Article Text

Abstract
OBJECTIVE Cells of the myeloid lineage comprise a very heterogeneous population with many phenotypes and functional activities including macrophages and dendritic cells. To investigate the status, differentiative potential and lineage commitment of monocytic cells in systemic lupus erythematosus (SLE) patients, this study isolated and cultured peripheral blood monocytes from patients and healthy donors.
METHODS Monocytes were isolated by gradient centrifugation and adherence to plastic dishes. The cells were then cultured for three days, partially supplemented with GM-CSF and interleukin 4 (IL4) to obtain dendritic cells. The differentiation status was monitored by the expression of surface markers using flow cytometry and cytokine secretion.
RESULTS Monocytes from SLE patients expressed significantly lower numbers of the monocytic marker CD14 and HLA-DR while secreting significantly more tumour necrosis factor α (TNFα) than monocytes from healthy donors. The addition of GM-CSF and IL4 resulted in an inhibition of TNFα secretion, but was not sufficient to generate monocytederived dendritic cells.
CONCLUSION Monocytes from SLE patients are severely altered in phenotype and function and have a limited differentiation flexibility towards the accessory type of monocytic cells.
- systemic lupus erythematosus
- monocytes
- differentiation
- tumour necrosis factor α
Statistics from Altmetric.com
Systemic lupus erythematosus (SLE) is characterised by the presence of antinuclear antibodies and anti-cytoplasmatic proteins.1 ,2 Immune complexes formed by these antibodies and their specific antigens contribute to multiple organ manifestations. The induction of autoantibody production, however, remains unknown and the role of antigen presentation in the pathogenesis of the disease has not been studied in greater detail so far although T lymphocytes contribute to the production of autoantibodies.3 An aberrant antigen presentation may initially result in an insufficient control of B cells by the activated T lymphocytes.
A reduced functional activity, decreased levels of HLA-DR+monocytes, and increased levels of soluble CD14 (sCD14) have been observed.4-6 Further studies have shown that the expression of CD54 (ICAM-1) and CD64 (FcgRI) was increased, whereas the expression of CD32 (FcgRII) was decreased on monocytes from SLE patients.7 ,8 Additionally, peripheral blood mononuclear cells (PBMC) from SLE patients were found to secrete higher levels of interleukin 1 (IL1), interleukin 6 (IL6) and interleukin 10 (IL10)9 ,10 compared with cells from healthy donors. This cytokine milieu may well increase or even trigger the immunoglobulin production of B cells. Spleen dendritic cells (DC) were first described some 20 years ago and are the most potent T cell stimulators.11 Since 1987, increasing evidence has been obtained that peripheral blood monocytes (PBM) may differentiate spontaneously into CD1a+/CD14low, “veiled”, monocyte derived accessory cells (MoAC).12-14 According to their phenotype and function, these cells represent intermediate and indeterminate precursors of DC.14 The combined action of GM-CSF and interleukin 4 (IL4) on monocytes was first demonstrated by the differentiation of highly potent CD1+ APCs specific for mycobacteria.15 Using this combination of cytokines, DC may be differentiated from monocytes (monocyte derived dendritic cells, MoDC) isolated by adherence, or magnetic separation.14 ,16 ,17
In this study, we isolated PBM from patients with SLE by plastic adherence and analysed the spontaneous differentiation towards MoAC as a prerequisite for obtaining MoDC. Additionally, we used GM-CSF and IL4 to generate MoDC.16 Monocytes from patients with active SLE were found to be severely altered, as documented by the expression of significantly lower amounts of monocytic markers compared with PBM from healthy donors. Moreover, these SLE monocytes produced significantly more tumour necrosis factor α (TNFα) than PBM from healthy donors. Spontaneously, the SLE monocytes did not differentiate into MoAC, but rather recovered from an activated state to a phenotype of normal monocytes. After the addition of GM-CSF and IL4, they did not differentiate towards DC. These findings indicate disturbances in the antigen presenting cells initiating the autoreactive cascade in SLE.
Methods
PATIENTS
Venous blood samples were obtained from 23 patients (18 women and 5 men) with SLE (age range 20–60 years) and from 20 normal donors from the blood bank as controls (age range 25–55 years). All patients fulfilled at least four of the 1982 revised ACR classification criteria for SLE.18 Disease activity was determined according to definitions recently described.19 Active SLE: 12 patients with manifestations of clinical activity (arthritis, vasculitis, myositis, serositis, fever and others) accompanied by a significant increase (negative/positive or fourfold) in anti-DNA antibodies, a significant decrease (below physiological boarder level) in C3 or C4 level, and with or without other laboratory abnormalities (proteinuria, leucopenia and other) that usually required restarting or increasing corticosteroids or immunosuppressive drugs. Inactive SLE: 11 patients without signs of clinical and immunological/laboratory activity in the previous three months and without necessity to increase dose of corticosteroids and immunosuppressive drugs. Here, the dose of prednisolone did not exceed 10 mg per day. The ECLAM score for disease activity20 ranged from 1–7, and 21 patients received prednisolone or equivalent corticosteroids at 5–75 mg/day.
MONOCYTE ISOLATION, CELL CULTURE AND FLOW CYTOMETRY
PBMC were obtained from 20–30 ml of citric blood from patients with SLE or from buffy coats from healthy donors. Isolation and cultivation of PBM was performed as described.16 To estimate the role of corticosteroids, we added 0.5μM or 5 μM methylprednisolone (MP). To obtain the differentiation of DC, 200 U/ml (40 ng/ml) recombinant human (rhu) GM-CSF (PBH, Hannover, Germany) and 50 U/ml (0.5 ng/ml) rhuIL4 (PBH, Hannover, Germany) were added as described.16
Surface antigens of monocytic cells were stained and analysed as described,21 using fluorochrome labelled (FITC/PE) antibodies directed against CD1a (Dianova, Hamburg, Germany), CD3 (Dako, Hamburg, Germany), CD14 (Dianova), CD19 (Dako), CD33 (Dako), CD66b (Dianova), CD71 (Dako), or HLA-DR (Dako). Flow cytometry was performed on a FACScan (Becton Dickenson, Heidelberg, Germany), using Cell-Quest software and acquiring at least 10 000 events in total and 2000 events in the monocyte gate. Monocytic cell populations were identified on a scattergram gate as described before, including control labelling of monocytes in peripheral blood and using isotype matched control antibodies MOPC21 and UPC-10 (Sigma).21
ELISAS FOR TNFα AND IL6
Supernatants of monocyte cultures were analysed in a sandwich ELISA for the presence of TNFα, using matched pairs of monoclonal mouse antihuman TNFα (Pharmingen, Hamburg, Germany) using standard methods. Briefly, ELISA plates (Nunc, Wiesbaden, Germany) were coated with primary antibody (2 μg/ml), diluted in 0.05 M carbonate buffer pH 9.6 (50 μl/well) and incubated overnight at 4°C. After washing the plates three times with phosphate buffered saline (PBS) (200 μl/well) they were blocked with PBS/10% FCS (200 μl/well) for one hour at 37°C, and washed three times with PBS/0.05% Tween 20. Subsequently 100 μl/well test samples and standard protein were added and incubated overnight at 4°C. The plates were washed seven times with PBS/0.05% Tween 20. After adding 100 μl/well biotinylated monoclonal mouse antihuman cytokine antibody (2 μg/ml in PBS/10% FCS), plates were incubated for one hour at 37°C and washed eight times with PBS/0.05% Tween 20. One hundred μl/well of peroxidase conjugated streptavidin (Dianova, Hamburg, Germany) were added to the plates, which were then incubated for 30 minutes at room temperature. Afterwards, they were washed eight times with PBS/0.05% Tween 20. One hundred μl/well TMB-substrate (Sigma, Deisenhofen, Germany) were added to the plates. Colour development was stopped with 50 μl 1 M H2SO4. Optical densities were read at 450 nm using an ELISA reader (Anthos, Cologne, Germany). Detection limit was 50 pg/ml.
STATISTICS
Statistical analysis was perfomed using the Instat software package. To evaluate the statistical significance of flow cytometric data we used the percentage of positive cells and geometric mean of fluorescence intensity (marker expression, measured in log scale) provided by CellQuest. Medians were compared using the unpaired Mann-Whitney U test. Likewise, we compared the medians of cytokine secretion (ELISA).
Results
PHENOTYPE AND SPONTANEOUS DIFFERENTIATION OF MONOCYTES
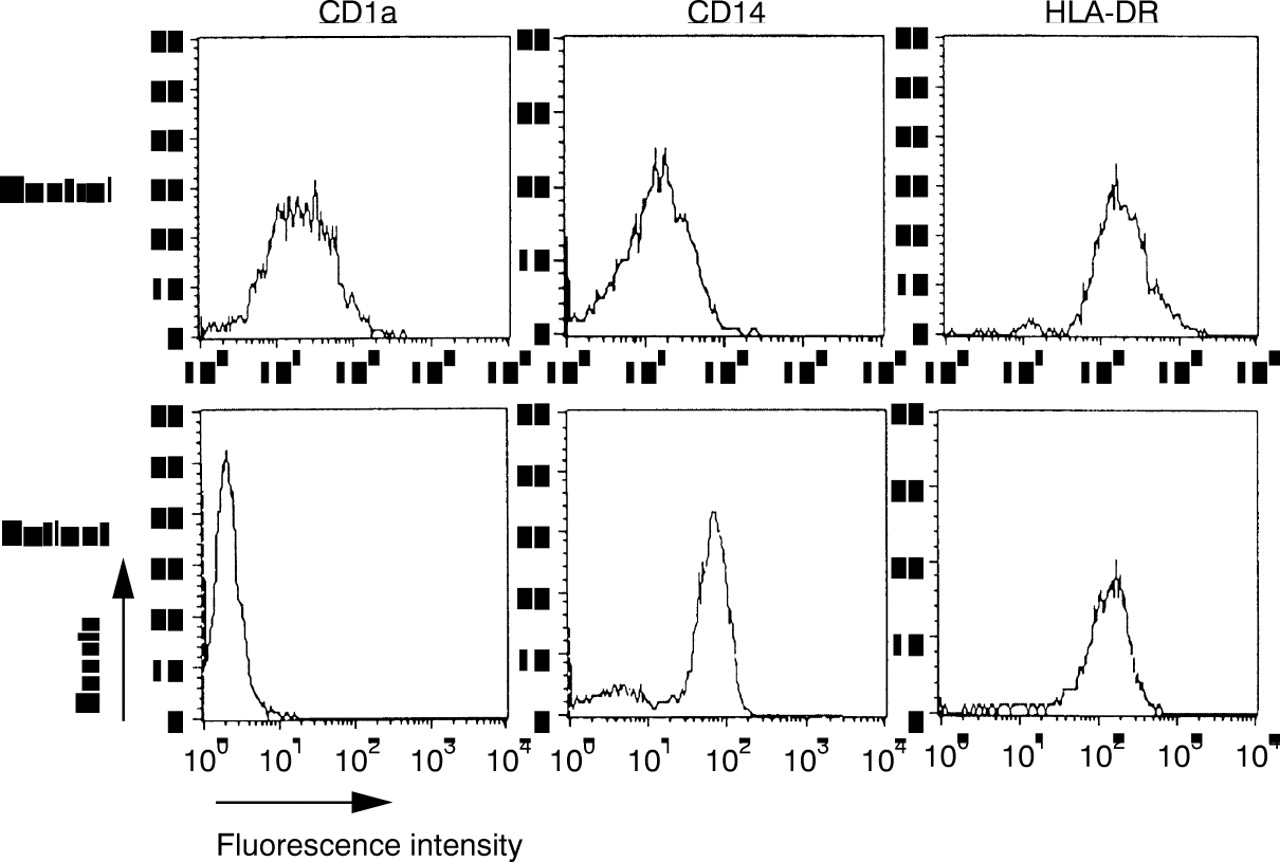
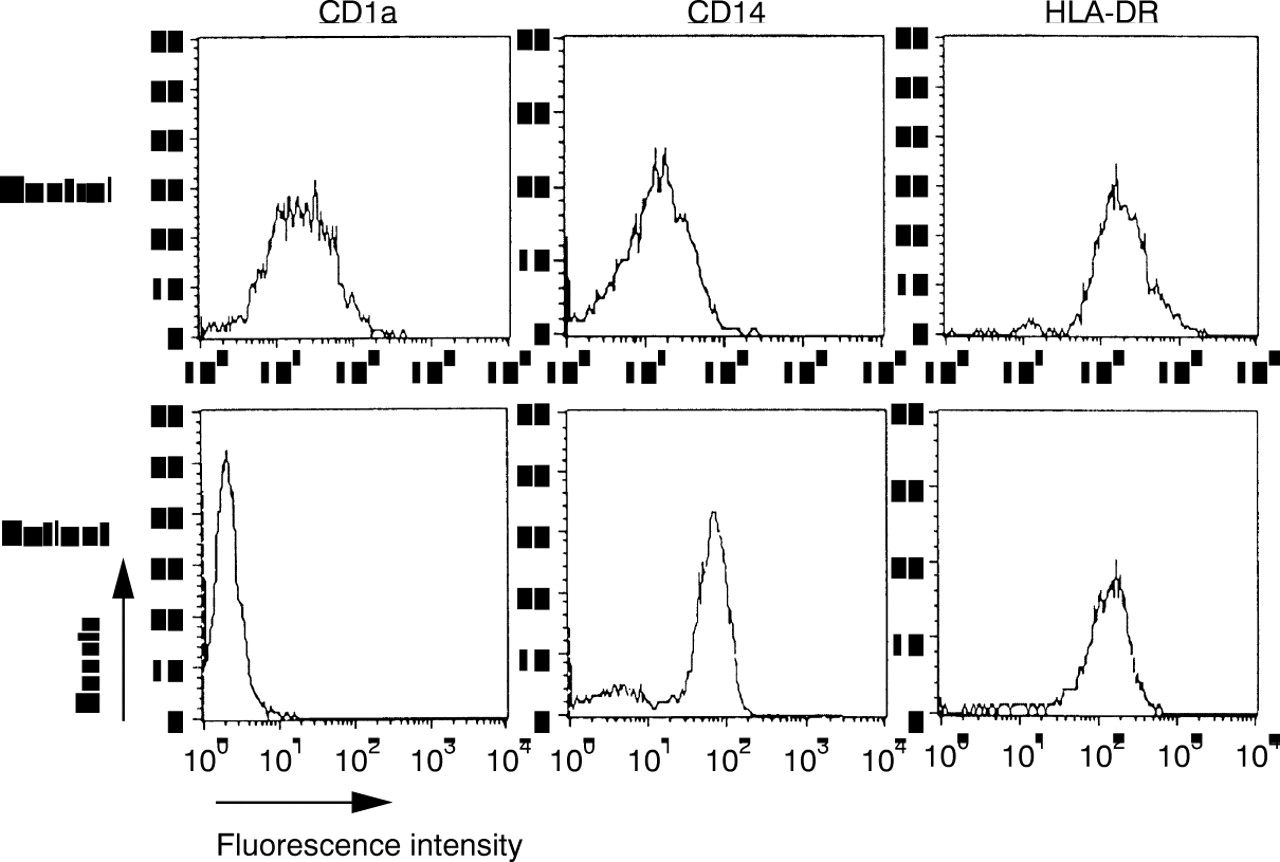
Monocytes from healthy donors (n=8) were analysed by flow cytometry and compared with PBM from SLE patients (n=17). PBM from healthy donors expressed the cell surface antigens CD14 and HLA-DR, as shown in figure 1 (control row). In contrast, PBM from SLE patients expressed significantly less CD14 or HLA-DR (table 1), with populations not expressing these markers at all. In four cases less than 20% of the cells, defined by scattergram analysis, expressed the LPS coreceptor CD14. Moreover, the amount of CD14 on positive monocytes was significantly suppressed (table 1). A direct comparison of the two myelomonocytic markers CD14 and CD33 (example in fig 1, patient 3) showed in most cases more CD33+ cells than CD14+ cells, indicating a selective loss of CD14. Additionally, the expression of CD71 was detected on freshly isolated monocytes from SLE patients (5 of 17), but never on monocytes from healthy donors. To exclude the possibility of missing SLE monocytes because of a shift of cell size or granularity, the scattergram was analysed as a whole for the existence of CD14+ monocytes, but no monocyte population of unusual size or granularity could be observed (data not shown). Controls were performed with αCD3, αCD19 and αCD66b to exclude the presence of T cells, B cells and neutrophils, respectively. These cells were not present in the monocyte gate to an extent greater than 15%. Control labellings of SLE monocytes in peripheral blood also demonstrated that this phenotype was not attributable to the isolation procedure via Ficoll. Likewise, control labellings from healthy volunteers demonstrated that the differences observed were not attributable to the buffy coat preparation (data not shown).

Representative examples of surface markers on monocytes from three patients with active SLE. Monocytes from healthy donors (control) showed an almost homogenous staining for CD14 (strong) and HLA-DR (weak). Monocytes from SLE patients showed various amounts of positive cells. In many cases, more cells were HLA-DR+than CD14+ (see patient 1), but the expression of the two markers did not correlate. Cells expressed myeloid markers such as CD33, but the activation marker CD71 was also detected. The cells were analysed after scattergram gating of monocytic cells. Controls and patients did not react with isotype controls of irrelevant specificities. Control labelling revealed <15% of contaminating lymphocytes or neutrophils.
Statistics of monocyte analysis from SLE patients and healthy controls
PBM from SLE patients (n=12) upregulated the expression of both CD14 and of HLA-DR within 72 hours in vitro, now representing the phenotype of freshly isolated PBM from healthy donors (not shown). The percentage of CD14+ cells was increased whereas MHC class II expression was slightly reduced on monocytic cells from SLE patients (table 1). The Langerhans cell marker CD1a that was expressed on MoAC from healthy donors13 was only rarely seen on monocytic cells from SLE patients (table 1).
In this context, we analysed the effects of corticosteroids on the phenotype and differentiation of monocytes. MP was added to monocyte cultures from healthy donors at final concentrations of 0.5 or 5μM respectively. No change, however, in CD14 or HLA-DR expression was observed as a result of the addition of MP (not shown).
SECRETION OF TNFα AND EFFECTS OF GM-CSF AND IL4
Culture supernatants were tested for the presence of TNFα, which has been described as an important mediator of monocyte activity and possibly changed in SLE patients.9 ,22 As figure 2 shows, monocytes from SLE patients produced 10-fold more TNFα than monocytes from healthy donors (table 1).

Cytokine secretion by monocytes from healthy donors (control) and SLE patients. Cytokine secretion was measured 24 hours after cell isolation. One group of SLE monocytes was supplemented with GM-CSF and IL4. Individual data and medians are presented.
Finally, it was analysed, if this restricted capacity of differentiation could be overcome by culturing PBMs of SLE in patients in the presence of GM-CSF and IL4. After three days of culture, PBMs from healthy donors expressed CD1a, CD14 was down regulated, while HLA-DR was up regulated (fig 3). In contrast, only few PBM from SLE patients expressed CD1a, the expression of CD14 was significantly increased and HLA-DR was not modulated to the extend of healthy controls (fig 3, table 1). Accessory markers such as CD40 or CD80 were not up regulated during culture (data not shown). Investigating the production of TNFα, it became evident that this combination of cytokines suppressed the production of TNFα significantly (p=0.036), but not to the level of monocytes from healthy donors (table 1; fig2).
{kind=link}
{kind=link}
{kind=link}
Differentiation of monocytes after treatment with GM-CSF and IL4. At day 3, monocytes from healthy donors were CD1a+/CD14low/HLA-DR++, therefore representing monocyte derived dendritic cells. In contrast, monocytes from SLE patients were CD1a-/CD14+/HLA-DR++. This patient was representative for the six patients analysed. The differences in CD1 and CD14 expression were highly significant (both p=0.0007). Flow cytometric analysis was performed as in figure 1, isotype controls not shown.
As the study was designed to allow retrospect comparisons of alterations in vitro with clinical course of the disease, we compared the results in monocyte biology with clinical parameters, such as anti-DNA-Ab titres, C3/C4 levels, leucocyte counts or corticosteroid dose. None of these parameters, however, indicated any (positive or negative) correlation, nor was it possible to find a statistically significant difference between parameters of active and inactive patients. The statistical inability to dissect active and inactive patients was attributable to the clinical and laboratory variability of some patients in both groups. Some data, however, conferred the impression that alterations were most severe in acutely diseased patients.
Discussion
Antigen presenting cells, and especially DC are of growing interest for understanding regulatory mechanisms of the immune response. Here, we investigated freshly isolated monocytes from SLE patients with special emphasis on their potential to differentiate towards accessory cells/dendritic cells.
Surprisingly, monocytes were severely altered and subpopulations did not even express CD14 and MHC class II molecules. These results are in accordance with earlier investigations showing increased levels of sCD14 and decreased numbers of HLA-DR+monocytes.5 ,6 Neither CD14 nor HLA-DR expression correlated to the disease activity. As the monocytes were enriched by more than 85% as monitored by Giemsa staining and flow cytometry, a contamination by other cell types can be excluded. It needs to be emphasised that methods using the expression of CD45 and CD14 for the classification of monocytes are not suitable to detect such changes.21 The monocytes from SLE patients up regulated CD14 and HLA-DR within 72 hours, thus reconverting into normal PBM. By expressing markers such as CD71 (transferrin receptor), they showed a parameter of activation associated with macrophage populations, thus representing the antipodes of accessory cells.14Accordingly, monocytes from SLE patients did not spontaneously differentiate into accessory cells. Medication was shown to be a most unlikely candidate to explain the changes. The tested concentrations of MP did not show any effect on the differentiation and the two patients not receiving corticosteroids also exhibited the altered phenotype of monocytes. In patients receiving corticosteroids, no correlation was observed between the dose of corticosteroid application and the intensity of marker expression. This is in accordance with the data of Shirikawa et al 5 finding no correlation of the reduced MHC class II levels and the application of corticosteroids. Inhibitory effects of corticosteroids, however, were reported both for macrophages and DC in vitro referring to an impaired function, for example, in T cell stimulation.23 ,24
The inflammatory cytokine TNFα was strongly overexpressed. Regarding the biology of TNFα, it is probable that this disturbance in cytokine production was linked to the alterations in phenotype and differentiation seen under spontaneous culture conditions. Further analysis, however, will be required to determine if all the TNFα produced in disease status is biologically active. It is also possible that the increased levels of TNFα production are counterbalanced by an increased sTNFR production or antagonistic cytokines in vivo. Some patients had decreased levels of IL6 (data not shown). As IL6 has been attributed to accessory activity of myeloid cells and differentiation towards DC,14 its low expression in these cases might further help explaining the changes observed in monocyte biology.
The limited differentiation flexibility of monocytes from SLE patients was underlined by their inability to respond to GM-CSF and IL4 in order to generate monocyte derived DC.14-16 Here, CD14 was not modulated (compared with SLE-MoAC), nor was an expression of CD1a seen. MHC class II was up regulated but not quite to the extent observed on MoDC from healthy donors. Additionally, these altered cells from patients did not fulfil any other requirements of DC, like non-adherence, or a stimulatory capacity for T cells. TNFα was down regulated under the aegis of GM-CSF and IL4, but not to the level of healthy controls. Further studies will have to investigate the extent to which increased amounts of one or two of the cytokines might improve the differentiation of DC or which cytokines or anti-cytokines are required in addition.
In summary, we detected severe alterations in the biology of monocytes from patients suffering from SLE. Our data indicate an imbalance of stimuli in vivo that may be monitored ex vivo. As of yet, it remains to be investigated whether these alterations are a result of the systemic inflammation in SLE or represent intrinsic features of a pathogenic impact of the disease itself. Although tolerance induction has been studied well in recent years and professional antigen presenting cells such as DC might induce tolerance,25 the role of an altered antigen presentation remains uncertain. An imbalance in monocyte/DC biology and an inability to generate the appropriate T cells help, may contribute to the initial induction of autoimmunity, even in antibody/B cell mediated diseases such as SLE. Future investigations of monocytes and other accessory cells may help to resolve the enigmatic induction of autoimmunity in SLE.
Acknowledgments
The authors thank Dr S Bläβ and Mrs Y Sommer for critical reading of the manuscript.
References
Footnotes
Funding: the work was supported by grants of the Deutsche Forschungsgemeinschaft (DFG) Hi 620/1–1 and by institutional grants from the Charité University Hospital.