Article Text

Abstract
Background: Autoantibodies to cyclic citrullinated peptides (anti-CCP) are present in most patients with rheumatoid arthritis (RA), and associate with HLA-DRB1 shared epitope (SE) alleles.
Objective: To investigate reactivities of anti-CCP to various citrullinated proteins/peptides, which represent potential autoantigens in RA, and to examine the relationship between such antibodies, and their association with genetic variants within HLA-DRB1 SE alleles.
Methods: Serum samples from 291 patients with established RA and 100 sex- and age-matched healthy subjects were included in this study. Sera were first analysed for presence of anti-CCP antibodies and further for IgG and IgA antibodies towards candidate autoantigens in both their native and citrullinated form including: fibrinogen, α-enolase peptide-1 and the C1-epitope of type II collagen (C1III). Antibody specificity was confirmed by cross-reactivity tests. HLA-DR genotyping was performed.
Results: 72% of patients with RA were anti-CCP positive. Among the candidate autoantigens examined, IgG antibodies to citrullinated fibrinogen were found in 66% of patients’ sera and in 41% for both citrullinated α-enolase peptide-1 and citrullinated C1III. These antibodies were mainly seen in the anti-CCP-positive patient group; they were specific for their respective antigen and displayed limited cross reactivity. IgA responses were also detected, but less frequently than IgG. Anti-CCP and anti-citrullinated protein antibodies were associated with HLA-DRB1*04 rather than with HLA-DRB1*01 alleles.
Conclusions: Antibodies directed against several citrullinated antigens are present in CCP-positive RA, with many patients displaying multireactivity. All specific reactivities were primarily associated with the HLA-DRB1*04 alleles, suggesting common pathways of anti-citrulline immunity.
Statistics from Altmetric.com
Most patients with RA develop anti-citrullinated protein antibodies (ACPA) (reviewed by van der Helm-van Mil).1 These antibodies are highly specific for rheumatoid arthritis (RA) and can be detected several years before disease onset.2 Anti-cyclic citrullinated peptide (anti-CCP) antibody testing is a simple laboratory examination, which when positive is associated with bone erosion and a severe course of the disease3 without revealing the underlying adaptive immune responses. However, RA can be divided into two different subsets: anti-CCP-negative and anti-CCP-positive RA.1 The latter has been associated with smoking and certain HLA-DRB1 alleles, which both are associated with an increased risk for anti-CCP-positive RA.4 5
Several HLA-DRB1 alleles (within the DRB1*04 and DRB1*01 loci) have been described as risk factors for the development of RA. These alleles share an amino acid sequence at the peptide-binding site in the third hypervariable region of the DRβ1 chain—the “shared epitope” (SE).6 The HLA-DRB1 SE alleles are associated particularly with anti-CCP-positive RA7 and also influence the magnitude of the anti-CCP response.8
The anti-CCP assay has a high specificity for RA.9 Yet, it does not provide information about the identity of which citrullinated proteins are recognised by the anti-CCP antibodies. A better knowledge of which autoantigens, citrullinated and non-citrullinated, which may be targeted by antibodies in patients with RA, is thus needed for further investigations of how adaptive immunity contributes to disease induction in RA. Several previous reports have described autoantibodies against specific citrullinated proteins in RA sera.10–16 Some of these proteins like fibrinogen, type-II collagen and α-enolase are expressed in the synovial tissue or in the joint. However, these reactivities have not been systematically compared and their relationship with the HLA-DRB1 SE alleles has not been studied in detail.
Recently, an epidemiological study showed that smoking is an environmental risk factor for development of anti-CCP-positive RA in people who are SE positive.4 It was postulated that smoking is associated with an increased presence of citrullinated proteins in the lungs, which may trigger an immune response against citrullinated protein. IgA is central in mucosal immunity—for example, in the lungs. Therefore, IgA responses to specific citrullinated antigens and the association between IgG and IgA within the response to a given antigen may also be of importance.
In this study we used serum samples from patients with established RA to investigate IgG and IgA responses towards CCP as well as three different citrullinated proteins (cit-fibrinogen cit-α-enolase and citC1III), which have previously been described as being targets of RA-specific autoantibodies. Our aim was to obtain specific knowledge about the relationship of these different autoantibodies with each other and their relationship with the two major HLA-DRB1 alleles that carry the SE motif.
PATIENTS AND METHODS
Patients and healthy subjects
Serum samples were collected from 291 patients with established RA (mean age 55, range 21–86, 82% female) according to the American College of Rheumatology criteria17. All patients attended the rheumatology clinic at the Karolinska University Hospital, Stockholm, Sweden, and were included based on a clinical need for withdrawal of synovial fluid from knee effusions. Thus, synovial fluids were obtained in parallel with serum sampling. Serum samples were stored at −70°C until use. One hundred serum samples from healthy subjects (mean age 54, range 24–82, 81% female) were also analysed and used as controls in this study, one for every three patient samples. The ethical review board of the Karolinska University Hospital approved this study, and all subjects gave informed consent.
Fibrinogen purification and deimination
Human fibrinogen (95% pure, Calbiochem, San Diego, California, USA) was dissolved in phosphate-buffered saline and further purified from immunoglobulins using goat-IgG anti-human immunoglobulins (H+L) antibodies (Jackson, ImmunoResearch Laboratories, West Baltimore, USA) attached to protein A-agarose beads (Santa Cruz, California, USA). Deimination was then performed with rabbit skeletal muscle peptidyl arginine deiminase (Sigma-Aldrich, St Louis, Missouri, USA) in 0.1 M Tris-HCl, pH 7.4, 10 mM CaCl2, 5 mM dithiothreitol for 2 h at 37°C. Citrullination was confirmed by dot blot using Senshu antibody (Upstate, New York, USA).18
Anti-CCP assay
An anti-CCP2 test (Immunoscan RA, Mark 2, Euro-Diagnostica, The Netherlands) was used to determine the levels of anti-CCP antibodies. Quantification of the results and use of a cut-off value of 25 U/ml were according to the manufacturer’s instructions.
ELISA assay
Microtitre plates (Nunc, Denmark) were coated and incubated overnight with one of the following antigens in their native or citrullinated (cit) form, all at a concentration of 10 μg/ml: (a) fibrinogen (full-length protein); (b) peptide-1 of α-enolase15 16 19; (c) triple helical peptides of the C1-epitope of type-II collagen (C1III), aa 359–369.10 20 Plates were washed and blocked with 1% bovine serum albumin (BSA) in phosphate-buffered saline for 1 h. For the detection of fibrinogen and cit-fibrinogen-specific IgG and IgA, serum samples were diluted 1:50 in RIA buffer (1% BSA, 350 mM NaCl, 10 mM Tris-HCl pH 7.6, 1% vol/vol Triton X-100, 0.5% wt/vol Na-deoxycholate, 0.1% sodium dodecyl sulphate) supplemented with 10% fetal calf serum. To determine α-enolase, cit-α-enolase, C1III and citC1III-specific IgG and IgA, serum samples were diluted 1:100 in RIA buffer supplemented with 10% fetal calf serum. Horseradish peroxidase-conjugated goat anti-human IgG or IgA (Jackson ImmunoResearch Laboratories) was added to detect bound antibodies. These, were then detected using the chromogenic substrate 3,3′,5,5′-tetramethylbenzidine (TMB, Sigma-Aldrich). Plates were read at 450 nm with a reference of 650 nm. A standard curve from a positive serum to specific antigen was included on each plate and was further used to translate optical density (OD) values to arbitrary units. Cut-off points for each of the citrullinated antigens was set to 99% specificity and determined according to the reactivity of control serum samples.
HLA-DR genotyping
DNA was extracted from EDTA blood by the salting-out method.21 Genotyping for HLA-DRB1 allotypes was conducted using the sequence-specific primer-polymerase chain reaction method (DR low-resolution kit; Olerup SSP, Saltsjöbaden, Sweden) as previously described.22 DRB1*04- and DRB1*01-positive patients were further subtyped by Olerup SSP DRB1*04 and DRB1*01 (Olerup SSP, Saltsjöbaden) subtyping kits, respectively. The following alleles were classified as SEs: DRB1*0101, *0102, *0104, *0401, *0404, *0405 and *0408.
Cross-reactivity assay
Serum samples from patients with RA reactive with cit-fibrinogen, cit-α-enolase and citC1III were selected for these assays. Sera (diluted 1:150 in RIA buffer) were first preabsorbed on ELISA plates coated with one of the antigens (either cit-fibrinogen, cit-α-enolase or citC1III) or BSA. After six repeated absorptions, the reactivity of preabsorbed sera was examined against each antigen and BSA. IgG reactivity was determined by ELISA (as described previously under “ELISA assay”). The quantification of serum reactivity was calculated as the ratio between the OD values exhibited by sera preabsorbed against one of the citrullinated antigens and the OD values displayed by BSA-preabsorbed sera. The OD values obtained by BSA-preabsorbed sera were used as maximum values (100%), since serum reactivity to BSA did not show any measurable absorbance capacity (data not shown).
Heat map
Hierarchical agglomerative dendrograms were constructed using average Euclidean distance measure to detect and cluster related samples and related antibody reactivities. Clustering illustrates the relationships between different samples and reactivities (see Eisen et al23 for a detailed description of the methodology).
A colour display was used to visualise the expression of different antibody levels in patients’ serum samples, as measured by ELISA. Each row represents one patient, and each column depicts the reactivity to one of the different antigens. Owing to the difference in dynamic range of each assay the levels of antibody reactivity against each antigen were scaled according to its standard deviation; this made it possible to transform different arrays to comparable reactivity levels. To better visualise the variation all values above 1.5 standard deviations were saturated.
Statistical analysis
One-way analysis of variance (Kruskal–Wallis test) followed by Dunn’s multiple comparison test were used to analysed the differences between groups. Spearman’s rank correlation was used to analyse the association between levels of IgG and IgA antibodies. p Values <0.05 were considered significant.
RESULTS
Serum reactivity to native and citrullinated fibrinogen, α-enolase (peptide 1) and the C1 epitope of type-II collagen
First, we examined the prevalence of anti-CCP-positive patients with RA in our cohort. Serum samples from 291 patients with well-established RA and 100 sex- and age-matched healthy subjects were examined. Seventy-two per cent of the RA sera were anti-CCP positive while only one (1%) healthy subject showed an anti-CCP-positive response (data not shown). Hence, the anti-CCP response in this cohort is similar to previous reports in which the anti-CCP2 test has been used to examine RA sera reactivity.
Sera were then further analysed for reactivity against fibrinogen, α-enolase-peptide 1 and triple helical peptide C1III in their native and citrullinated (cit) forms. The most frequent reactivity was detected toward cit-fibrinogen, 66% (fig 1A), while the reactivities against cit-α-enolase and citC1III were both about 41% (figs 1B and C). The native C1-epitope (C1III) was the only non-citrullinated antigen to which IgG antibody response was detected in serum, with 13% positivity in patients with RA as compared with 5% in healthy controls (fig 1C). However, the medians for antibody levels toward non-citrullinated C1III did not reach statistical significant difference between patients with RA and controls.

Levels of IgG antibodies against native and citrullinated rheumatoid arthritis (RA)-associated antigens. IgG antibody recognition of (A) native and citrullinated fibrinogen; (B) native and citrullinated α-enolase and (C) C1III and citrullinated C1III was monitored by ELISA in serum samples from patients with RA (n = 291) and healthy subjects (n = 100). Bars indicate the median levels of specific antibody response. Dotted lines represent the cut-off values, above which all points above were considered positive for the specific antigen. *p<0.05; **p<0.01; ***p<0.001.
Independence and cross-reactivity of citrulline-specific immune responses
Next we examined to what extent antibody reactivities against cit-fibrinogen, cit-α-enolase and citC1III were monospecific for the respective autoantigens, and to what extent cross-reactivities were present. Eleven RA serum samples reacting with all three citrullinated antigens (ie, cit-fibrinogen, cit-α-enolase and citC1III) were selected and their reactivities were examined after preabsorbance to each of these antigens.
Overall, maximum reduction in serum reactivity (ie, maximum preabsorbance) was seen after preabsorbance with the same citrullinated antigen for which the reactivity was examined (fig 2A–C). As shown in figs 2B and 2C, absorption with cit-α-enolase hardly affected the reactivity to citC1III, and preabsorption with citC1III hardly affected reactivities to cit-α-enolase. In contrast, preabsorbance with either cit-α-enolase or citC1III somewhat affected serum reactivity against cit-fibrinogen (figs 2B and 2C). Preabsorbance with cit-fibrinogen was relatively modest and had some effect on serum reactivity to citC1III (fig 2A), probably because the full-length protein was used rather than a synthetic peptide. Still, the greatest reduction in serum reactivity against cit-fibrinogen was achieved after preabsorbance with cit-fibrinogen, indicating some independence of the anti-cit-fibrinogen response.

Specificity and cross-reactivity between antibodies against different citrullinated antigens. The reactivities of 11 rheumatoid arthritis serum samples toward cit-fibrinogen, cit-α-enolase and citC1III were examined after preabsorbance to either (A) cit-fibrinogen, (B) cit-α-enolase or (C) citC1III. Data are expressed as the percentage of maximum reactivity, which was considered to be 100% for bovine serum albumin-preabsorbed serum responses. The results represent one of three determination performed. *p<0.05; **p<0.01; ***p<0.001.
Relationships between antibody reactivities to different citrullinated proteins
Several specific ACPA may be present in RA sera at the same time, particularly in the anti-CCP-positive patients. While most anti-CCP-positive patients react with cit-fibrinogen, the reactivity toward cit-α-enolase and citC1III only partly overlaps, and therefore four main subgroups can be distinguished. The largest subgroup within the CCP-positive patients reacts with all three citrullinated antigens examined (fig 3A). The other three main subgroups react with cit-fibrinogen in combination with either cit-α-enolase, citC1III or cit-fibrinogen alone (fig 3A).

Patterns of antibody reactivity in patients with rheumatoid arthritis (RA). (A) Subgroups of patients with RA (n = 291) according to their IgG antibody responses to cyclic citrullinated peptide (CCP), cit-fibrinogen, cit-α-enolase and citC1III. (B) Clustering of IgG RA-associated antibodies. IgG reactivity was measured in serum from 291 patients with RA against CCP and both native and citrullinated forms of the following antigens: fibrinogen, α-enolase and C1III. A colour scale shows the relative degree of antibody reactivity; from low reactivity (green) to high reactivity (light red). Outer lines illustrate the intimacy between clusters.
In order to visualise the various reactivity patterns at an individual level, and to examine their internal relationship, we constructed a heat map using clustering algorithms, in which the levels of the different antibody reactivities are presented in colour scales according to their levels in serum (fig 3B). As shown in fig 3B, serum samples from the majority of CCP-positive patients react with one or more specific citrullinated antigens. Most patients with high anti-CCP antibody levels also display high levels of antibodies against specific citrullinated antigens and are more likely to have antibodies against more than one specific citrullinated antigen. Nevertheless, many patients who display relatively low, yet positive, levels of anti-CCP antibodies, also react with more than one citrullinated protein.
In contrast to antibody responses against citrullinated antigens, the reactivity toward the non-citrullinated C1III was seen in both anti-CCP-positive and anti-CCP-negative sera.
Association between the HLA-DRB1 SE alleles and citrulline-specific immune responses
It has been shown that patients with RA carrying the HLA-DRB1 SE alleles produce higher levels of anti-CCP antibodies.8 Also, HLA-DRB1 SE alleles confer an increased risk for anti-CCP-positive RA.4 7 We therefore investigated the effects of SE alleles on citrulline-specific responses. A total of 223/291 patients could be genotyped and were divided into three groups: (a) non- SE carriers (−/−), n = 38; (b) carrying one SE allele (−/SE), n = 113; (c) carrying two SE alleles (SE/SE), n = 72. Patients with RA carrying two SE alleles displayed higher levels of anti-CCP antibodies as well as antibodies against cit-fibrinogen and cit-α-enolase than patients carrying single allele or no SE alleles (figs 4A–C). Further, patients carrying one SE allele displayed higher, yet not significant, antibody levels toward CCP, cit-fibrinogen and cit-α-enolase than patients who had no SE alleles (figs 4A–C). Unlike the other citrullinated antigens, there was no significant effect of the SE alleles on the antibody levels toward citC1III (fig 4D).

The effects of the number of shared epitope (SE) copies on anti-cyclic citrullinated protein (anti-CCP) antibodies. Two hundred and twenty-three patients with rheumatoid arthritis were genotyped for HLA-DRB1 allotypes and divided according to the number of their SE alleles: no SE (−/−), n = 38; heterozygote to the SE (−/SE), n = 113; homozygote to the SE (SE/SE), n = 72. The levels of (A) IgG anti-CCP antibodies as well as the IgG antibody levels towards (B) cit-fibrinogen, (C) cit-α-enolase and (D) citC1III were compared between the three groups. Bars indicate the median levels of antibody response. *p<0.05, ** p<0.01, ***p<0.001.
We next examined the identity of the SE alleles that affected the citrulline-specific antibody response. Patients were divided according to the identity of their SE alleles: (a) do not carry SE alleles (−/−), n = 38; (b) carry one or two copies of DRB1*01 alleles (DR*01), n = 41; or (c) carry one or two copies of DRB1*04 alleles (DR*04), n = 110. Patients who carried one DRB1*01 allele and one DRB1*04 allele (n = 34) were excluded from this analysis in order to stratify the analysis. The anti-CCP antibody levels, as well as the levels of antibodies against all three specific citrullinated antigens, were higher in the DRB1*04 group than in the other two groups (figs 5A–D). The DRB1*01 alleles had no influence on the reactivities to citrullinated proteins in this study.
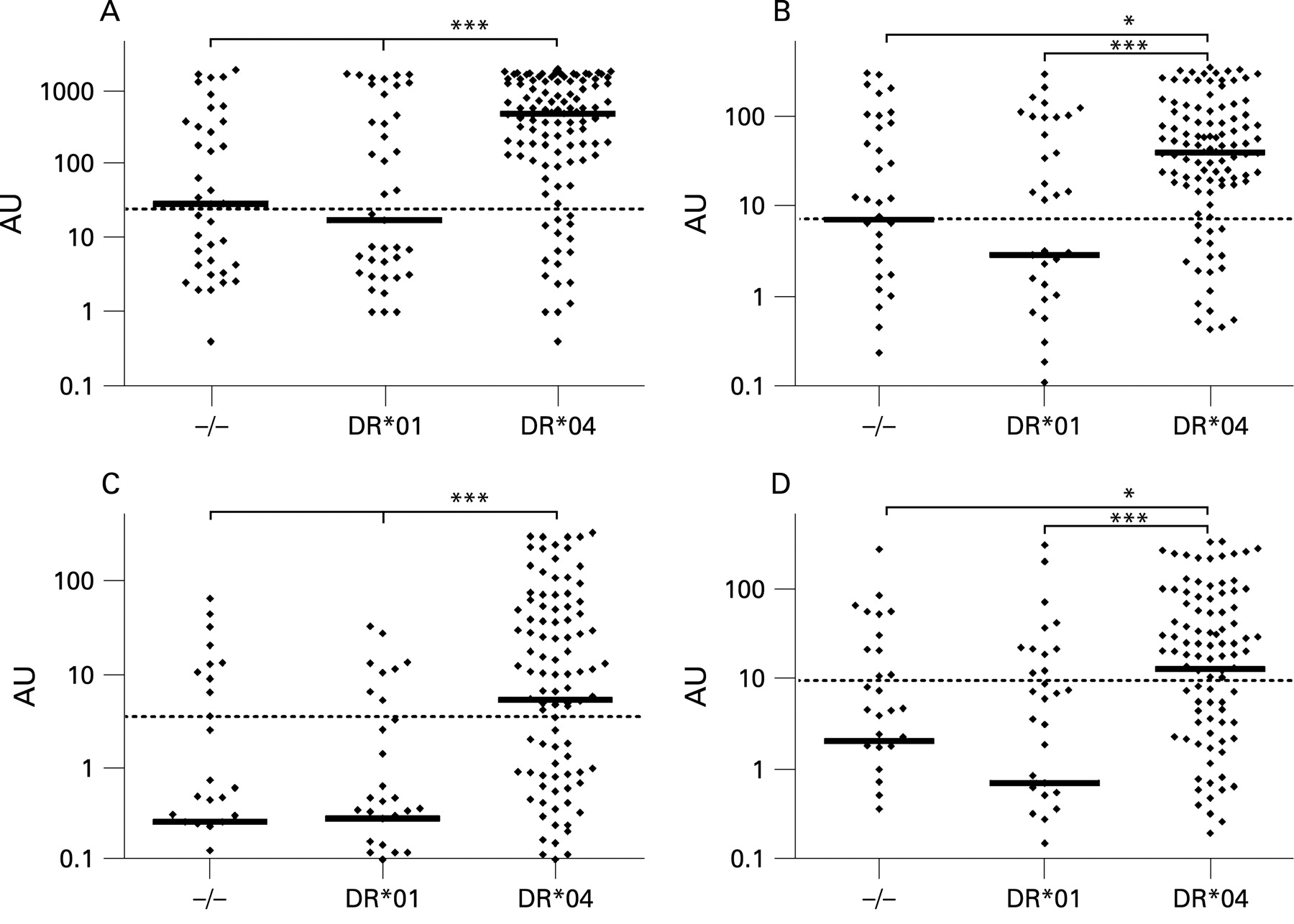
The effects of DR*01 and DR*04 shared epitope (SE) alleles on anti-citrullinated protein (anti-CCP) antibodies. One hundred and eighty-nine patients with rheumatoid arthritis were divided according to the identity of the SE alleles (ie, DR*01 or DR*04) they carried. No SE (−/−), n = 38; HLA-DRB1*01 (DR*01), n = 41; HLA-DRB1*04 (DR*04), n = 110. Then, the levels of (A) IgG anti-CCP antibodies, as well as the levels of IgG antibodies against (B) cit-fibrinogen, (C) cit-α-enolase and (D) citC1III were compared among the three groups. Bars indicate the median antibody levels in each group. *p<0.05; ***p<0.001.
Citrulline-specific IgA immune responses
Reactivities of IgA antibodies were found for all citrullinated antigens, but in lower frequencies than IgG reactivities (29.5%, 16% and 21% against cit-fibrinogen, cit-α-enolase and citC1III, respectively). Interestingly, IgA responses were almost exclusively detected in patients with IgG antibodies toward the same antigen (figs 6A–C).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Association between IgA and IgG antibodies. The levels of IgG and IgA antibodies against (A) cit-fibrinogen, (B) cit-α-enolase and (C) citC1III, as measured by ELISA, are shown on x, y coordinates. Dotted lines represent the cut-off values for each antigen, and all results above these lines were considered positive for the specific antigen.
DISCUSSION
Autoantibodies are fingerprints of single or multiple concurrent immune responses. Here, we examined the reactivity of antibodies against three specific citrullinated antigens in RA sera and drew an individual antibody profile for each patient. Our data demonstrate that RA sera show varying patterns of reactivity for these different specific citrullinated antigens. The reactivities to these antigens are mainly restricted to the anti-CCP-positive RA subset and are rare in a healthy population. Although a minor cross-reactivity was observed for anti-cit-fibrinogen antibodies, reactivities to citrullinated antigens were largely specific—that is, the degree of cross-reactivity between different citrullinated antigens was limited.
The HLA-DR SE alleles, which are found within the DRB1*04 and DRB1*01 families of HLA-DRB1 alleles are well-known risk factors for RA. They associate with anti-CCP-positive RA7 and also regulate the magnitude of anti-CCP antibodies.8 In this study we demonstrate that high levels of antibodies against specific citrullinated antigens particularly associate with the HLA-DRB1*04 SE and not with HLA-DRB1*01 SE. The strong association with DRB1*04 SE alleles is intriguing and suggests common pathways concerning “citrulline immunity”. Such potent and major histocompatibility complex (MHC) class II restricted antibody responses are normally dependent on T-cell activation, which is restricted by the same MHC class II alleles. Yet, the identity of autoreactive T cells and their specific antigens in RA is still largely unknown.
One obvious possibility is that the restriction is dependent on presentation of citrullinated peptides in the context of MHC class II molecules where β chains are coded by HLA-DRB1*04 alleles. Importantly, peptides presented to a T cell may not be identical to peptides recognised by B cells. However, support for a direct presentation of citrullinated peptides to T cells has been provided from a study in HLA-DRB1*0401 transgenic mice, demonstrating that citrullination increases the affinity of a certain vimentin peptide to MHC class II and leads to CD4+ T-cell activation.24 On the other hand, T-cell recognition of the RA-associated autoantigen type II collagen has in the mouse been shown to be dependent on another post-translation modification. Here, glycosylation of the major T-cell epitope 260–271 was required for an optimal presentation on HLA-DRB1*0401 allele.25 26 Also oxidation and acetylation, as well as other modifications, may in principle be able to brake immune tolerance, promote T-cell recognition and allow post-translationally modified peptides to be presented to HLA-DRB1*04 restricted autoreactive T cells.
A recent study showed also that IgM and IgA anti-CCP antibodies are present in RA and in undifferentiated arthritis but at lower frequencies than IgG.27 Interestingly, the patients with undifferentiated arthritis who had IgM and IgA anti-CCP antibodies were more likely to develop RA than those who were IgG CCP positive but did not have IgA or IgM.27 While isotype switching from IgM to IgG is likely to occur, the presence of anti-CCP IgA antibodies implies that mucosal tissues—for example, the lungs, may be involved in the pathogenesis of ACPA-positive RA. Indeed smoking, which has been identified as a main environmental risk factor for the development of anti-CCP-positive RA4 5 can increase the presence of citrullinated proteins in the lungs.4 In this study we detected IgA antibodies toward all three citrullinated antigens (cit-fibrinogen, cit-α-enolase and citC1III). These were detected mainly in patients who also had IgG antibodies reactive with the same antigens. Whether these responses are the result of an immune stimulation from mucosal surfaces is, however, not known.
The presently investigated citrullinated autoantigens may not be the only citrullinated targets of antibodies in patients with RA, and the role of immunity to the different citrullinated self-antigens deserve further study. Such studies should provide a basis for further examination of T-cell immunity to specific citrullinated peptides. These examinations obviously need to be performed in carefully defined genetic contexts, taking into account the presently defined associations between different HLA-DR allotypes and immunity to different citrullinated autoantigens.
Acknowledgments
We thank all personnel and patients at the rheumatology clinic at the Karolinska University Hospital, Eva Jemseby for organising sampling, storage and administration of biomaterial, Dr Christina Trollmo for scientific discussion, Dr Anca Catrina for clinical evaluation and scientific discussion and Henrik Källberg for statistical advice and support.
REFERENCES
Footnotes
Competing interests: None.
Funding: This study was supported by Margaretha af Ugglas Foundation, the Swedish Research Council and an EU FP6 project, AutoCure LSHB CT-2006-018661, 2. This publication reflects only the authors’ views; the European Community is not liable for any use that may be made of the information herein.
Ethics approval: Approved by the ethical review board of Karolinska University Hospital.
OS and MW contributed equally to this work