Article Text

Abstract
Background: Raised interleukin (IL)6 and IL10 levels are thought to contribute to the pathogenesis of systemic lupus erythematosus (SLE) by enhancing autoantibody production and immune complex (IC) formation. These immune complexes can then stimulate cellular reactions through Fc and complement receptors.
Objective: To investigate whether circulating SLE ICs stimulate type 2 cytokine production.
Methods: Twenty serum samples from patients with active SLE were compared with sera from 18 healthy controls. Sera and polyethylene glycol (PEG) precipitates from sera were added to peripheral blood mononuclear cell (PBMC) cultures, and the production of IL10 and IL6 was investigated by enzyme linked immunospot assay (ELISPOT) and enzyme linked immunosorbent assay (ELISA). Fc gamma receptor (FcγR) antibodies were used in blocking experiments, and flow cytometry was used to assess the correlation between monocyte FcγR expression and IC-induced cytokine production.
Results: Ten per cent dilutions of the SLE sera induced a significantly increased number of IL10-producing cells in comparison with control sera (median, 11.75 v 1.25 spot forming cells/50 000 PBMC; p<0.0001). PEG precipitates from SLE sera also induced significantly increased levels of IL10 (p=0.016) and IL6 (p=0.042) in comparison with control PEG precipitates. IL10 production induced by SLE PEG precipitates or by artificial ICs could be blocked by anti-FcγRII antibodies, and the FcγRII expression on CD14+ monocytes correlated with the IC-induced production of IL10 and IL6.
Conclusions: SLE sera stimulate IL10 and IL6 production from PBMC, and this effect is at least partly explained by precipitable ICs acting through FcγRII. This effect provides a possible mechanism for the enhanced production of IL10 in SLE, whereby B cell activation, antibody production, IC stimulated monocytes/macrophages, and type 2 cytokines create a vicious cycle that may help to maintain B cell hyperactivity in SLE.
- systemic lupus erythematosus
- immune complexes
- interleukin 10
- FcγRII
- ELISA, enzyme linked immunosorbent assay
- ELISPOT, enzyme linked immunospot
- FcγR, Fc gamma receptor
- FCS, fetal calf serum
- HAGG, heat aggregated immunoglobulins
- IC, immune complex
- IFN, interferon
- IL, interleukin
- PBMC, peripheral blood mononuclear cells
- PBS, phosphate buffered saline
- PEG, polyethylene glycol
- SFC, spot forming cells
- SLE, systemic lupus erythematosus
Statistics from Altmetric.com
- ELISA, enzyme linked immunosorbent assay
- ELISPOT, enzyme linked immunospot
- FcγR, Fc gamma receptor
- FCS, fetal calf serum
- HAGG, heat aggregated immunoglobulins
- IC, immune complex
- IFN, interferon
- IL, interleukin
- PBMC, peripheral blood mononuclear cells
- PBS, phosphate buffered saline
- PEG, polyethylene glycol
- SFC, spot forming cells
- SLE, systemic lupus erythematosus
Systemic lupus erythematosus (SLE) is a systemic disease of presumed autoimmune aetiology, that is characterised by polyclonal B cell activation. In this disease, an increase in antibody production with a preferential increase in autoantibodies, together with defective clearing of immune complexes (ICs), gives rise to increased levels of ICs. This contributes to the disease manifestations associated with deposition of ICs in kidney, skin, pleura, and other organs.
Antibody production by B lymphocytes is stimulated by type 2 cytokines—for example, interleukin (IL)4, IL6, IL10, and IL13. Increased levels of IL101,2 and IL63,4 have been demonstrated in SLE, and both cytokines have been shown to be of importance for the production of disease associated autoantibodies recognising double stranded DNA.5,6 A number of studies have also reported a correlation between SLE disease activity and IL10, either measured as serum levels7,8 or as ratios between IL10 and interferon γ (IFNγ) or IL2 measured in different in vitro systems.9,10
ICs have a significant role in immune responses by inducing effector functions that are important in inflammatory responses and elimination of complexed antigens. They exert their physiological effects through stimulation and cross ligation of cellular receptors expressed on macrophages and neutrophils, thereby stimulating cells to phagocytosis and cellular activation.
In animal models ICs can stimulate the production of IL10 in vivo, an effect that has been shown to have either beneficial or detrimental effects on disease manifestations depending on the experimental system studied. In IC-induced lung disease in rats in vivo the IL10 produced ameliorates the inflammatory disease process.11 On the other hand, in mice sublethally infected by Listeria monocytogenes, injection of ICs caused the mice to succumb, an effect of macrophage suppression resulting from IC-induced IL10 production.12 Thus, IC-induced IL10 production has been shown to exert important physiological effects in vivo.
Artificial ICs have recently been shown to induce production of IL6 and IL10 from human macrophages, thus shifting the immune response from a type 1 to a type 2 pattern.13–15 The possibility therefore exists that SLE ICs obtained in vivo also might exhibit type 2 cytokine inducing effects by stimulating macrophage production of IL10 and IL6. These cytokines might then further stimulate antibody production and subsequent IC formation.
In this study we therefore considered the possibility that circulating ICs act by stimulating type 2 cytokine responses. We show that this is the case and that the effect is at least partially mediated through Fc gamma receptor II (FcγRII), an IC-binding receptor which has been implicated in SLE pathogenesis.
PATIENTS AND METHODS
Patients
Twenty patients (19 women and one man; mean age 48 years, range, 25–78) fulfilling the 1982 criteria for the diagnosis of SLE16 were incorporated into the study. Serum samples were collected during an earlier study on patients with active SLE,17 in which many of the patients had IC associated anti-C1q antibodies.18 For FcγR blockade experiments we used three SLE sera showing low functional activity of the classical complement pathway (33%, 35%, and 29%; reference range 80–120%19) as a sign of IC-induced activation of the classical complement pathway. As controls, sera from healthy blood donors were used. All sera were centrifuged, collected and frozen at −70°C within four hours of sampling. The local ethical committees at the Karolinska Hospital in Stockholm and the University Hospital in Uppsala approved the investigation.
PEG precipitation of immune complexes
Sera were precipitated with 3% (wt/vol) ice cold polyethylene glycol (PEG) 6000, centrifuged, and washed three times in sterile phosphate buffered saline (PBS) and subsequently diluted to the initial serum volume in sterile PBS. The PEG precipitates were used immediately for FcγR blocking experiments. (Erroneous results were produced when PEG precipitates were frozen and thawed before cell culture experiments.)
Preparation of heat aggregated immunoglobulins (HAGG)
Artificial ICs were prepared by heating human IgG (Gammagard, Baxter, Belgium), 50 mg/ml, to 63°C for 30 minutes immediately before each experiment, and diluted in PBS to the desired concentration. As a control, the same IgG preparation without heat treatment was used.
Preparation of mononuclear cells
Heparinised blood was obtained from healthy blood donors or laboratory personnel after informed consent. Blood samples were diluted in equal amounts of PBS at room temperature, then separated on Ficoll-Paque (Amersham Pharmacia, Uppsala, Sweden). After two washes in PBS, cells were suspended in RPMI-1640 (Flow Laboratories, Irvine, Scotland, UK) supplemented with glutamine, HEPES buffer, 10% native fetal calf serum (FCS; Flow Laboratories), penicillin, and streptomycin in adequate amounts. In the FcγR blocking experiments which were performed at a later time, FCS was exchanged for normal human serum supplemented with 1% Ultrocer G (Flow), as this was shown to better sustain IC-induced cytokine production. Cell suspensions were diluted with complete medium to 1×106 peripheral blood mononuclear cells (PBMC)/ml for supernatant cultures, to 5×105 PBMC/ml for IL10 enzyme linked immunospot (ELISPOT) investigations, and to 2×104 PBMC/ml for IL6 ELISPOT investigations.
All cytokine-producing responder cells used in this investigation were obtained from healthy controls. The magnitude of cytokine response to ICs differs substantially between PBMC preparations from different donors. A single predefined cell donor was therefore used for the experiments with SLE sera and SLE PEG precipitates, whereas 10 randomly selected blood donors were used in the cross sectional flow cytometry experiment.
Cytokine enzyme linked immunosorbent assays (ELISAs)
Supernatants were harvested after 20 hours’ incubation, and IL6 and IL10 concentrations were determined by ELISA following a recently described protocol.20 In some experiments the alkaline phosphatase system was exchanged for a horseradish peroxidase system employing 3,3‘,5,5‘-tetramethylbenzidine as substrate (DAKO A/S Glostrup, Denmark) for the IL10 ELISA. Monoclonal antibodies 13A5 (IL6) and 9D7 (IL10) were used as primary antibodies at a concentration of 1 μg/ml, and 39C3 (IL6) and 12G8 (IL10) were used as secondary antibodies at concentrations of 2 and 1 μg/ml, respectively. All monoclonal antibodies were obtained from Pharmingen (San Diego, CA, USA).
Cytokine ELISPOT method
The method has been described recently21, and also used in previous SLE studies in our laboratory.22,23 Monoclonal antibodies 13A5 recognising IL6 and 9D7 recognising IL10 (Pharmingen) were used as primary antibodies. One hundred microlitres of cell suspension was added to duplicate wells, followed by serum (10%), PEG precipitates (10%) or HAGG/control monomeric IgG. Plates were placed in a 37°C cell incubator with 5% CO2 for 20 hours. This time had earlier been shown to be optimal for the detection of IL6 and IL10 producing cells. After washing the plates, secondary biotinylated antibodies (39C3 for IL6 and 12G8 for IL10; Pharmingen) at 1 μg/ml were added and the plates incubated at 4°C overnight. As substrate for avidin alkaline phosphatase (DAKO), 50 μl of a BCIP phosphatase substrate solution (product code 710–3, Sigma, St Louis, MO, USA) was used, with a development time of 5 hours. After final washes in deionised water, the spot forming cells (SFC) were counted with an inverted microscope.
FcγR blocking experiments
For studies of the interaction between ICs and cell surface receptors, monoclonal antibodies IV.3 against FcγRII (Fab fragment) and 3G8 against FcγRIII (F(ab‘)2 fragment were obtained from Medarex (Nutley, NY). PBMC were incubated with FcγR blocking antibodies (1.5 μg/ml; concentrations defined in initial titration experiments) for 30 minutes at 4°C in a cell incubator before addition of HAGG or PEG precipitates. Cell suspensions were then incubated for 20 hours before the supernatants were harvested for ELISA evaluation of cytokine levels.
Flow cytometry
To determine correlation between monocyte FcγR expression and cytokine production, PBMC from 10 healthy blood donors were double stained with fluorescein isothiocyanate or phycoerythrin conjugated monoclonal antibodies against CD14 (clone TYK) and one of FcγRI/CD64 (clone 10.1), FcγRII/CD32 (clone KB61), or FcγRIII/CD16 (clone DJ130c). All antibodies were obtained from DAKO. Measurements were made on a FACSCalibur flow cytometer using the Cellquest software (Becton Dickinson, Mountain View, CA). Parallel cell cultures were stimulated with either HAGG or control IgG (100 μg/ml), and supernatant levels of IL10 and IL6 were measured as described above. Background cytokine production differed between the cell donors, and net IC-induced cytokine production was therefore calculated as the difference between cytokine levels in HAGG stimulated cultures and control IgG stimulated cultures. (In this and in all other experiments performed, adding 100 μg/ml of monomeric IgG to PBMC cultures had no apparent effects on cytokine production.) Correlation between the mean fluorescence intensity for the three FcγRs on gated CD14+ monocytes and net IC-induced cytokine production was then calculated.
Statistics
Non-parametric statistics were used throughout the study. For comparisons between patients and controls, the Mann-Whitney U test was used, and for estimation of the correlation between mean fluorescence for FcγR staining and cytokine production, Spearman’s rank correlation test was used.
RESULTS
Artificial immune complexes induce IL6 and IL10 production in PBMC cultures
Supernatant analyses cannot be used to investigate cytokine-inducing properties of intact serum samples. For that purpose, sensitive techniques measuring single cell cytokine production are more suitable because they are not influenced by carry over from the serum samples. First of all, we therefore wanted to investigate whether the ELISPOT technique could be used to detect IC-induced production of IL6 and IL10.
When HAGG was added to PBMC cultures overnight, a dose dependent increase in the number of IL10 producing cells was observed, whereas addition of control monomeric IgG had no effect (fig 1A). In contrast, the number of IL6 producing cells did not increase beyond the level in the control wells with monomeric IgG (fig 1B). However, density of the single IL6 spots increased substantially for HAGG samples at the highest concentrations, whereas no such effect was seen for monomeric IgG (data not shown). In supernatant experiments, both IL10 and IL6 showed a dose dependent increased in HAGG cultures as compared with monomeric IgG (figs 1C and D).
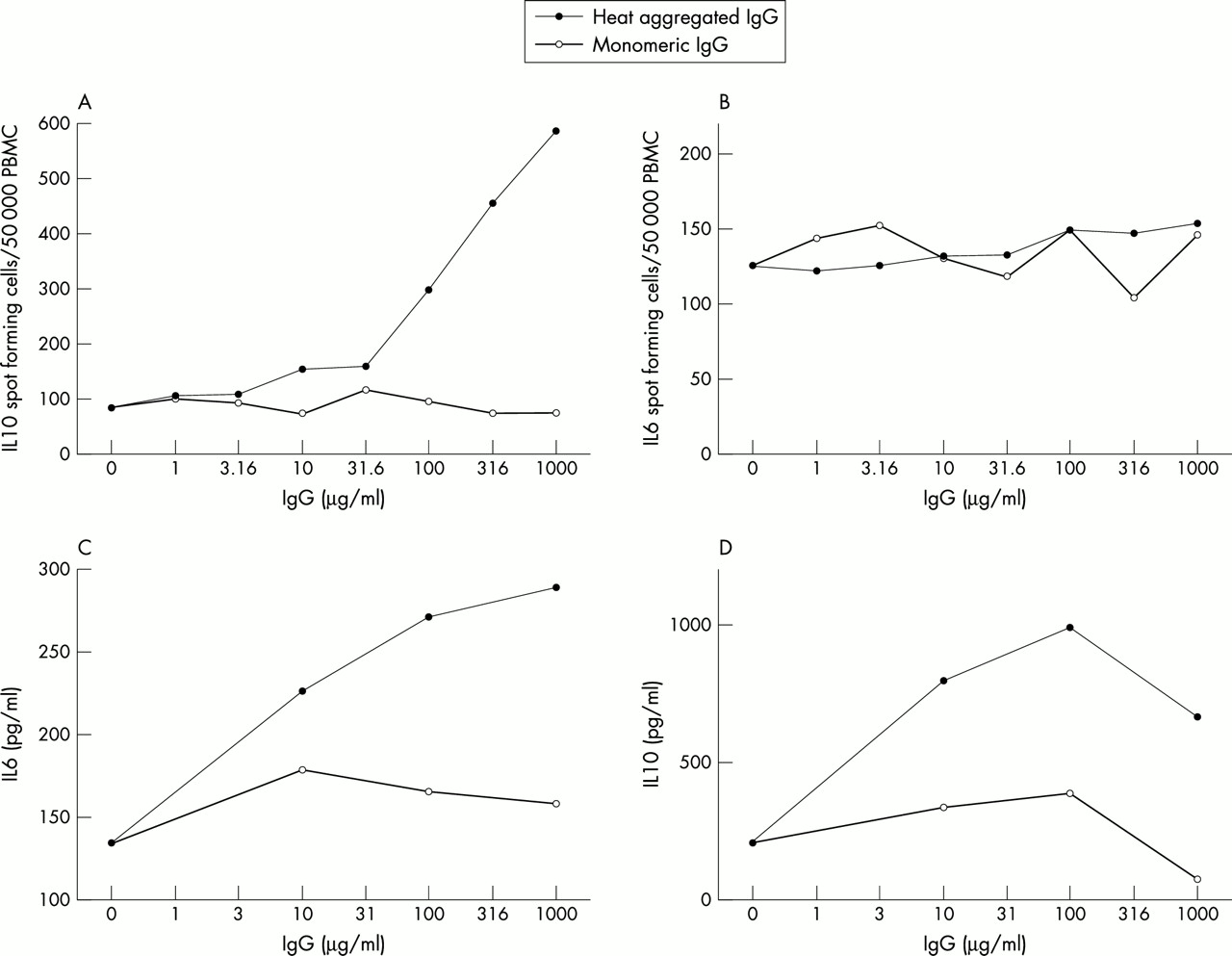
Heat aggregated IgG induces production of IL10 and IL6 in cultures of human PBMC by increasing the number of cells producing IL10, but not IL6. Human monomeric IgG was heat aggregated and diluted to various concentrations in PBS, and then added to PBMC cultures (5×105, 2×104, and 1×106 PBMC/ml for IL10 ELISPOT, IL6 ELISPOT, and supernatants, respectively) in complete medium and incubated overnight. As a control, non-aggregated IgG was used. (A) and (B) show the number of IL10 and IL6 spot forming cells, respectively. In parallel cultures, supernatant levels of (C) IL10 and (D) IL6 were determined by ELISA. These results are representative of five experiments for ELISPOT and supernatants respectively, using PBMC from different healthy donors.
Given these results, we decided to investigate the effects of SLE sera on single cell cytokine production for IL10 only.
By neutralising IL10, we often noted a suppressive effect of IC-induced IL10 on concomitant IL6 production (sometimes as high as 100% compared with culture supernatants with control antibodies; data not shown).
SLE sera have IL10 inducing properties
When serum samples from patients with SLE and from healthy controls were added to healthy control PBMC cultures overnight, a highly significant increase in the number of IL10 producing cells was noted in the cultures with SLE sera as compared with control sera (median 11.5 v 1.25 SFC/50 000 PBMC, p<0.0001; fig 2). The analysis was repeated using the same sera and responder cells, with the second run showing an equally significant difference between patients with SLE and controls (data not shown).

SLE sera stimulate an increase in the number of IL10 SFC. Twenty serum samples from patients with SLE and 18 sera from healthy controls (10% of final volume) were added to 100 μl PBMC suspensions from a healthy donor (5×105 cells/ml) in duplicate. After 20 hours of incubation, the number of IL10 producing cells was enumerated by the ELISPOT technique. The results are representative of two experiment using the same sera and cell donor, with comparable results. Horizontal bars show the median values for each group.
PEG precipitates from SLE sera induce IL10 production
When the SLE sera used above were subjected to PEG precipitation and the precipitates then added to PBMC cultures from a healthy control, an increase in the supernatant levels for IL10 (p=0.016) and for IL6 (p=0.042) was found, as compared with the cultures with PEG precipitates from control sera (figs 3A and B). We also tried to perform parallel ELISPOT measurements, but the addition of PEG precipitates yielded technically unacceptable results on two occasions, and the technique was therefore not used here.

High molecular weight PEG precipitates from SLE sera induce IL10 and IL6 production in human PBMC cultures. Serum samples from 16 patients with SLE and 17 healthy controls were subjected to PEG precipitation, and the precipitates were diluted to the original serum volume in PBS. Ten per cent (final concentration) of each precipitate was added to a 250 μl suspension of PBMC (1×106 cells/ml) from the healthy subject investigated in fig 2. After incubation for 20 hours, supernatant levels of (A) IL10 and (B) IL6 were assessed by ELISA. Horizontal bars show the median values for each group. Owing to a shortage of serum, not all of the samples studied in fig 2 were subject to PEG precipitation.
IL10 production induced by artificial ICs or by SLE PEG precipitates can be blocked by FcγRII antibodies
Addition of anti-FcγRII inhibited the increase in IL10 production induced by HAGG or by PEG precipitates from SLE sera. No blocking effect was seen when the same antibodies were added to a cell culture with PEG precipitate from normal serum, to cells with monomeric IgG or to unstimulated cells where no increase in IL10 production had been induced (fig 4).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Effects of blockade of FcγRII and FcγRIII on IC-induced cytokine production. Cell cultures were incubated with 1.5 μg/ml of blocking Fab (FcγRII) or F(ab‘)2 (FcγRIII) fragments for 30 minutes before addition of stimulus. PBMC (106/ml) from the healthy donor used in figs 2 and 3 were stimulated with PEG precipitates from three active SLE sera or with HAGG (100 μg/ml), or with a PEG precipitate from a healthy control (NHS), with monomeric IgG (mIgG; 100 μg/ml), or with unstimulated cells. Supernatants were collected after 20 hours’ incubation and levels of IL10 analysed by ELISA.
The effect of anti-FcγRIII antibodies varied between individual experiments with different PBMC donors, whereas the blocking effect of FcγRII antibodies was more consistent.
The effect of FcγR blockade on IC-induced IL6 production was more variable, and did not show the uniform results obtained for IL10. A blocking effect of FcγRII antibodies was, however, noted in most experiments (data not shown).
In initial experiments whole antibodies to FcγRI, II, and III were used, but these antibodies as well as isotype matched control antibodies both showed intrinsic cytokine stimulatory effects in our systems and were therefore abandoned. Attempts to block FcγRI with purified Fc fragments also regularly resulted in stimulation of cytokine production.
Monocyte FcγRII expression is correlated with IC-induced cytokine production
This experiment was performed to further substantiate the importance of FcγRII in IC-induced cytokine production. When PBMC from 10 healthy subjects were stained for expression of FcγRI/CD64, FcγRII/CD32, and FcγRIII/CD16 and parallel cultures were stimulated with HAGG or monomeric IgG, a statistical correlation was found between the monocyte expression of FcγRII/CD32 and IC-induced production of IL10 and IL6, whereas no such correlation was noted for either FcγRI/CD64 or FcγRIII/CD16. For IL10 this correlation was observed both with and without normalisation of the cytokine production for the monocyte percentage in PBMC cultures, whereas the correlation with IL6 was apparent only after correction for the percentage of monocytes in the cultures (table 1).
Correlation between monocyte expression of Fcγ receptors and cytokine production. PBMC from 10 healthy donors were stained for monocyte expression of FcγRI/CD64, FcγRII/CD32, and FcγRIII/CD16 and parallel cultures stimulated with HAGG or monomeric IgG (100 μg/ml). Figures represent correlation between mean FcγR expression and net IC-induced production of IL10 and IL6 (difference in cytokine levels in HAGG stimulated cultures and cultures stimulated with monomeric IgG)
DISCUSSION
In this study we have investigated the possibility that circulating ICs may stimulate the production of the B cell activating cytokines IL6 and IL10 in SLE. We have shown a highly significant increase in IL10 production in cell cultures incubated with SLE sera, as compared with cell cultures with control sera. The effect was at least partially localised to the PEG precipitate fraction, where high molecular weight ICs are to be found. IL10 production induced by either PEG precipitates from highly active SLE sera or by HAGG could be partially blocked by a monoclonal anti-FcγRII antibody, and the importance of FcγRII for IC-induced production of IL6 and IL10 was further substantiated in an experiment where monocyte surface expression of FcγRII correlated with IC-induced production of IL10 and IL6.
Earlier reports have shown that ELISPOT is 10–200 times more sensitive than supernatant measurement for the detection of produced cytokine.24 The ELISPOT technique may therefore have advantages in the detection of low grade cytokine production. Primary experiments using HAGG showed our ELISPOT method to be very sensitive in detecting the stimulatory effects of ICs on IL10 production. It did not, however, detect increased numbers of IL6 producing cells, although the single IL6 spots in the HAGG treated wells in repeated experiments showed markedly intensified staining for high concentrations of ICs as compared with the parallel control wells with monomeric IgG. In parallel cultures, supernatant levels of IL10 and IL6 were, however, both increased after HAGG stimulation. A possible interpretation is that ICs increase the number of IL10 producing cells, whereas in the case of IL6, cytokine production is increased on the basis of single cells but without increasing the number of secreting cells.
The FCS batch selected for the ELISPOT experiments also yielded substantial spontaneous cytokine production in supernatants, a fact that might account for the generally high cytokine production shown in fig 3. The difference between serum stimulation in fig 2 and PEG precipitate stimulation in fig 3 might, however, also depend on serum factors other than the contribution made by PEG precipitable ICs to IL10 induction by SLE sera.
In this study the IC stimulating effects on IL10 production were found in all experiments involving either artificial or SLE ICs, whereas a parallel effect on IL6 production was noted in most, but not all, experiments. The dose dependent effect of HAGG on IL6 production was in some cases much less obvious than for IL10, and it was in such cases only visible at the highest HAGG concentration used (1000 μg/ml; data not shown). The difference in the findings for IL10 and IL6 may be a reflection of IL10/IL6 cross regulation. A prominent effect of IL10 is its inhibitory effect on the production of other cytokines such as IFNγ25 and IL6.25,26 By neutralising in vitro produced IL10 in IC stimulated PBMC cultures, we also obtained increased levels of IL6. It therefore seems reasonable to regard our data on IC-induced IL6 as a possible net sum of the stimulatory effects of ICs and suppression by concomitantly produced IL10. The suppressive effect of IL10 on IL6 production in SLE has, however, been questioned, with one report showing this effect to be absent in the case of B cells and monocytes from patients with SLE but not in cells from normal controls27, whereas another paper states that SLE monocytes have normal suppressive IL6 response to exogenous IL10.28 In our study, however, we focused on the serological component of SLE, whereas all responder cells were obtained from healthy donors.
The IC-induced production of IL10 may be a phenomenon secondary to the primary IC-induced induction of other cytokines. TNFα can induce IL10 production from PBMC.29 Type I interferons, on the other hand, have dual effects, by stimulating IL10 production from T cells and suppressing monocyte production of IL10.30 IFNα is also implicated in the pathogenesis of SLE,31,32 and the relative order of induction of IFNα and IL10 by ICs should be investigated in further studies.
Two experiments were performed which aimed at assessing the importance of the differing FcγRs on IC-induced cytokine production. The IL10-inducing effect of three SLE PEG precipitates as well as of HAGG could be partially blocked by an FcγRII antibody, whereas no such effect was evident in control cultures (unstimulated cells, and cell cultures stimulated with normal serum, PEG precipitate or monomeric IgG, respectively). When 10 PBMC cultures were analysed for monocyte expression of FcγRs I, II, and III by flow cytometry, and the results were compared with the cytokine-inducing effects of a standardised IC preparation, a significant association was found between the expression of FcγRII on CD14+ cells and the IC-induced production of IL10 and IL6, whereas no such correlations could be found for FcγRII and FcγRIII.
Given the reported association between the FcγRII polymorphism R131 and severe SLE (see, for example, van der Pol and van de Winkel33), including the production of IC associated anti-C1q autoantibodies18,34 and hypocomplementaemia,35 the association between FcγRII and IC-induced cytokine production found in these two experiments strengthens the argument for the functional importance of FcγRII in SLE. Further investigations of the possible effects of FcγR usage on IC-induced cytokine production are currently underway.
The inhibitory effect of FcγRII blockade on IC-induced IL10 production has been repeated in many experiments in our laboratory, while the effect of FcγRIII blockade has been variable, with PBMC from some cell donors being blocked by antibodies to FcγRII and FcγRIII to about the same extent, whereas PBMC cultures from other donors showed unaltered or enhanced cytokine production after incubation with anti-FcγRIII F(ab‘)2 fragments (fig 4). Therefore, our data have yielded no conclusive evidence about the functional importance of FcγRIII for IC-induced cytokine production.
A recent paper has reported a parallel finding that IgG-rich fractions of SLE sera induce production of IFNα, and that this production can be blocked by FcγRII antibodies.36 Whether there are parallel effects on the induction of IL10 and IFNα by ICs has not yet been established.
IC-induced production of IL10 and IL6 in human PBMC cultures has been shown to reside in the monocyte fraction.13 The increased production of IL10 and IL6 in SLE has earlier been shown to stem from monocytes and B cells.1 The importance of monocytes/macrophages in IC-induced production of IL6 and IL10 is substantiated by our experiments showing a correlation between IC-induced cytokine production and monocyte FcγRII expression (table 1).
Levels of ICs in SLE sera have long been shown to correlate with disease activity, 37,38 as do serum levels of IL10.7,8 Serum IL10 has also shown an inverse correlation with serum C3 levels,7 which in SLE are lowered by IC-induced complement activation through the classical pathway and negatively correlated with levels of circulating ICs.
These earlier reports, together with the data obtained in this study, argue for a role for in vivo formed ICs in the induction of IL10 and IL6 from monocytes in SLE. These type 2 cytokines both stimulate production of antibodies, in general, and disease associated anti-DNA autoantibodies. In SLE, the effect might be increased production of ICs containing antibody, with the creation of a vicious cycle with IC-induced cytokine production paralleling increased IC deposition in tissues and resulting in SLE associated pathology. These effects suggest a more active role than earlier anticipated for ICs in the perpetuation of a type 2 cytokine dominated disease process in SLE.
Acknowledgments
This investigation was supported by grants from the Swedish Society of Medicine, King Gustav V’s 80-years fund, the Karolinska Institute, the Swedish Medical Research Council, the Swedish League Against Rheumatism, the af Ugglas Foundation, the Golge Foundation, the Hierta Foundation, the Craffoord Foundation, the Groschinsky Foundation, the Grönberg Foundation, the Svartz Foundation, the Jeppsson Foundation, the Bergvall Foundation, the Dahlin Foundation, and the Sund Foundation.