Article Text

Abstract
Objective: To evaluate the role of the single-nucleotide polymorphism (SNP) at position −670 in the FAS gene promoter (FAS−670G>A) in influencing the susceptibility, clinical features and severity of systemic sclerosis (SSc).
Methods: 350 white Italian SSc patients (259 with limited cutaneous SSc (lcSSc) and 91 with diffuse cutaneous SSc (dcSSc)) and 232 healthy individuals were studied. Patients were assessed for the presence of autoantibodies (anticentromere, anti-topoisomerase I (anti-Scl-70) antibodies), interstitial lung disease (ILD), pulmonary arterial hypertension and scleroderma renal crisis. FAS−670G>A SNP was genotyped by PCR restriction fragment length polymorphism assay. Serum levels of soluble FAS (sFAS) were analysed by ELISA.
Results: A significant difference in FAS−670 genotype distribution was observed between SSc patients and healthy individuals (p = 0.001). The frequency of the FAS−670A allele was significantly greater in SSc than in controls (p = 0.001). No significant difference in genotype distribution and allele frequencies was observed between lcSSc and dcSSc, although a greater frequency of the FAS−670A allele was found in dcSSc. The FAS−670AA genotype significantly influenced the predisposition to SSc (OR 1.97, 95% CI 1.35 to 2.88, p = 0.001) and to both lcSSc (OR 1.84, 95% CI 1.23 to 2.75, p = 0.003) and dcSSc (OR 2.37, 95% CI 1.41 to 3.99, p = 0.001). FAS−670A allele frequency was greater, although not significantly, in anti-Scl-70 antibody-positive dcSSc and ILD dcSSc. sFAS was significantly higher in patients and controls carrying the FAS−670AA genotype compared with those carrying the FAS−670GG genotype (p = 0.003 in SSc, p = 0.004 in controls).
Conclusion: The FAS−670A allele is significantly associated with susceptibility to SSc, suggesting a role for a genetic control of apoptosis in the pathogenesis of the disease.
Statistics from Altmetric.com
Systemic sclerosis (SSc; scleroderma) is a chronic autoimmune disease of unknown aetiology affecting the skin and various internal organs, and can be classified in two main clinical subsets: limited cutaneous (lcSSc) and diffuse cutaneous (dcSSc) SSc.1
The immunological abnormalities commonly observed in SSc involve both B and T cells, with implications for autoantibody and profibrotic cytokine production and infiltration of T-cell subsets in targeted organs.2–8 Furthermore, a marked resistance to apoptosis and reduced activation-induced cell death have recently been shown in different subsets of circulating T lymphocytes from these patients.9
Apoptosis is an essential physiological process regulating immune homeostasis and contributing to the pathogenesis of autoimmunity via an impairment in the deletion of autoreactive immune cells.10 FAS antigen (Apo-1/CD95) is a cell-surface receptor belonging to the tumour necrosis factor receptor superfamily and has been identified as a key molecule involved in the apoptosis of a wide variety of cell types, including activated immune cells.11 The human FAS gene has been mapped to chromosome 10q23 and is highly polymorphic.12 In mice, mutations in the FAS gene are associated with massive lymphadenopathy and lupus-like autoimmunity.11 In humans, FAS mutations have been found in patients with Canale–Smith syndrome, an inherited lymphoproliferative disorder associated with autoimmunity.13 Following these reports, a different single-nucleotide polymorphism (SNP) in the enhancer region of the FAS gene promoter, caused by a G to A base change at nucleotide position −670 (FAS−670G>A SNP, rs1800682), has been described.14 Recent evidence suggests that this SNP may be of functional importance, occurring at the consensus binding sequence of the gamma-interferon activation site.14 Moreover, it has been suggested that the FAS−670A allele has a higher binding ability to a gamma-interferon activation site-binding protein, the transcription factor signal transducer and activator of transcription type 1 and correlates with soluble FAS (sFAS) levels.15 16 The FAS−670G>A polymorphism might thus influence the expression of the FAS gene and/or FAS-mediated apoptotic signalling.
There is increasing evidence that autoimmune diseases are complex polygenic disorders, which result from interactions between genetic and environmental factors.17 The combinations of common genetic variants, including SNP, might influence both the susceptibility to the disease and specific features of the disease phenotype.18 Moreover, it appears that many autoimmune disorders share a common pool of disease susceptibility genes, despite their unique clinical features and specific pathophysiological mechanisms.17
Several studies indicate that the FAS−670G>A polymorphism may be associated with different autoimmune diseases. In particular, it has been shown that this SNP confers susceptibility to systemic lupus erythematosus,15 multiple sclerosis19 20 and autoimmune hepatitis.21 When rheumatoid arthritis22 23 and primary Sjögren’s syndrome (SS)24 25 are considered, conflicting results are available in the literature. Furthermore, no association between the FAS−670G>A polymorphism and susceptibility to other autoimmune conditions, such as coeliac disease,26 juvenile idiopathic arthritis,27 Hashimoto’s thyroiditis and Graves’ disease28, was found.
The lack of information concerning a possible association between the functional FAS−670G>A SNP and susceptibility to SSc prompted us to study a large cohort of white Italian SSc patients to evaluate the role of this SNP in influencing the susceptibility to the disease and its clinical subsets and in modulating both the autoantibody profile and severity of the disease.
PATIENTS AND METHODS
Patients and control subjects
We enrolled 350 SSc patients (316 women and 34 men, mean age 56.4 years (SD 12.5)) attending the outpatients clinics of the Divisions of Rheumatology of the universities of L’Aquila, Florence, Pisa, Pavia, the Catholic University of Rome and II University of Naples. Patients were representative of the whole Italian population, deriving from the northern, central and southern areas of Italy. The clinical subsets of lcSSc (259 patients) and dcSSc (91 patients) were defined according to the classification of LeRoy et al,1 which takes into account, along with the extent of skin thickening, the length of the interval between the appearance of Raynaud’s phenomenon and that of skin sclerosis, the timing and features of internal organ involvement, the autoantibody profile and the capillaroscopic pattern. The mean disease duration was 8.2 years (SD 5.3), calculated from time of the onset of the first non-Raynaud symptom of SSc. In order to assess associated SS, all patients were interviewed for symptoms of ocular and mouth dryness and parotid enlargement, and were investigated for anti-SSA and anti-SSB antibodies. As screened on the basis of their clinical histories, clinical features and laboratory tests, patients with symptoms overlapping with those of SS and other autoimmune and/or connective tissue diseases were excluded from the study.
Healthy controls were recruited from 232 volunteers comparable for age and gender with patients, and residing in the same geographical areas as the patients. Exclusion criteria for control subjects were familial histories of autoimmune diseases and/or chronic inflammatory diseases.
All subjects were white, unrelated to each other and gave informed consent to participate in the study. The protocol was approved by the local ethics committees at the six hospitals.
Autoantibody analysis
All patients were tested for circulating autoantibodies, such as antinuclear antibodies (ANA), anticentromere antibodies (ACA) and anti-topoisomerase I (anti-Scl-70) antibodies, following standard methods. Briefly, ANA were determined using indirect immunofluorescence and HEp-2 cells as antigen substrate (Antibodies Inc, Davis, California, USA). ACA were determined by their distinctive indirect immunofluorescence pattern on HEp-2 cells. Anti-Scl-70 antibodies were determined by passive immunodiffusion against calf thymus extract, using a commercially available kit (Inova Diagnostics, San Diego, California, USA).
Organ involvement evaluation
All patients underwent a complete assessment for organ involvement according to recent guidelines.29 Briefly, interstitial lung involvement was studied with pulmonary function tests (diffusing capacity for carbon monoxide (DLCO) and forced vital capacity) and high resolution computed tomography.30 Interstitial lung disease (ILD) was diagnosed when forced vital capacity and DLCO were 70% or less. In the presence of isolated DLCO reduction, patients were investigated for pulmonary arterial hypertension (PAH). PAH was defined as estimated peak right ventricular systolic pulmonary artery pressure greater than 40 mm Hg on echocardiography, or mean pulmonary artery pressure greater than 25 mm Hg at rest or greater than 30 mm Hg on exercise by right heart catheterisation.31
Scleroderma renal crisis (SRC) was defined according to the presence of new-onset accelerated phase hypertension with evidence of renal impairment, microangiopathic haemolysis or significant end-organ damage.
Skin involvement was evaluated according to the modified Rodnan skin score.32
Genotyping for the FAS−670G>A SNP by PCR restriction fragment length polymorphism assay
Peripheral venous blood from all subjects was obtained for genomic DNA extraction. Blood was collected in EDTA tubes and DNA was purified using a QIAamp DNA Blood Mini kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions.
FAS−670G>A SNP was genotyped by PCR followed by MvaI restriction enzyme digestion, as previously described.14 We used the previously reported primer sequences (forward primer 5′–CTACCTAAGAGCTATCTACCGTTC–3′, reverse primer 5′–GGCTGTCCATGTTGTGGCTGC–3′; GenBank accession no AY450925), which generate a 332-bp fragment containing the polymorphic site.14 PCR were carried out in a final volume of 25 μl containing 60 ng DNA, using the AccuPrime SuperMix II System (Invitrogen, Carlsbad, California, USA). The reaction mixtures were amplified in a Techne TC-512 thermocycler (Techne Inc, Cambridge, UK) for 35 cycles, with each cycle consisting of 30 s at 94°C (denaturation), 30 s at 58°C (annealing) and 1 minute at 68°C (extension). The cycling was preceded by a single denaturation of 2 minutes at 94°C and followed by a final extension of 5 minutes at 68°C. PCR products were checked on 1% agarose gel and a 15 μl aliquot of each sample was digested for 2 h at 37°C with 1 μl (10 units) of MvaI (M-Medical, Milan, Italy). The restricted products were separated by 3% low-melting-point agarose gel electrophoresis with ethidium bromide staining and visualised under ultraviolet transillumination. Two polymorphic alleles, FAS−670G (189 + 99 + 44-bp) and FAS−670A (233 + 99-bp), could be easily distinguished (fig 1). Control samples of predetermined genotypes and negative controls were included in each sample set, and more than 10% of the samples were randomly selected for confirmation. PCR restriction fragment length polymorphism results were analysed by two independent assessors blinded with regard to the sample classification and were 100% concordant (ie, in the case of discordant results, samples were genotyped again in order to avoid any errors in genotype assessment).
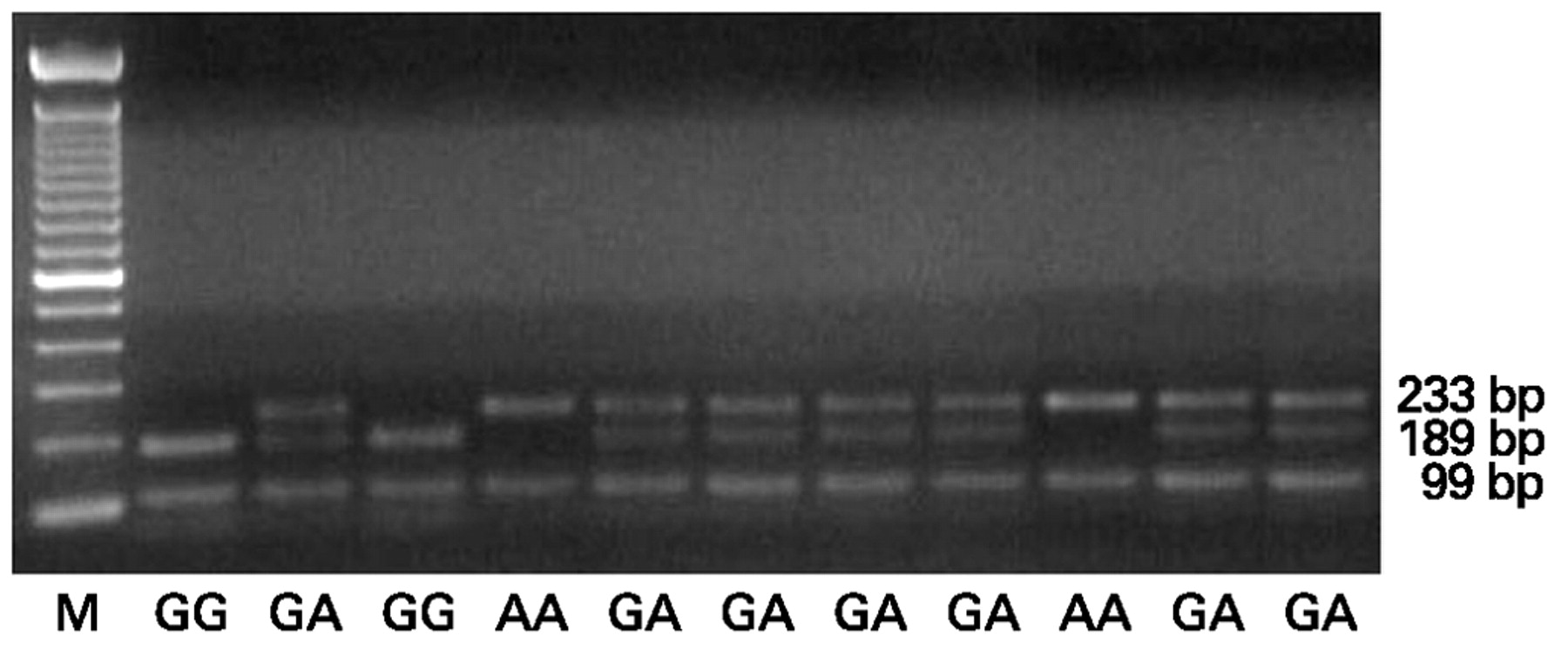
Agarose gel showing the specific PCR restriction fragment length polymorphism products of the FAS−670G>A polymorphism in DNA samples from different systemic sclerosis patients and control subjects. A 100 bp DNA marker (M) was used to distinguish alleles G (189 bp) and A (233 bp). AA, FAS−670AA genotype; GA, FAS−670GA genotype; GG, FAS−670GG genotype.
Measurements of sFAS serum levels by ELISA
Serum-circulating sFAS levels were analysed in 90 randomly selected SSc patients, subdivided into three groups according to their genotypes, as well as in 90 healthy controls. In this cohort, patients taking steroids and/or immunosuppressive medications discontinued therapy for at least one month before sample collection. No other drugs were discontinued, on the basis of their limited impact on immune system activity. The levels of sFAS were determined by ELISA using a commercially available kit (Cytimmune, College Park, Maryland, USA) according to the manufacturer’s instructions.
Statistical analysis
Statistical analysis was performed by using the statistical package for social sciences software for Windows and Prism software (Graph-Pad, San Diego, California, USA). Categorical and quantitative variables were respectively described as numbers, percentages (%) and mean (SD). The genotype distribution and allele frequencies in both patients and controls were compared by using χ2 analysis. A χ2 test was used to compare the observed numbers of each genotype with those expected for a population in Hardy–Weinberg equilibrium. Allele frequency of the FAS−670G>A polymorphism was obtained by direct count. The association between the FAS−670G>A polymorphism and SSc was assessed by using univariate analysis under a recessive genetic model of inheritance. The recessive genetic model compares the −670AA genotype with the combined −670GA+GG genotypes, which form the baseline group. Odds ratios (OR) with the 95% CI were determined. A sample size of at least 300 subjects per group was deemed sufficient to prove/exclude an association between the FAS−670G>A polymorphism and SSc with a statistical power (β) of 80%, and significance value of 0.05 (α). A sample size of approximately 300 subjects in each group would limit the β error of 0.10 (ie, the power of the study would be 80%). The Mann–Whitney U test was used to analyse differences in sFAS levels between subjects of different genotypes. Results are expressed as mean and range. p Values less than 0.05 were considered statistically significant.
RESULTS
Demographic, clinical and serological characteristics of the study population are reported in table 1.
FAS−670G>A polymorphism and SSc
No deviation from the expected population genotype proportions predicted by Hardy–Weinberg equilibrium was detected at the FAS−670 polymorphism site both in SSc patients and control subjects (table 2). A significant difference in genotype distribution for the FAS−670G>A polymorphism was observed between SSc and controls (χ2 = 13.45, p = 0.001; table 2). The frequency of the FAS−670A allele was significantly higher in SSc compared with controls (χ2 = 12.61, p = 0.001; table 2). No significant difference in genotype distribution and allele frequencies was observed between lcSSc and dcSSc, although a higher frequency of the FAS−670A allele was found in dcSSc (63% in dcSSc vs 57% in lcSSc, p = 0.2; table 2).
At univariate analysis, the FAS−670AA genotype significantly influenced the predisposition to SSc (OR 1.97, 95% CI 1.35 to 2.88, p = 0.001). In particular, we observed that the FAS−670AA genotype influenced the predisposition to both dcSSc and lcSSc (dcSSc vs controls OR 2.37, 95% CI 1.41 to 3.99, p = 0.001; lcSSc vs controls OR 1.84, 95% CI 1.23 to 2.75, p = 0.003).
FAS−670G>A polymorphism and correlation with ACA and anti-Scl-70 antibodies in SSc
This study confirms, in a large cohort of Italian SSc patients, the association between the two SSc clinical subsets and specific circulating autoantibodies (table 1). A significant association of ACA with lcSSc and anti-Scl-70 antibodies with dcSSc was found (p<0.001; table 1).
We evaluated the relationship between the FAS−670G>A polymorphism and the autoantibody profile in the whole SSc cohort, and we observed no significant difference in genotype distribution and allele frequencies either between ACA-positive and ACA-negative patients, or between anti-Scl-70 antibody-positive and anti-Scl-70 antibody-negative patients (table 3). Moreover, in both lcSSc and dcSSc, the FAS−670G>A polymorphism was not associated with the autoantibody pattern, although the prevalence of the FAS−670A allele was higher in anti-Scl-70 antibody-positive compared with anti-Scl-70 antibody-negative dcSSc (65% vs 60%; χ2 = 0.28, p = 0.9) and in ACA-negative compared with ACA-positive dcSSc (64% vs 57%; χ2 = 2.47, p = 0.3).
FAS−670G>A polymorphism in SSc according to organ involvement
A significantly greater incidence of ILD (62.6% of dcSSc patients compared with 41.7% of lcSSc patients; χ2 = 11.85, p = 0.001) and SRC (14.3% of dcSSc patients compared with 6.2% of lcSSc patients; χ2 = 5.83, p = 0.01) was observed in dcSSc in comparison with lcSSc (table 1).
No association between the FAS−670G>A polymorphism and internal organ involvement (ILD, PAH and SRC) was found (data not shown). No significant difference in genotype distribution and allele frequencies was observed between dcSSc patients with ILD and those without ILD, although a higher frequency of the FAS−670A allele was found in ILD dcSSc (67% vs 57%; χ2 = 1.59, p = 0.2). Moreover, we did not find any association between the FAS−670AA genotype and ILD (OR 2.16, 95% CI 0.87 to 5.33, p = 0.09). The FAS−670A allele frequency was higher, although not significantly, in ILD anti-Scl-70 antibody-negative in comparison with no-ILD dcSSc patients (75% vs 50%, respectively).
FAS−670G>A polymorphism and sFAS levels
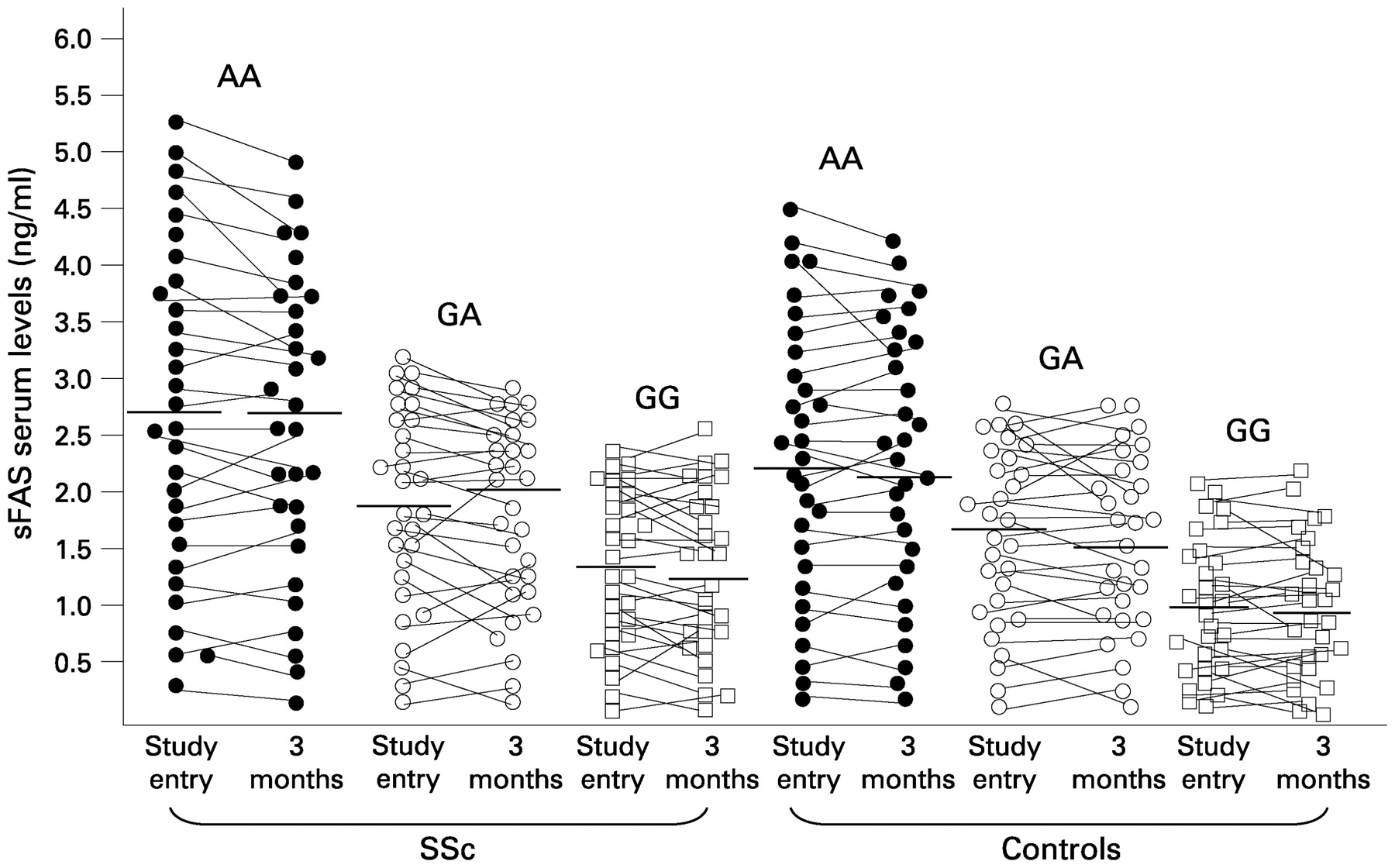
In order to evaluate the role of the FAS−670G>A polymorphism in affecting the levels of sFAS, we analysed, at study entry and after 3 months, serum-circulating sFAS in 30 patients carrying the FAS−670GG genotype, 30 patients carrying the FAS−670GA genotype and 30 patients carrying the FAS−670AA genotype. These groups were comparable for age and gender. The mean sFAS concentration was higher in patients with the FAS−670AA genotype than in those carrying the FAS−670GG genotype (2.60, 5.4–0.3 ng/ml vs 1.30, 2.40–0.1 ng/ml, p = 0.003). The mean sFAS concentration of patients carrying the FAS−670GA heterozygous genotype corresponded to an intermediate level (1.70, 3.20–0.3 ng/ml, p not significant vs the other two groups). After 3 months the results mirrored what was observed in the first experiments, with only few individual modifications (FAS−670AA genotype vs FAS−670GG genotype 2.60, 5.20–0.20 ng/ml vs 1.20, 2.50–0.10 ng/ml, p = 0.003; FAS−670GA genotype 1.85, 3.00–0.20 ng/ml, p not significant vs the other two groups; fig 2). Similar results were observed in the control cohort subdivided into three groups according to FAS−670 genotypes (fig 2).
{kind=link}
{kind=link}
Serum levels of sFAS in systemic sclerosis (SSc) patients and healthy control subjects according to FAS−670 genotypes, at study entry and after 3 months. Horizontal bars indicate the mean sFAS concentration of the corresponding genotypes (AA, GA, GG). In both SSc patients and controls, differences between carriers of the FAS−670AA and FAS−670GG genotypes were statistically significant and did not change after 3 months (p = 0.003 in SSc group, p = 0.004 in control group, by Mann–Whitney U test).
DISCUSSION
In this study, we demonstrate an association between the FAS−670G>A polymorphism and both lcSSc and dcSSc clinical subsets of SSc, thus identifying FAS as a novel susceptibility gene for this heterogeneous and complex disease.
There is increasing evidence to suggest a complex genetic involvement in the pathogenesis of SSc. Candidate genes for SSc include a number of genes encoding proteins involved in fibrosis, vasculopathy and immune responses, thus reflecting the three main areas of disease expression.33 34 Multiple genes are thought to influence the susceptibility to SSc and to modulate disease onset and/or the clinical course, in combination with environmental factors. Furthermore, several SNP or gene variations have been associated with SSc, disease subtypes and/or specific autoantibodies.33 34
Our results confirm that functional SNP in genes coding for immunoregulatory molecules, such as the FAS antigen, may influence the genetic susceptibility to autoimmunity, as already reported in systemic lupus erythematosus,15 multiple sclerosis,19 20 autoimmune hepatitis,21 rheumatoid arthritis22 and primary SS.24
Several well-recognised associations between specific autoantibodies and the clinical features of SSc are known,1 35 and our study confirms the association between specific circulating autoantibodies and disease subsets. Nevertheless, the distribution of anti-Scl-70 antibodies and ACA in our patients deserves to be discussed; anti-Scl-70 antibody positivity has long been known to occur in patients with lcSSc,36–39 as we found in the present study. Moreover, Italian patients are known to present a higher percentage of anti-Scl-70 antibody positivity.40 On the other hand, although we found a higher-than-expected positivity, ACA have previously been detected in patients with dcSSc.38 39 41
To date, little is known about the cellular and molecular mechanisms leading to autoantibody production and how autoimmunity is linked to organ involvement and damage during SSc. FAS-mediated apoptosis plays a crucial immunoregulatory role through the deletion of autoreactive B and T cells during the normal development of the immune system.10 42 43 Therefore, a dysregulation of FAS-mediated signalling might contribute to the pathogenesis of SSc via its biological effects on cell survival, differentiation, proliferation and cell death on the one hand and/or causing organ-specific damage on the other. In fact, SSc is characterised by sustained activation and clonal expansion of B and T lymphocytes, with the production of specific autoantibodies and profibrotic cytokines.2–8 Different subsets of circulating T cells from SSc patients have been shown to be resistant to FAS-mediated apoptosis.9 In addition, a decreased susceptibility to FAS-induced apoptosis has also been reported in SSc dermal fibroblasts, suggesting that abnormal apoptotic regulation may directly contribute to the pathogenesis of progressive fibrosis during SSc.44 45 Moreover, it has recently been shown that FAS and FAS ligand-deficient mice are resistant to bleomycin-induced scleroderma, further arguing for a critical role of the FAS/FAS ligand pathway in the development of fibrosis.46 Taken together, these data suggest that apoptosis disturbances and dysregulated FAS-mediated signalling might play a role in the pathogenesis of SSc through the genetic control of apoptosis resistance in different cell types, such as activated lymphocytes and fibroblasts.
Furthermore, a recent study showed that the FAS−670G>A polymorphism correlates with the serum-circulating levels of sFAS in normal individuals, and that subjects carrying the FAS−670AA genotype have significantly higher sFAS levels than those carrying the FAS−670GG genotype.16 Serum levels of sFAS are increased in SSc patients in comparison with healthy individuals,9 47 and our data show a significant association between the FAS−670 genotype and the levels of sFAS in the sera of SSc patients, as well as in normal individuals. Increased levels of sFAS have been described in several autoimmune diseases, and mice injected with sFAS display autoimmune features.48 49 sFAS downregulates the number of membrane-associated FAS receptors and binds to and neutralises FAS ligand, thereby functionally antagonising the FAS/FAS ligand proapoptotic pathway.49 50 Nevertheless, it must be pointed out that apoptosis is a very complex cellular mechanism in which related molecules are likely to be controlled not only by the genetic background, but also by several pathways in order to ensure physiological cell survival. In this light, some transcriptomic studies will be necessary for a full understanding of the functional impact of genotypes.
In addition, as already reported in other autoimmune conditions, we did not find any significant association between the FAS−670G>A polymorphism and the autoantibody profile or the severity of the disease.20 22 27 In particular, we further investigated the possible association between the FAS−670G>A polymorphism and ILD, which is the most frequent cause of death among SSc patients30 and did not find any association. However, we cannot exclude the possibility that a linkage between the FAS−670G>A SNP and other different functional variants in this and/or other genes affecting cell apoptosis might contribute to determine specific SSc phenotypes within the disease. Further studies are needed to address this important aspect.
In summary, the present case–control study performed on a large cohort of white Italian SSc patients provides the first evidence that the FAS−670A allele significantly influences the susceptibility to SSc. Our data suggest that a genetic control of apoptotic processes might contribute to both persistent immunological and fibrogenic abnormalities in the course of SSc. In addition, these data further strengthen the classification of SSc within the spectrum of autoimmune diseases via shared inherent immunoregulatory defects. Further studies are warranted to elucidate the functional role of this SNP in the pathogenesis of SSc. Further research on other polymorphic genes that influence apoptosis, such as FAS-ligand, bcl-2 and bcl-x, might contribute to a better understanding of the genetic context in which the FAS−670G>A SNP influences the susceptibility to SSc and the role of a genetic control of apoptosis in the disease process.
Acknowledgments
The authors would like to thank Mrs Federica Sensini for excellent technical assistance.
REFERENCES
Footnotes
Competing interests: None.
Funding: This study was supported by the Ministero dell’Istruzionze, dell’Università e della Ricerca, Programmi di ricerca di Rilevante Interesse Nazionale (PRIN 2004/2006, grant 2004064281_002).
Ethics approval: The protocol was approved by the local ethics committees at the six hospitals.
Patient consent: Obtained.