RNPs such as Ro60/SSA, La/SSB, and Ro52.6 Anti-PM/Scl autoantibodies recognise two components, PM/Scl-100 and PM/Scl-75, of an RNA processing complex called the exosome.7 The Mas autoantigen has not been identified yet, although it is known to be a tRNASel-binding protein of about 48 kDa molecular weight, which plays a part in the pathway of selenocysteine incorporation.8 ,9 The U1 snRNP is involved in pre-messenger RNA splicing.10 The function of the Ro RNPs remains to be established.11
Some patients have autoantibodies which are specific for IIM. These myositis specific autoantibodies (MSAs) target a subset of aminoacyl-tRNA synthetases, transfer RNAs, components of the signal recognition particle (SRP), and the nuclear helicase/ATPase Mi-2.6 ,12 ,13 Anti-synthetase autoantibodies are directed towards a subset of aminoacyl-tRNA synthetases.13-15 These enzymes are critical components of the translation machinery because each enzyme catalyses the esterification of one amino acid to its cognate tRNA. The most commonly detected and best characterised MSA is the anti-Jo-1 antibody (anti-histidyl-tRNA synthetase), which is found in up to 25% of the myositis sera.5 ,13 ,14 Autoantibodies directed toward five other synthetases (specific for alanine (anti-PL12), glycine (anti-EJ), isoleucine (anti-OJ), threonine (anti-PL7), and asparagine (anti-KS)) have been reported to occur much less commonly.5 ,13-15 Also, naked tRNA may be recognised by autoantibodies in about a third of the anti-Jo-1 sera and in most anti-PL12 sera.16 ,17
The SRP complex consists of an 7SL RNA and six proteins of 9 kDa, 14 kDa, 19 kDa, 54 kDa, 68 kDa, and 72 kDa.18 This particle functions in the co-translational translocation of secretory and membrane proteins to the membrane of the endoplasmic reticulum.18 Anti-SRP autoantibodies have been detected in about 4% of patients with myositis who show a distinct seasonal onset of disease, no association has been found with other MSAs, and almost all patients had PM rather than DM.5 ,19 ,20
Two autoantigenic forms of the Mi-2 antigen, Mi-2α and Mi-2β, have been reported.12 ,21 ,22 The Mi-2β antigen is part of a protein complex which exhibits several chromatin remodelling functions to regulate transcription.23 ,24 Autoantibodies recognising the Mi-2 antigen are detected in about 20% of the myositis sera, and reported to occur primarily in patients with DM.12
Analysis of autoantibody profiles has been shown to be important in the diagnosis of some autoimmune diseases, such as anti-dsDNA and anti-Sm autoantibodies in systemic lupus erythematosus (SLE)25 and anti-cyclic citrullinated protein in rheumatoid arthritis.26 A classification system based on the MSA profiles has been proposed as a useful alternative for classification into clinical groups.5 The most important clinical association is referred to as the “anti-synthetase syndrome” which is characterised by the presence of autoantibodies directed toward a subset of aminoacyl-tRNA synthetases and a clinical syndrome consisting of myositis, arthritis, Raynaud's phenomenon, interstitial lung disease, and other clinical features.4 ,5 Other remarkable clinical associations are the high incidence of anti-Mi-2 autoantibodies in sera of patients with DM, the exclusion of IBM when anti-Jo-1 autoantibodies are present,27 and more severe myositis which is often resistant to treatment when anti-SRP autoantibodies are present.5 ,6
Over the past two decades a number of studies analysing the autoantibody profiles in small cohorts of sera from patients with myositis have been published.28-36 However, studies describing the more or less complete serological analysis of large numbers of myositis sera (n>200) are limited.5 ,37 Here, we present a serological study analysing the MSA and MAA antibody profiles of more than 400 European patients with myositis.
Patients and methods
PATIENTS
Serum samples were collected from 417 patients with IIM from several European rheumatological and neurological institutes located in the Czech Republic (n=72), Finland (n=30), Germany (n=8), Greece (n=56), Iceland (n=13), Italy (n=16), the Netherlands (n=122),16 ,27 ,38 ,39 Slovenia (n=27), Sweden (n=37), Switzerland (n=6), and the United Kingdom (n=30). Patients were classified as PM, DM, or IBM using recently published criteria.1 ,40 When anti-Mi-2 autoantibodies were found, patients were also classified according to the established criteria published by Bohan and Peter,41 which, however, did not change the diagnosis. The patient group studied comprised 198 patients with PM, 181 with DM, and 38 with IBM. Standardised questionnaires containing clinical data on signs and symptoms, course of the disease, response to treatment, results of laboratory investigations, electromyographic findings, and muscle biopsy specimens were provided by the treating doctors. Incomplete questionnaires and questionnaires that contained insufficient information for an accurate diagnosis were excluded from the study.
A pool of 10 human sera from subjects without any known history of autoimmune disease was used as a control in all assays performed.
SEROLOGICAL ANALYSIS OF MYOSITIS SERA
All autoantibody specificities were analysed in the Nijmegen laboratory, except for the anti-Mi-2 and anti-PM/Scl enzyme linked immunosorbent assays (ELISAs), which were performed in Professor Dr HP Seelig's laboratory (Karlsruhe, Germany).
ANTI-SYNTHETASE AUTOANTIBODIES
Anti-synthetase autoantibodies were detected by immunoprecipitation (IP) using a HeLa cell S100 extract as described previously.16 Sera which precipitated RNA in the tRNA range as determined by denaturing polyacrylamide gel electrophoresis were designated anti-synthetase positive. Anti-Jo-1 autoantibodies were distinguished from “other anti-synthetase autoantibodies” by performing additional immunoblotting and ELISA analyses as described previously.38 All anti-Jo-1 sera were positive in both IP and ELISA or immunoblot. Presence of anti-Jo-1 activity was also determined by the dotblot analysis described below. In addition, anti-synthetase positive sera were analysed for the presence of anti-tRNA autoantibodies by IP using total RNA from a deproteinised HeLa cell S100 extract as described previously.16
ANTI-MI-2 AUTOANTIBODIES
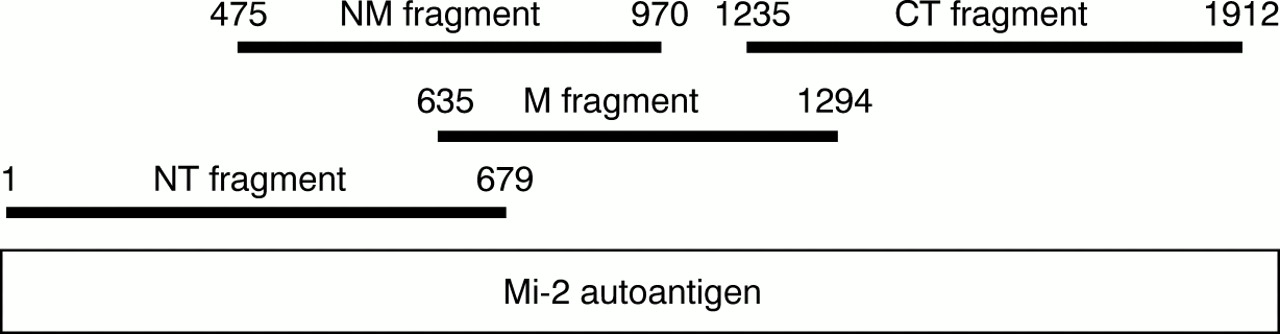
The presence of anti-Mi-2 autoantibodies was determined by ELISA. Four overlapping fragments depicted in fig 1 (NT = N-terminal, M = middle, NM = N-terminal/middle, and CT = C-terminal) spanning the complete amino acid sequence of the Mi-2β autoantigen were used. The cDNAs of NT, M, and CT fragments were synthesised essentially as described for fragment NM12 and ligated into an in house modified pEX34 expression vector containing only 10 amino acids of the vectors MS2 polymerase fusion protein. Proteins were expressed in transformed E coli 537 cells and purified by Ni2+ affinity chromatography followed by preparative sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE). With 0.2 μg/well of each of the proteins, medium binding microtitre plates (Greiner, Nürtingen, Germany) were coated as described.12Antibody screening was performed with 200 μl of diluted sera (1:100 in 20 mM Tris-HCl pH 7.4, 150 mM NaCl, 0.05% NaN3, 0.05% Tween-20, 2.5% bovine serum). After incubation (30 minutes, 37°C) and washing (three times with phosphate buffered saline, 0.01% Tween-20) plates were incubated (30 minutes, 37°C) with 200 μl/well of alkaline phosphatase conjugated AffiniPure Fcγ fragment specific goat antihuman IgG (Jackson Laboratories, West Baltimore Pike, USA) diluted 1:2000 in 20 mM Tris-HCl pH 7.4, 150 mM NaCl, 0.5% bovine serum albumin, 0.05% Tween-20, 1 mM MgCl2, 0.1 mM Zn acetate, 0.05% NaN3. Bound antibodies were visualised with 38 mM disodium p-nitrophenylphosphate, 50 mM diethanolamine pH 9.8, 1 mM MgCl2, 0.05% NaN3. The enzyme reaction was stopped by adding 50 μl 3 N NaOH. Optical densities were measured bichromatically at 492 nm/620 nm (Titertek Multiscan MCC340 Mkll; Flow, Meckenheim, Germany). Tests were regarded as positive at ⩾2.5-fold optical densities compared with that of three negative control sera. Positive and negative controls as well as patients' sera were run in duplicate on every test plate. The anti-Mi-2 activity detected by ELISA was also analysed by western blotting using the purified recombinant Mi-2β fragments shown in fig1.

Schematic representation of the Mi-2β autoantigen and the fragments used in ELISA. NT = N-terminal; M = middle; NM = N-terminal/middle; CT = C-terminal.
ANTI-PM/SCL AUTOANTIBODIES
Reactivity with PM/Scl-100 and PM/Scl-75 was determined by ELISA. For synthesis of the PM/Scl proteins cDNAs from HepG2 cell RNA (ATCC HB 8065) were constructed according to their sequence data (PM/Scl-75: Acc. No M58460; PM/Scl-100: Acc. No X66113) and processed essentially as described above for Mi-2 protein synthesis and purification. Each protein (0.4 μg/well of PM/Scl-75 and 0.2 μg/well of PM/Scl-100) was coated onto microtitre plates. Coating, blocking, and assay performance followed the Mi-2 autoantibody protocols.
ANTI-RO60, ANTI-RO52, AND ANTI-LA AUTOANTIBODIES
Expression and purification of the Ro60, Ro52, and La autoantigens and corresponding ELISAs were performed as described previously.38
DETECTION OF ANTI-MAS, ANTI-SRP, ANTI-JO-1, AND ANTI-U1 SNRNP AUTOANTIBODIES BY DOTBLOT ANALYSIS
Immunoprecipitations were performed as previously described.16 Immunoprecipitated RNA recovered after phenol/chloroform extraction was spotted onto Hybond N+ membranes (Amersham, United Kingdom). After drying and ultraviolet cross linking, the dotblots were hybridised with various antisense RNA probes in (pre)hybridisation mix (6 × saline sodium citrate (SSC), 5 × Denhardt's solution, 0.1% SDS, 100 μg/ml sheared, denatured herring sperm DNA). After overnight incubation at 65°C, membranes were washed three times with 2 × SSC/0.1% SDS (antisense tRNASel and antisense tRNAHis) or with 0.1 × SSC/0.1% SDS (antisense 7SL RNA) at 65°C and exposed to Kodak X-Omat AR imaging film (Eastman Kodak, Rochester, NY). Antisense RNA probes were transcribed in vitro in the presence of [32P]UTP from linearised templates. To determine anti-Jo-1 reactivity, antisense tRNAHis was transcribed by SP6 RNA polymerase using aHindIII linearised template of tRNAHis cDNA/pSP65p. To detect anti-Mas reactivity, a construct containing the tRNASel cDNA, kindly provided by Dr A Krol (CNRS, Strasbourg, France), was used. After recloning into pSP65 (BamHI/EcoRI), antisense tRNASel probe was transcribed in vitro fromBamHI linearised template by SP6 RNA polymerase. To detect anti-SRP reactivity, a 7SL cDNA/pSP64 construct, kindly provided by Dr K Strub (University of Geneva, Switzerland), was used. Antisense 7SL RNA probe was transcribed in vitro from theEcoRI linearised template by SP6 RNA polymerase. Anti-U1 snRNP reactivity was detected by the antisense U1 RNA probe, which was produced by in vitro transcription using anEcoRI linearised U1 cDNA/pGEM7-ZF(+) construct.
Results
In this study sera from 417 European patients with myositis were analysed for the presence of major MSAs and MAAs using various established methods as well as a newly described dotblot assay which was used to determine the presence of several anti-ribonucleoprotein complex autoantibodies. Most patients had either PM (n=198 (48%)) or DM (n=181 (43%)), while IBM was diagnosed in 38 patients (9%).
Table 1 provides an overview of the results. Autoantibodies (MAAs and/or MSAs) were detected in 232 sera (56%), including 122 PM (62% of PM sera), 98 DM (54% of DM sera), and 12 IBM sera (32% of IBM sera). MSAs were detected in 157 sera (38%), including 76 PM (38% of PM sera), 74 DM (41% of DM sera), and seven IBM sera (18% of IBM sera).
Prevalence of myositis specific autoantibodies and myositis associated autoantibodies in serum samples from European patients with myositis
ANALYSIS OF MSAS
Anti-synthetase autoantibodies
Only two groups of anti-synthetase autoantibodies were discriminated: anti-Jo-1 positive and “other anti-synthetase” positive sera. Owing to the low incidence of autoantibodies directed towards other synthetases and the fact that a detailed specification of the anti-synthetase antibody is clinically of minor importance, the precise nature of the antigen recognised by “other tRNA” precipitating sera was not determined.
As shown in table 1, anti-Jo-1 was found in 73 (18%) of the myositis sera, which were diagnosed mostly as PM (43 (22%) of the PM sera) or DM (28 (16%) of the DM sera). Only a few IBM sera contained anti-Jo-1 reactivity (2 (5%) of the IBM sera). A similar tendency was observed for “other anti-synthetase” reactivities, though the total number of positive sera was much lower (12 (3%)). Again, mostly PM sera (six) and DM sera (five) immunoprecipitated tRNA (other than tRNAHis), while only one IBM serum sample contained autoantibodies directed to a non-Jo-1 synthetase.
Some myositis sera contain anti-tRNA autoantibodies next to the cognate anti-synthetase autoantibodies.16 ,17 Twenty six anti-Jo-1 positive sera (36% of Jo-1 positive sera) appeared to contain anti-tRNAHis activity. Anti-tRNA activity was also found in two of the 12 sera which contained an anti-synthetase activity that was not analysed in detail.
Anti-SRP autoantibodies
Anti-SRP positive sera are known to co-immunoprecipitate the 7SL RNA component of the particle.42 Therefore, all sera were screened by IP followed by a dotblot assay rather than by ELISA or immunoblotting using purified recombinant proteins. In the dotblot assay, RNAs recovered after IP were hybridised to an antisense 7SL RNA probe. As shown in fig 2, the antisense 7SL RNA probe hybridised specifically to RNA precipitated by the anti-SRP reference serum and not to RNA precipitates of control sera. When a number of SRP positive sera were used the specificity of the dotblot assay was confirmed by IP of 7SL RNA from a metabolically 32P labelled HeLa cell extract (data not shown).

{kind=link}
{kind=link}
Analysis of reference myositis sera and a pool of 10 normal human sera (NHS) by dotblot. RNA precipitated by autoantibodies in myositis sera was spotted on nylon filters and probed with antisense RNA probes to detect anti-Mas, anti-SRP, and anti-Jo-1 reactivity.
Because the 7SL RNA is an RNA polymerase III transcript and the La autoantigen is known to bind the pre-RNA polymerase III transcripts, the possibility existed that false positives were included. However, IPs performed with 32P labelled HeLa cell extract and anti-La positive sera indicated that the amount of 32P labelled pre-7SL RNA precipitated by anti-La positive/anti-SRP negative sera is rather low and clearly distinguishable from the signals obtained with anti-SRP sera (data not shown).
As shown in table 1, 20 myositis sera (5%) were anti-SRP positive, including 14 PM sera (7%), five DM sera (3%), and one IBM serum (3%). None of the SRP positive sera were anti-La positive (see below, table 4).
Associations of myositis specific autoantibodies and myositis associated autoantibodies in serum samples from European patients with myositis
Anti-Mi-2 autoantibodies
Detection of anti-Mi-2 autoantibodies has previously been performed by ELISA using a recombinant internal fragment of the Mi-2β antigen.12 Although this NM fragment (formerly known as the MB fragment) appears to contain the major epitope region, some positive sera may not be detected using only this fragment. Therefore, in this study, four overlapping fragments spanning the complete amino acid sequence of the Mi-2β antigen were used in ELISA, including the NM fragment (fig 1). As shown in table 1, 58 myositis sera (14%) were reactive with one or more fragments. As expected, the majority of the Mi-2 positive patients had DM (38 (21%)). However, also, a substantial number of patients were diagnosed as PM (17 (9%)) and three patients had IBM (8%). We did not find consistently lower ELISA titres for PM or IBM sera compared with DM sera, as has been reported by Rouxet al.43 The anti-Mi-2 reactivity of most of the PM, DM, and IBM sera could be confirmed by western blotting using the purified recombinant Mi-2β fragments shown in fig 1 (data not shown).
Table 2 shows the reactivity of the Mi-2 positive sera with each of the four fragments. These results confirm that most Mi-2 positive sera (42 (72%)) recognise epitope(s) present in the NM fragment. Table 3 shows the reactivity of individual sera with the fragments. It is evident that the neighbouring sequences of the NM fragment also contribute to the antigenicity of the Mi-2 protein, because a substantial number of NM positive sera also recognise the NT or M fragments (30 (71%) of NM positive sera). Unexpectedly, 16 Mi-2 positive sera did not recognise the NM fragment, but exclusively the NT or CT fragment, or both. Seven sera recognised exclusively epitopes on the NT fragment, eight sera reacted only with the CT fragment, and one serum recognised epitopes present on both CT and NT fragments, but not on the NM or M fragments. Furthermore, these results indicate that while 29 sera recognised the M fragment (table 2), this reactivity does not increase the sensitivity of the assay. All of the M positive sera also reacted with the NM fragment. As shown in table 3, 18 sera were reactive with both NM and M fragments, 10 sera recognised both NT, NM, and M fragments, and one serum recognised all four fragments.
Reactivity of anti-Mi-2 positive sera with recombinant Mi-2 fragments
Detailed analysis of the reactivity of anti-Mi-2 positive sera with the recombinant fragments
Taking into account the fact that the NM and M fragments share epitopes, a difference in the anti-Mi-2 response may be noticed between PM and DM sera. Results shown in tables 2 and 3 suggest that Mi-2 positive PM sera recognise the NT, NM, and CT fragments more or less equally. In contrast, Mi-2 positive DM sera seem preferentially to recognise the NM fragment. However, since there is overlap between the fragments and the total number of Mi-2 positive sera in each diagnosis is rather small, no statistically meaningful conclusions can be drawn.
ANALYSIS OF MAAS
Anti-Mas autoantibodies
Anti-Mas autoantibodies have been detected in only a few patients with myositis or autoimmune chronic active hepatitis and may also be associated with alcoholic rhabdomyolysis.8
In this study anti-Mas autoantibodies were identified by detection of the co-precipitating RNA component in the dotblot assay. As shown in fig 2, the antisense tRNASel probe hybridised specifically with the precipitated RNA of the anti-Mas reference serum and not with the RNA precipitates of control sera which included anti-synthetase sera. In this way (table 1), eight sera (2%) were shown to contain anti-Mas antibody (3% of PM sera v 1% of DM sera). Anti-Mas reactivity was not detected in IBM sera.
Anti-PM/Scl autoantibodies
Anti-PM/Scl autoantibodies have been reported to occur in sera of patients with myositis (5–8%), myositis-systemic sclerosis overlap (24–50%), and systemic sclerosis (2–3%).3 ,44PM/Scl-100 and PM/Scl-75 are recognised by ∼98% and ∼63% of the PM/Scl positive sera, respectively.45-48
In our collection of sera from European patients with myositis, anti-PM/Scl-100 and anti-PM-Scl-75 autoantibodies were detected in 23 (6%) and 11 (3%) of the myositis sera, respectively. Both specificities were only detected in PM or DM sera without any significant association with either diagnosis (table 1). In agreement with previous reports, all of the PM/Scl-75 positive sera were also reactive with PM/Scl-100.48
Anti-La, anti-Ro60, and anti-Ro52 autoantibodies
Anti-Ro60 and anti-La autoantibodies are mainly associated with SLE or primary Sjögren's syndrome, but have been reported to occur in 5–15% of myositis sera as well.49 ,50 Anti-Ro52 autoantibodies may be present in ∼20% of myositis sera.38
Anti-Ro60, anti-Ro52, and anti-La autoantibodies were found in 17 (4%), 106 (25%), and 21( 5%) of the European myositis sera, respectively (table 1). None of these antigens was strongly associated with PM, DM, or IBM. However, anti-Ro52 autoantibodies appear to be associated with particular types of autoantibodies (see below).
Anti-U1 snRNP autoantibodies
Anti-U1 snRNP autoantibodies are most commonly detected in sera of myositis patients with overlap disease (SLE, mixed connective tissue disease, or systemic sclerosis), but have been reported to be present in 5–15% of the patients with myositis.5 ,37 Myositis appears to be generally milder in patients with anti-snRNP autoantibodies than in patients without them.32
In this study anti-U1 snRNP autoantibodies were found in 25 (6%) of the sera analysed. Most of the anti-U1 snRNP positive patients had PM (17 patients) rather than DM (seven) or IBM (one). In all cases the anti-U1 snRNP reactivity was confirmed by [32P]pCp labelling of the immunoprecipitated RNA or by ELISA using purified recombinant U1-A protein (data not shown)
ASSOCIATIONS BETWEEN MSAS AND MAAS
The incidence of combined MSA specificities found in the screened European group was low (table 4). Anti-synthetase and anti-Mi-2 autoantibodies were found in four sera (5% of anti-synthetase positive sera), including three PM sera and one DM serum. The DM serum was weakly reactive with the Jo-1 antigen and moderately reactive with the NT fragment of the Mi-2 antigen. The three PM sera were moderately reactive with the Jo-1 antigen and moderately to strongly reactive with the CT fragment alone (one serum) or in combination with the NT fragment (one serum, strongly) or with the NM fragment (one serum, moderately). Thus all four sera were anti-Jo-1 positive and reactive with either or both NT and CT fragments of the Mi-2 antigen, while one serum also recognised the NM fragment. In addition, anti-SRP autoantibodies co-occurred with anti-synthetase autoantibodies in two sera, including one anti-Jo-1 positive PM serum and one “other anti-synthetase” positive DM serum.
Associations between MSAs and MAAs seem to occur more frequently.37 ,38 ,51 The most apparent correlation was found between anti-Ro52 and several MSAs (table 4). In this study anti-Ro52 autoantibodies were present in 56 out of 85 anti-synthetase positive sera (66%). This association is more clear for anti-Jo-1 positive sera (52 (71%) of Jo-1 positive sera) than for “other synthetase” positive sera (4 (33%)). In addition, anti-Ro52 antibody was present in combination with anti-SRP antibodies (6 (30%)) and anti-PM/Scl antibodies (3 (13%)). A number of anti-Mi-2 positive sera (both PM and DM) were found to contain anti-Ro52 autoantibodies as well (12 (21%)). Other autoantibody activities, like anti-Ro60 or anti-La, appeared not to be associated with MSA activities (table 4). Surprisingly, most of the anti-Ro52 activities in IIM sera were not accompanied by anti-Ro60 autoantibodies as is seen in patients with primary Sjögren's syndrome, but not, for example, in patients with SLE.52
PREVALENCE OF AUTOANTIBODY TYPES RELATIVE TO THE LATITUDE
A previous study, which included some of the patients studied here, reported that patients with DM were more prevalent in the cohorts from the southern countries of Europe, while the prevalence of patients with PM was more pronounced in the northern countries.39Therefore, the autoantibody prevalence in northern (Finland, Iceland, and Sweden (80 patients)), middle (UK, the Netherlands, Germany, Switzerland, and Czech Republic (238)), and southern (Italy, Slovenia, and Greece (99)) European countries was assessed.
Although the total number of positive sera is small for some autoantibody types, an interesting trend was noticed in the autoantibody prevalence relative to the latitude. A latitudinal gradient similar to the DM prevalence was found for the MSAs (anti-synthetase, anti-Mi-2, and anti-SRP autoantibodies)—that is, they occur more commonly in the southern countries. In contrast, the analysed MAAs, with exception of the anti-Ro52 and anti-U1 snRNP autoantibodies, were more commonly detected in patients from the northern countries as compared with the southern countries (data not shown). Anti-Ro52 autoantibodies seem to be more evenly distributed, whereas the anti-U1 snRNP autoantibodies were most abundant in the middle of Europe.
Discussion
This study describes the serological analysis of the largest group of patients with myositis to date. Sera were collected from various European rheumatological and neurological institutes located in the north (Iceland, Finland, Sweden), the middle (UK, the Netherlands, Germany, Czech Republic, Switzerland), and the south of Europe (Italy, Greece, Slovenia). This is relevant because we recently showed that the relative prevalence of PM and DM appeared to be dependent on the geographical latitude.39 All patient sera were analysed for the presence of a number of well known myositis specific and myositis associated autoantibody specificities by established methods as well as by a newly described dotblot assay.
Autoantibodies were detected in 53% of the sera. Because sera were only analysed for well defined autoantigens, these results do not imply that only half of the myositis sera contain autoantibodies. Indeed, it has been suggested that up to 90% of patients with myositis may produce autoantibodies when tested by indirect immunofluorescence or other serological techniques.3
In most cases the prevalence of the autoantibody activities tested in this study corresponded well with those previously reported in (smaller) cohorts of North American and Japanese patients with myositis.5 ,28 ,37 However, some interesting differences were noted. Firstly, our European collection contained more PM/Scl positive sera than the North American or Japanese groups (6%v 2% and 0%, respectively). Although we could not find significant differences in PM/Scl positivity between the individual European countries included in this study, it should be mentioned that these antibodies were not found in 40 Polish patients with PM/DM without overlap disease studied by Hausmanowa and coworkers.29 Secondly, the number of anti-Mi-2 positive sera is significantly higher in this study than the prevalence reported by others,5 ,29 ,53 but comparable with results obtained by Roux and coworkers.43 Because the number of anti-Mi-2 positive DM sera (21% of DM sera) corresponds with results of other studies, the difference is obviously caused by the relatively high number of anti-Mi-2 positive PM sera (9% of PM sera). About one in three anti-Mi-2 positive sera were from patients with PM and in most cases (8/17) the anti-Mi-2 activity could be confirmed by western blotting. It is likely that the increased sensitivity of the extended ELISA used in this study compared with the techniques used by others may account for the observed discrepancy. The inclusion of the NT and CT fragments, as well as the NM fragment, spanning the complete open reading frame of the Mi-2 autoantigen, clearly improved the sensitivity of our ELISA assay by 25–30% (see tables 2 and 3) compared with the ELISA originally described by Seelig and coworkers.12Additionally, anti-Mi-2 antibodies in patients with PM appeared to recognise the NT and CT fragments more frequently than the NM fragment, whereas antibodies from patients with DM seem preferentially to recognise the NM fragment. Application of the diagnostic criteria of Bohan and Peter,41 which were used in similar studies assessing autoantibody prevalence in patients with myositis,5 ,28 ,29 ,37 did not alter the diagnoses of the anti-Mi-2 positive patients with PM or DM. Only the three anti-Mi-2 positive patients with IBM were classified as PM according to the Bohan and Peter criteria. Thus it may be concluded that the extended ELISA as described in this study is responsible for the high incidence of anti-Mi-2 autoantibodies in the European group studied. Our results confirm the fact that anti-Mi-2 autoantibodies occur predominantly in sera of patients with DM, but suggest that they are more commonly present in PM sera than has been described previously.5 ,53 For future studies we therefore recommend the use of an ELISA assay which includes the NT, NM, and CT fragments as substrates for the detection of anti-Mi-2 autoantibodies.
It has been suggested that the immunogenetic background influences the autoantibody status of patients and may explain (in part) the observed discrepancies outlined above.5 ,37 ,54 Indeed, it has been proposed that genetic or environmental factors, or both, may regulate the occurrence of anti-PM/Scl autoantibodies, and such influences could explain the difference found between our study and the North American and Japanese results.5 ,28 Analyses of these genetic factors may also be interesting in view of the observed trend in latitudinal occurrence of some of the analysed autoantibodies.
In addition to the extended ELISA for detection of anti-Mi-2 autoantibodies, a second newly described application of an established method, the dotblot assay, was used to detect some more uncommon autoantibody activities. The major advantage of this application is that sera can be screened for multiple autoantibody activities after one single IP because the blots can be reprobed several times. In this study, the dotblot assay was used to detect anti-SRP, anti-Mas, anti-Jo-1, and anti-U1 snRNP reactivity, but the method would probably also detect autoantibodies directed to other ribonucleoprotein complexes, such as the Ro RNPs using anti-Y RNA probes. Furthermore, using different antisense tRNA probes, it should be possible to discriminate between the known autoantigenic aminoacyl-tRNA synthetases without needing to carry out, for example, in vitro inhibition assays.14
This study focused on the prevalence of some of the autoantibody activities that are common to patients with myositis. It gives an overview of the autoantibody status of European patients with myositis, which will hopefully lead to further understanding of these diseases and their cause.
Acknowledgments
The authors thank Ben de Jong and Eugenie Terwindt for technical assistance. We are grateful to Dr K Strub (Geneva, Switzerland) and Dr A Krol (Strasbourg, France) for their kind gifts of 7SL and tRNASel cDNAs, respectively and Dr M Renz (Karlsruhe, Germany) for the purified recombinant Mi-2 fragments. We acknowledge Dr K Steinsson for his contribution.
References
Footnotes
The work of RB and WJvV was supported by the “Prinses Beatrix Fonds” (grant 93–1112) and by the Netherlands Foundation for Chemical Research, with financial aid from the Netherlands Technology Foundation (grant 349–3699). The work of JV was supported by grants from IGA MZ CR NI/5369–3 and from the Ministry of Education of the Czech Republic VS96129.