Article Text

Abstract
Background Interleukin 6 (IL6) blockade raises blood lipid levels in patients with rheumatoid arthritis.
Objective To examine the influence of IL6 on lipid metabolism.
Methods Vascular smooth muscle cells (VSMC) were cultured in the presence of IL6, soluble IL6 receptor (sIL6R), IL6+sIL6R or tumour necrosis factor α (TNFα) for 24 h. After culture, the expression of very-low-density lipoprotein receptor (VLDLR), low-density lipoprotein receptor (LDLR) and low-density lipoprotein-related protein-1 (LRP-1) were measured by real-time PCR. Human IL6 was injected into mice twice a day for 2 weeks and then VLDLR expression in several tissues and the change of total cholesterol (TC) and triglyceride (TG) levels were investigated. Finally, the effect of anti-IL6 receptor (IL6R) antibody injection on blood lipid levels was examined.
Results IL6+sIL6R significantly induced expression of VLDLR mRNA in VSMC (8.6-fold, p<0.05), but IL6 or sIL6R alone and TNFα did not do so. None of these cytokines induced LDLR and LRP-1 mRNA expression. IL6 injection into mice increased the expression of VLDLR in heart, adipose tissue and liver and decreased TC and TG levels. The injection of anti-IL6R antibody normalised the reduced levels of TC and TG caused by IL6 injection, whereas it had no influence on the levels of TC and TG in normal mice.
Conclusions Overproduced IL6 decreased blood lipid levels by increasing VLDLR expression in several tissues. It is concluded that IL6 blockade normalises reduced lipid levels caused by IL6, but does not affect normal lipid metabolism.
Statistics from Altmetric.com
Introduction
Inflammatory cytokines such as interleukin 6 (IL6) and tumour necrosis factor α (TNFα) play crucial roles in the pathogenesis of rheumatoid arthritis (RA). It is reported that blockade of IL6 and TNFα in patients with RA increased blood levels of total cholesterol (TC), triglycerides (TG) and high-density lipoprotein cholesterol, which were inversely related to the disease activity of RA.1 2
It is also reported that the persistent inflammatory condition reflected by elevated serum TNFα levels results in low levels of TC and TG in RA.3 4 Moreover, studies in animal models have shown that TNFα contributes to changed lipid metabolism in lipopolysaccharide-treated mice.5 On the other hand, the relationship between serum IL6 and lipid levels in patients with RA seems not to have been reported, although it is known that treatment with the anti-IL6 receptor (anti-IL6R) antibody, tocilizumab increases TC and TG levels in patients with RA.1 We previously reported that IL6-transgenic mice had low TC and TG levels compared with their littermates, and that anti-IL6R antibody treatment increased their lipid profiles up to normal level.6 Others have shown that high TG and very-low-density lipoprotein (VLDL) levels are present in IL6 knockout mice.7 However, it is not clear why blockade of IL6 and TNFα results in changed lipid levels in the blood.
The low-density lipoprotein receptor (LDLR) family includes the LDLR, the LDL receptor-related protein (LRP) and the very-low-density lipoprotein receptor (VLDLR). The last of these is most abundantly expressed in heart, skeletal muscle and adipose tissue, but only to a slight extent in the liver, whereas LDLR is abundantly expressed in the liver. Because heart and skeletal muscle use fatty acids (FAs) as an energy source and adipose tissue uses FAs for energy storage, it was hypothesised that the VLDLR has a role in the delivery of FAs derived from VLDL-TG to peripheral tissues from blood. It has been reported that VLDLR-knockout mice fed a normal chow diet had 2.2-fold greater levels of TC and TG in plasma than wild-type mice.8 Another group reported that VLDLR-transgenic mice under conditions of LDLR deficiency had decreased TG levels, whereas VLDLR and LDLR double-knockout mice showed an increase.9 These findings imply that the VLDLR has a crucial role in lipid metabolism.
From these lines of evidence, we hypothesised that inflammatory cytokines decreased blood lipids level by increasing LDLR family expression in patients with RA and that anti-cytokine treatment normalises the alteration of the blood lipids level. To prove this hypothesis, first we examined the effect of IL6 on the expression of the LDLR family using vascular smooth muscle cells (VSMC), which express the LDLR family.10 Second, we confirmed the influence of IL6 on the relationship between VLDLR expression and blood lipids levels in IL6-treated mice.
Materials and methods
Reagents
Recombinant human IL6, human soluble IL6 receptor (sIL6R) and anti-mouse IL6R antibody (MR16-1) were prepared in our laboratories. Recombinant human TNFα was purchased from Wako (Osaka, Japan). Anti-mouse VLDLR antibody was purchased from R&D Systems (Minneapolis, Minnesota, USA).
Cell culture
VSMC were purchased from DS Pharma Biomedical (Osaka, Japan) and maintained in CSC complete defined medium (CellSystems, Kirkland, Washington, USA). VSMC (2×105 cells/well) were seeded in 18 wells of 12-well plates. After confluence had been reached, cytokines were added and incubated for a further 24 h.
Quantitative real-time PCR analysis
Total RNA was extracted using RNeasy kits (Qiagen, Valencia, California, USA). Synthesis of cDNA was performed using Omniscript RT kits (Qiagen) with a random nine primers (TAKARA, Shiga, Japan), according to the manufacturer's protocol. The quantitative real-time PCR was performed using the TaqMan gene expression assay, targeting human VLDLR, human LDLR, human LRP-1 and human glyceraldehyde-3-phosphate dehydrogenase, with the ABI PRISM 7000 system (Applied Biosystems, Foster City, California, USA), according to the manufacturer's protocol.
Animals
Female C57BL/6 mice (7 weeks old) were purchased from Charles River (Yokohama, Japan). Mice were specific pathogen-free and were kept in cages in a room maintained at 20–26°C, and 35–75% relative humidity. The experimental protocols were approved by the Institutional Animal Care and Use Committee of Chugai Pharmaceutical Co, Ltd.
Experimental protocol
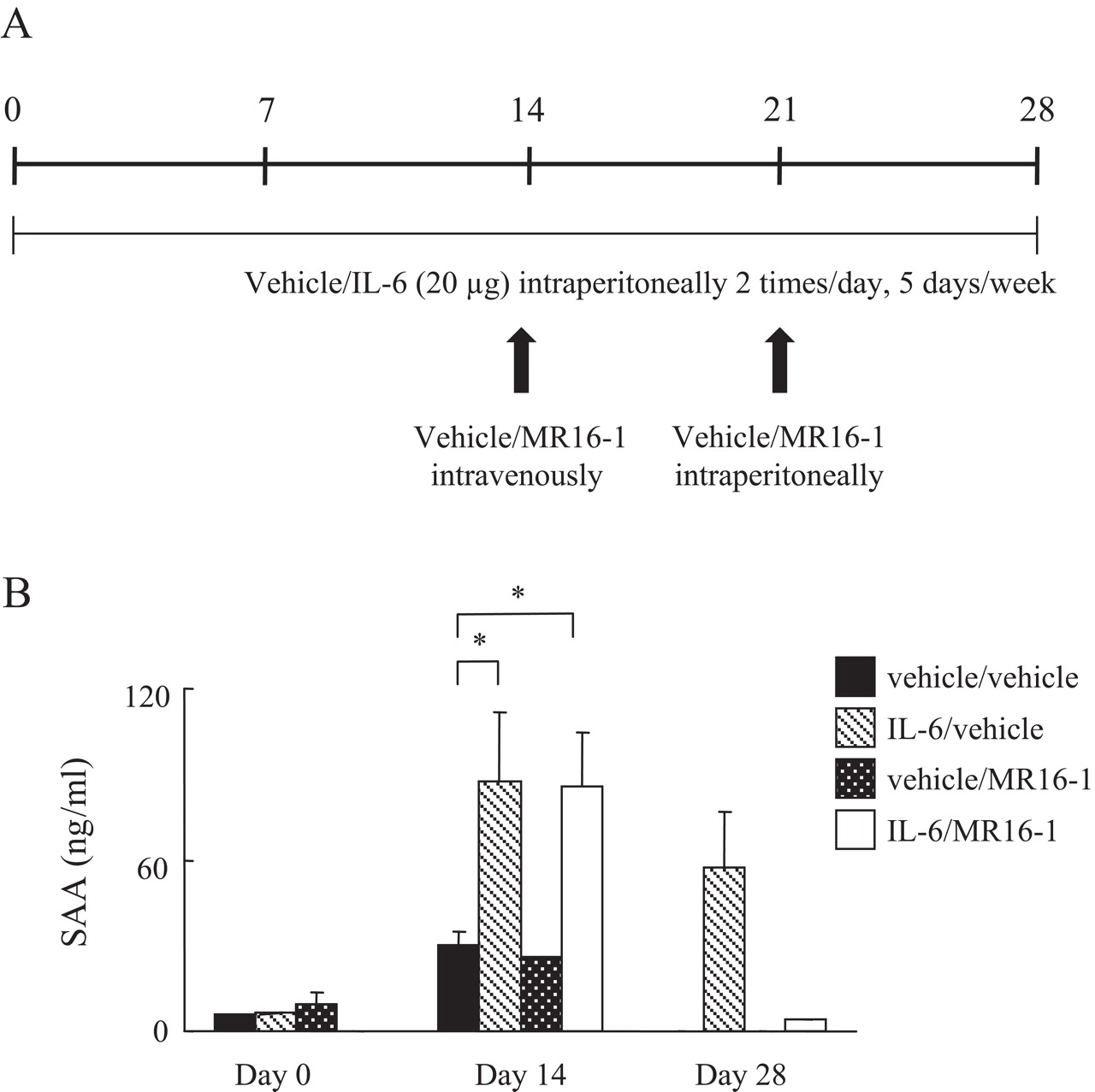
Female C57BL/6 mice (n=6) received 20 µg of IL6 or the same volume of saline intraperitoneally twice a day 5 times a week for 2 weeks (see figure 4A). In the study to examine the efficacy of anti-IL6R antibody, 20 µg of IL6 or the same volume of saline was given intraperitoneally twice a day five times per week for 4 weeks. MR16-1 (4 mg) or saline was injected intravenously at day 14 and then 1 mg of MR16-1 or saline intraperitoneally at day 21 (see figure 5A). Body weight and food intake were measured in the morning. Serum samples were collected in the morning on day 0 (just before injection), days 7 and 14 (the day of killing), and on day 0 (just before injection), 14 and 28 (the day of killing).
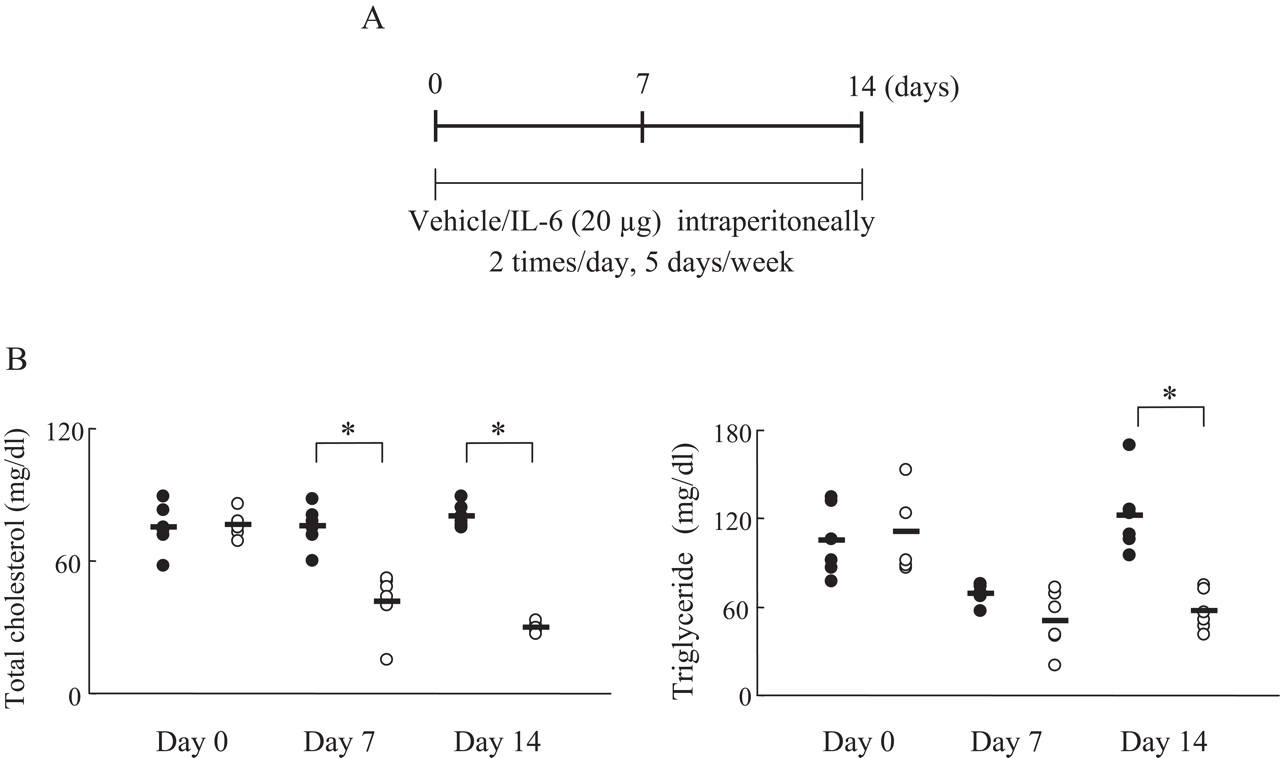
Serum lipid levels in interleukin 6 (IL6)-treated mice. (A) Experimental protocol. Mice (n=6) were given IL6 (20 µg) intraperitoneally or vehicle twice a day five times a week for 2 weeks. (B) Serum total cholesterol and triglyceride were measured using an automatic analyser. Closed and open circles indicate control and IL6-treated mice, respectively. Statistical significance between the control and the IL6 group on days 0, 7 and 14 was analysed by the unpaired t test (*p<0.05).

Anti-interleukin 6 (IL6) receptor antibody neutralised IL6 activity in IL6-treated mice. (A) Mice (n=6) received intraperitoneally IL6 (20 µg) or vehicle twice a day five times a week for 4 weeks. Anti-IL6R antibody (MR16-1, 4 mg) or vehicle was injected intravenously once at day 14, and then intraperitoneally at day 21 (MR16-1 (1 mg) or vehicle). (B) Serum amyloid A (SAA) measured by ELISA. Statistical significance between groups on days 0, 14 and 28 was analysed by the unpaired t test (*p<0.05).
Measurement of lipids, serum amyloid A, leptin and adiponectin
Measurements of TC and TG were performed using Fuji Dry-Chem (Fujifilm Medical, Tokyo, Japan). Concentrations of serum amyloid A (SAA), leptin and adiponectin in mouse serum were evaluated by ELISA kits according to the manufacturer's protocol as follows: mouse SAA (Biosource, Camarillo, California, USA), leptin (R&D Systems) and adiponectin (R&D Systems).
Western blotting for mouse VLDLR
Animals were killed and then heart, adipose tissue and liver were collected. Ten milligrams of tissues were lysed with reagents from the total protein extraction kit (Millipore, Billerica, Massachusetts, USA), according to the manufacturer's protocol. The protein concentrations were determined using BCA protein assay reagent kits (Pierce, Rockford, Illinois, USA). Cell lysates (10 µg protein) were separated by 7.5% sodium dodecyl sulphate-polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes (Millipore). After blocking with 3% bovine serum albumin in Tris-buffered saline for 2 h at room temperature, mouse monoclonal antibody to mouse VLDLR or β-actin (Sigma Aldrich, St Louis, Missouri, USA) in 1% bovine serum albumin/Tris-buffered saline (1:500 and 1:2000 dilution, respectively) was added and incubated for overnight at 4°C. After a 1 h incubation with secondary antibody (horseradish peroxidase-conjugated goat anti-mouse IgG (H + L)), binding was visualised using ECL western blotting substrate (Pierce), according to the manufacturer's protocol.
Statistical analysis
Statistical significances were calculated by the unpaired t test using a statistical analysis system software package (SAS Institute Japan, Tokyo, Japan). The significance level was set to 5%. Statistical analyses of plasma parameters were also performed by the unpaired t test.
Results
Induction of mRNA for lipoprotein receptors in VSMC
We examined the effects of IL6 and TNFα on the induction of lipoprotein receptors such as VLDLR, LDLR and LRP-1 in VSMC by real-time PCR. IL6+sIL6R strongly induced expression of VLDLR mRNA, but IL6 or sIL6R alone and TNFα did not do so (figure 1). In contrast, neither LDLR nor LRP-1 mRNA were induced by IL6, sIL6R, IL6 + sIL6R or TNFα

(A) Very-low-density lipoprotein receptor (VLDLR), (B) low-density lipoprotein receptor (LDLR) and (C) low-density lipoprotein receptor-related protein (LRP-1) mRNA expression in vascular smooth muscle cells (VSMC) induced by interleukin 6 (IL6) and tumour necrosis factor α (TNFα). VSMC were cultured with IL6 (100 ng/ml), soluble IL6 receptor (sIL6R; 100 ng/ml), IL6 (100 ng/ml) + sIL6R (100 ng/ml) or TNFα (10 ng/ml) for 24 h. After culture, relative expression levels of VLDLR, LDLR and LRP-1 mRNA were measured by real-time PCR. Each column and vertical line indicates mean and SD of triplicate cultures. Statistical significance was analysed by unpaired t test (*p<0.05).
VLDLR expression in IL6-treated mice
IL6 treatment did not result in changed body weight and food intake during the experiment (figure 2). Western blotting of VLDLR using tissues such as heart, adipose tissue and liver from IL6-treated mice is depicted in figure 3 at day 14. Controls showed that VLDLR expression levels were in the same range in heart, adipose tissue and liver. IL6 treatment augmented VLDLR expression in all tissues. We also confirmed that VLDLR mRNA expression was augmented by IL6 treatment but LDLR and LRP-1 mRNA expression were not, in the all organs tested (data not shown).
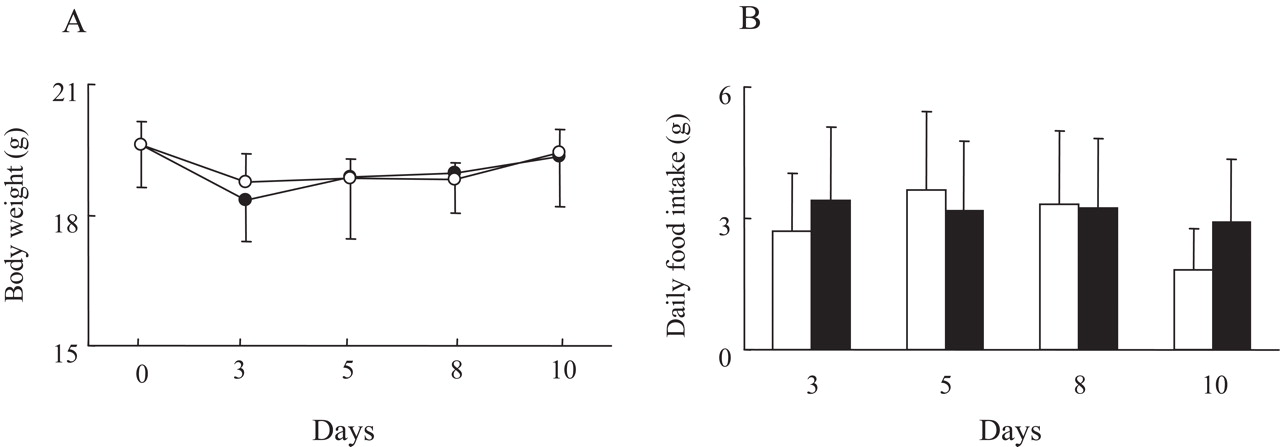
(A) Body weight gain and (B) daily food intake in interleukin 6 (IL6)-treated mice. Closed and open symbols indicate control and IL6-treated mice, respectively. Each circle and vertical line indicates mean and SE of six mice.
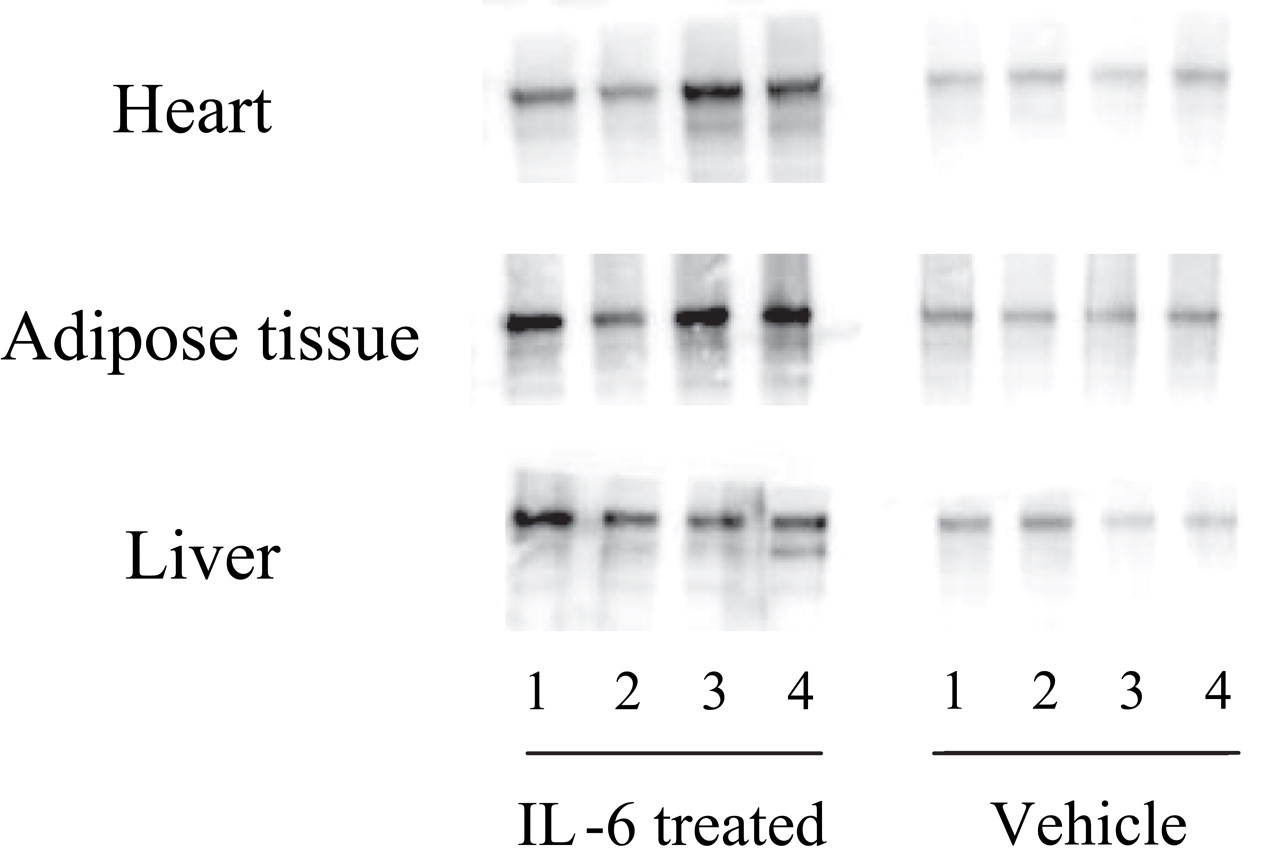
Very-low-density lipoprotein receptor expressions in heart, adipose tissue and liver in interleukin 6 (IL6)-treated mice. The mice (n=4) were killed and heart, adipose tissue and liver were collected on day 14 after injection with vehicle or IL6. Total protein was extracted from the tissue, and western blotting performed.
Lipids level in IL6-treated mice
Female C57BL/6 mice received 20 µg of human IL6 intraperitoneally twice a day for 14 days. It was found that IL6 significantly reduced the level of TC and TG in the blood, whereas saline treatment (control) had no effect (figure 4B).
Anti-IL6R antibody normalises TC and TG levels in IL6-treated mice, but does not change the level of leptin and adiponectin.
We tested whether anti-IL6R antibody (MR16-1) treatment could reverse the IL6-induced decrease in lipid levels. We reported that MR16-1 increased blood IL6 levels because of the delay of IL6R-mediated clearance of IL6.11 Therefore, serum IL6 level is not an appropriate maker to show IL6 blockade. To confirm whether the IL6 signal was blocked by MR16-1, we measured the SAA level, which is induced by IL6. Just before MR16-1 treatment, we confirmed that SAA was elevated (figure 5B) and that TC and TG levels were reduced by IL6 treatment. Thereafter, mice were treated with saline or MR16-1 weekly for 2 weeks from day 14. At day 28, the administration of MR16-1 had reduced SAA levels and increased TC and TG levels up to those in control mice (figure 6). However, administration of MR16-1 to control mice not treated with IL6 did not result in any changes to TC and TG levels. We also noted that levels of adiponectin and leptin were unchanged in all groups (data not shown).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Anti-interleukin 6 (IL6) receptor antibody reversed lipid level changes of IL6-treated mice. Closed and open circles indicate vehicle/vehicle and IL6/vehicle-treated mice, respectively. Closed and open triangles indicate vehicle/MR16-1 and IL6/MR16-1-treated mice, respectively. Statistical significance between groups on days 0, 14 and 28 was analysed by the unpaired t test (*p<0.05).
Discussion
VLDLR has a key role in the regulation of blood levels of lipoproteins such as TC, TG and free FAs.12 13 It has been reported that VLDLR-knockout mice fed a normal chow diet had greater levels of TC and TG in plasma than wild-type mice. Therefore, we think that VLDLR is involved in the adjustment of both TC and TG levels. Here, we demonstrated that IL6 induced the expression of VLDLR in VSMC in vitro and that IL6 injection into mice increased the expression of VLDLR in heart, adipose tissue and liver, and subsequently lowered lipids in blood. These findings strongly suggest that IL6 reduces the levels of lipids in the blood via the upregulation of VLDLR.
IL6 exerts its biological activities through two membrane molecules, a ligand-binding 80 kDa chain (IL6R) and a non-ligand-binding signal transducer gp130. After binding of IL6 to membrane-bound IL6R (mIL6R), the IL6/IL6R complex associates with gp130 and then the signal is transmitted into the cell. In addition, soluble IL6R (sIL6R), which lacks transmembrane and cytoplasmic domains, can associate with gp130 in the presence of IL6 and transduce the signal through gp130. Thus, both mIL6R and sIL6R have essential roles in IL6 signalling.14 In this study, IL6 + sIL6R, but not IL6 alone could induce VLDLR in VSMC, suggesting that VSMC expresses gp130, but not mIL6R. As there is sufficient sIL6R in blood, we think that the phenomenon seen in this study is likely to occur in vivo.
Regulation of VLDLR gene expression has been investigated, showing that CCAAT/enhancer-binding protein-β (C/EBP-β) and nuclear factor-Y bind to regulatory elements in the VLDLR promoter region.15 C/EBP-β is downstream of IL6 signalling and has a role in the regulation of energy homoeostasis and in adipocyte differentiation.16 17 These lines of evidences raise the possibility that C/EBP-β induced by IL6 has an important role in the regulation of VLDLR expression.
In a collagen-induced arthritis model, TC and TG did not decrease, although SAA clearly increased. This may be because the elevation of serum IL6 is transient unlike in a patient with RA in this model. In addition, one shot of IL6 into mice rapidly increased SAA, but did not decrease TC and TG. These facts led us to believe that continuous stimulation by IL6 is necessary to reduce TC and TG, so we decided to administer IL6 daily. In the preliminary study, we found that 40 µg/day of IL6, but not 20 µg/day of IL6, decreased TC and TG. Therefore we used 40 µg/day in this study.
Food intake affects the level of lipids in the blood. However, food intake did not differ between control and IL6-treated mice throughout this study. Also, serum was always sampled at the same time (in the morning). Therefore, we can exclude the possibility that decreased food intake or other related factors are involved in the decreases of TC and TG levels.
LDLR, LRP-1, adiponectin and leptin are also involved in lipid metabolism. Absence of LDLR and LRP-1 in mice results in the accumulation of TC and TG in plasma.18 Adiponectin also reduces plasma TG and increases high-density lipoprotein cholesterol.19 The leptin level in plasma correlates with the LDL-cholesterol and TG concentration.20 Since IL6 treatment did not change the expression s of LDLR, LRP-1, adiponectin or leptin, we consider that these molecules did not participate in IL6-induced lipid level changes.
Increases in lipid levels have been reported when using TNFα blockers such as infliximab and etanercept in RA treatment.2 21 However, when we assessed the effect of TNFα on LDLR, LRP-1 and VLDLR expression by VSMC, we found no induction of these molecules. TNFα is a potent inducer of IL6 and its blockade rapidly reduces serum IL6 levels in patients with RA.22 It is likely that the suppression of IL6 production partly contributes to the lipid profile changes following TNFα blockade.
In conclusion, our data clearly suggest that in patients with RA, overproduced IL6 decreased blood lipids level through the upregulation of VLDLR expression and, therefore, that IL6 blockade normalises these reduced lipid levels by IL6. Further studies in humans are warranted to confirm our result in mice.
References
Footnotes
-
Competing interests None.
-
Provenance and peer review Not commissioned; externally peer reviewed.