Article Text

Abstract
Objectives To dissect the role of toll-like receptor (TLR) signalling and B cell survival/proliferating factors in the crosstalk between rheumatoid arthritis synovial fibroblasts (RASF) and B cells.
Methods RASF, rheumatoid arthritis dermal fibroblasts (RADF) and osteoarthritis synovial fibroblasts (OASF) were analysed for the expression of B cell survival/proliferating factors BAFF and APRIL in resting conditions and upon stimulation with TLR2/TLR3/TLR4 ligands. Unswitched IgD+ B cells were co-cultured with RASF/OASF/RADF in the presence/absence of TLR ligands and with/without BAFF/APRIL blocking antibodies. Activation-induced cytidine deaminase (AID) mRNA expression, Iγ-Cμ and Iα-Cμ circular transcripts (CTs; markers of ongoing class-switching to IgG and IgA) and IgM/A/G production were measured to assess functional activation of B cells.
Results TLR3 and to a lesser extent TLR4, but not TLR2 stimulation, induced up to ∼1000-fold BAFF mRNA and increased soluble BAFF release. APRIL was less significantly regulated by TLR3. Resting and TLR3-stimulated RASF released higher levels of BAFF/APRIL compared with RADF. TLR3 stimulation of RASF but not RADF in co-culture with B cells strongly enhanced AID expression, Iγ-Cμ and Iα-Cμ CTs and class-switching to IgG/IgA. Blockade of BAFF/APRIL signalling completely inhibited TLR3-induced, RASF-dependent expression of AID, CTs and the secretion of IgG/IgA.
Conclusions RASF produce high levels of BAFF and APRIL constitutively and in response to TLR3 stimulation. These factors are critical in directly modulating AID expression, class-switch recombination and IgG/IgA production in IgD+ B cells. Overall, this work highlights a novel and fundamental role for the TLR3/B cell survival factor axis in sustaining B cell activation in the rheumatoid arthritis synovium.
Statistics from Altmetric.com
Introduction
Rheumatoid arthritis (RA) is a chronic autoimmune/inflammatory disease characterised by immune cell infiltration in the synovial tissue leading to cartilage destruction and bone erosion.1 One of the hallmarks of RA is a dysregulated activation of autoreactive B cells, leading to the production of autoantibodies such as rheumatoid factor and anti-citrullinated protein antibodies (ACPA), which are important diagnostic and prognostic markers and are implicated in disease pathogenesis.2 The critical relevance of B cells in RA pathogenesis is demonstrated by the clinical effectiveness of B cell depletion.3 4 However, the cellular and molecular mechanisms regulating autoreactive B cell activation in RA are poorly understood. We and others have shown that RA synovial tissues with ectopic lymphoid structures (ELSs), characterised by perivascular aggregates of T and B cells with the differentiation of follicular dendritic cell (FDC) networks,5 6 represent functional niches of autoreactive B cell activation and differentiation. In particular, ELSs promote the expression of activation-induced cytidine deaminase (AID), the enzyme required for somatic hypermutation and class-switch recombination (CSR) of the Ig genes, triggering synovial B cells to undergo in situ differentiation towards autoreactive plasma cells.7 8 ELS+ RA synovia engrafted into severe combined immunodeficient (SCID) mice sustain AID expression, B cell survival and proliferation, ongoing CSR, and production of ACPA in the absence of re-circulating immune cells,8 suggesting that resident synovial cells are critically involved in regulating humoral autoimmunity in RA. B cell functionality in the SCID/RA model is critically dependent on the B cell survival and proliferating factors BAFF and APRIL,8 9 although the source of these factors in the synovial microenvironment is unclear.
Rheumatoid arthritis synovial fibroblasts (RASF) share properties of FDCs in rescuing B cells from apoptosis and promoting their proliferation and differentiation into antibody-producing cells.10,–,14 However, the mechanisms regulating the crosstalk between RASF and B cells are unclear. Recent evidence demonstrated that B cells can be directly activated to undergo Ig class-switching in the presence of BAFF and APRIL released at mucosal sites by epithelial cells sensing pathogens via toll-like receptors (TLRs), particularly TLR3.15 16 RASF can express high levels of BAFF and APRIL,17 18 although the effective capacity of these factors in supporting AID expression and ongoing CSR is unclear. In addition, although accumulating evidence suggests that the activation state of RASF is critically dependent on the expression of TLRs,19,–,21 the relationship between TLRs and BAFF/APRIL expression in RASF is controversial.19,–,21
In the present work, we investigated the effect of TLR2/3/4 ligands in modulating BAFF/APRIL expression in RASF and their role in promoting AID expression and CSR in unswitched IgD+ B cells.
Materials and methods
Synovial tissue collection
Synovial tissues were obtained from patients undergoing total knee/hip replacement. A total of six RA and eight osteoarthritis (OA) samples were available at Queen Mary University of London and used for isolation of synovial fibroblasts (SF). Samples were collected after informed consent (LREC07/Q0605/29 granted by the East London & The City Research Ethics Committee 3). Matched synovial and skin fibroblasts from five RA patients undergoing total knee/hip replacement were obtained from the University of Birmingham (LREC5735), as previously reported.22 All RA patients (10 women/1 man, mean age 64.1 years, 82% rheumatoid factor positive) fulfilled the American College of Rheumatology criteria for the classification of RA23 and had long-standing disease requiring joint replacement. All patients had received disease-modifying antirheumatic drugs (in monotherapy or combination) and two patients received anti-TNF therapy. Patients had discontinued treatment before surgery for an appropriate washout period. Patients with OA had advanced disease and were all diagnosed with primary OA.
Generation of fibroblast-like synoviocytes from RA and OA patients
RASF and osteoarthritis synovial fibroblasts (OASF) were obtained as previously described.24 After discarding fat and dense fibrous tissues, synovium was minced, mashed under a 70 μm nylon mesh cell strainer (Becton Dickinson, Oxford, UK) and digested overnight at 37°C with 1.5 mg/ml Dispase II in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% foetal calf serum (FCS), 50 IU/ml penicillin-streptomycin and 10 mM HEPES buffer (Gibco/Invitrogen, Paisley, UK). The resulting suspension was passed through a cell strainer, centrifuged and the cell pellet was re-suspended in culture medium and incubated in T75 tissue-culture flasks. When 90% confluent, SFs were passaged 1/3 using 0.25% trypsin/EDTA (Sigma, Poole, UK). Synoviocytes were used between passages 4 and 8 when the culture is devoid of contaminating lymphocyte and macrophages.25
Purification of unswitched IgD+ human B cells
Human B cells were immunomagnetically selected from tonsils obtained following routine tonsillectomy. Tonsils were washed with chilled (4°C) MACS buffer (phosphate buffered saline (PBS), 0.5% FCS and 2 mM EDTA), minced and gently mashed through a cell strainer. Cells were washed, centrifuged and incubated with 0.5 μg/106 cells of an anti-IgD biotinylated antibody (Southern Biotechnologies, Birmingham, Alabama, USA) for 30 min on ice. After washing, cells were incubated with anti-biotin MicroBeads (Miltenyi Biotec, Bisley, UK) for 15 min at 4°C, washed and passed through magnetic separation columns (MS; Miltenyi Biotec). The effluent was collected as flow through fractions and the bead-bound cells (after-sorted) were collected as enriched, positively selected IgD+ B cells. All sorting procedures were performed under sterile conditions. The purity of IgD+ unswitched B cell isolation was checked by fluorescence-activated cell sorting (FACS; online supplementary figure 1).
TLR stimulation of RASF, OASF and rheumatoid arthritis dermal fibroblasts (RADF)
Fibroblasts were seeded into 24-well plates at 4×104 cells/well and stimulated at 90% confluence with optimal concentrations of TLR2–4 ligands19 (300 ng/ml of bacterial lipopeptide, 10 µg/ml of polyinosinic:polycytidylic acid (PIC) (InvivoGen, Toulouse, France), 100 ng/ml of lipopolysaccharide (LPS) from Escherichia coli J5 (Sigma)) or left untreated. Supernatants were harvested after the allocated time period, centrifuged and stored at −20°C until tested. For RNA extraction, adherent fibroblasts were rinsed with PBS and lysed in RLT buffer (Qiagen, Crawley, UK).
Co-cultures of B cells with RASF, OASF and RADF
Fibroblasts were seeded into 24-well plates (4×104 cells/1 ml/well) and used for co-culture experiments at 90% confluence. In cell–cell contact experiments, IgD+ B cells (1×106 cells/300 µl/well) were added onto the fibroblast layer with or without 10 µg/ml PIC. In transwell experiments, microporous PET cell inserts (1 µm pore; Becton Dickinson) were employed. B cells were grown in the upper well and PIC treated or untreated fibroblasts were grown on the base of the well. Control co-cultures received no fibroblasts in order to investigate baseline expression in isolated IgD+ B cells. Co-cultures were kept for 24 h, 72 h or 8 days, depending on the experimental conditions after which cell-free supernatants and B cells were collected. For gene expression profiling, pelleted B cells were lysed in RLT buffer.
Blockade of BAFF/APRIL signalling on B cells with anti-BCMA and anti-BAFFR neutralising antibodies
BAFF/APRIL-dependent signalling in B cells was blocked using neutralising polyclonal goat anti-human BAFF receptor (BAFFR)and anti-human B-cell maturation antigen (BCMA) antibodies (R&D Systems, Abingdon, UK) at 10 µg/ml alone or in combination, 30 min before co-culture and throughout the experiment. After 72 h or 8 days of co-culture, supernatants and cells were harvested as described above.
Gene expression analysis by quantitative real-time PCR
Total RNA was isolated using the RNeasy Mini kit (Qiagen). RNA (500 ng) was reverse transcribed to cDNA using the Thermoscript RT-PCR System (Invitrogen). For gene expression profiling quantitative real-time-PCR was performed on cDNAs using specific TaqMan primers and probes8 (Applied Biosystems, Warrington, UK; online supplementary table 1). Taqman quantitative real-time PCR was run in triplicate with 10 ng/well at 40 cycles of amplifications in an ABI 7900HT Instrument. Human 18S and human Β-actin were used as endogenous controls. Relative quantification was measured using the Comparative CT (Threshold Cycle) Method.
Detection of circular transcripts by PCR
Detection of Iγ1/2-Cμ and Iα-Cμ circular transcripts (CTs) were performed as previously described with minor modifications8 using 0.5 U of Taq polymerase (MP Biomedicals, Illkirch, France)/20 µl reaction volume. Primers sequences and PCR conditions are summarised in online supplementary table 2. After final extension at 72°C for 5 min, PCR products were run on 1% agarose gel/0.5 μg/ml ethidium bromide in tris-acetate-EDTA buffer and visualised under UV light.
ELISA
BAFF and APRIL protein concentrations in cell-free supernatants were measured using the Quantikine Human BAFF/BLyS/TNFSF13B (R&D Systems) and Human APRIL (eBioscience, Hatfield, UK) ELISA kits, respectively, following the manufacturers' instructions.
Total IgA, IgG and IgM were detected using the Human IgA/IgG/IgM ELISA quantitation set (Bethyl Laboratories, Cambridge, UK) according to the manufacturer's instruction.
Detection of BAFF and APRIL expression in fibroblasts using immunocytochemistry
Fibroblasts were seeded at 5×104 cells/200 µl onto cover slides and grown to 90% confluence. Cells were then stimulated with 10 µg/ml PIC or left untreated. After 24 h, cells were washed in PBS and fixed using ice-cold 1:1 acetone:methanol. After washing in Tris-buffered Saline (TBS) and blocking with avidin-biotin blocking kit (Vector Laboratory, Peterborough, UK) and Dako, Ely, UK serum-free protein block, rat anti-human BAFF (Buffy-2; Enzo Life Sciences, Exeter, UK), mouse anti-human APRIL (Aprily-2, Enzo Life Sciences) or isotype controls at 10 μg/ml were applied for 1 h at room temperature. After washing, biotinylated donkey anti-rat (Serotec, Kidlington, UK) or rabbit anti-mouse (Dako) antibodies were added for 1 h at room temperature. After washing, slides were incubated for 30 min with Streptavidin-ALEXA555 or Streptavidin-FITC (1:300 dilution) and counterstained with DAPI. Slides were examined under Olympus BX60 microscope.
Statistical analysis
Differences in quantitative variables were analysed by the Mann–Whitney U test when comparing two groups and by the Kruskal–Wallis with Dunn's post-test when comparing multiple groups. Wilcoxon's signed-rank test was used to analyse repeated measurements at different time-points. All analyses were performed using GraphPad Prism version 5.01 (GraphPad Software, USA). A p value <0.05 was considered statistically significant.
Results
TLR3-stimulated induction of BAFF and APRIL mRNA in RASF, OASF and RADF
We first examined whether TLR stimulation of RASF and OASF (to evaluate disease specificity) and paired RADF (to investigate site specificity) induced BAFF and APRIL mRNA. At baseline, RASF and OASF displayed increased expression of BAFF mRNA, between four- and eightfold higher than RADF (data not shown), as previously reported.17 BAFF mRNA expression in RASF and OASF was markedly induced by PIC (figure 1A, E), while there was a more modest induction with LPS and no upregulation with bacterial lipopeptide. PIC triggered a rapid induction of BAFF transcripts at 4 h, which peaked at 8 h and gradually decreased afterwards. A comparable increase in BAFF mRNA was induced by PIC in RADF (figure 1C). Interestingly, RADF produced significantly higher levels of BAFF mRNA in response to TLR4 stimulation than RASF or OASF, and similar levels to that obtained with PIC.
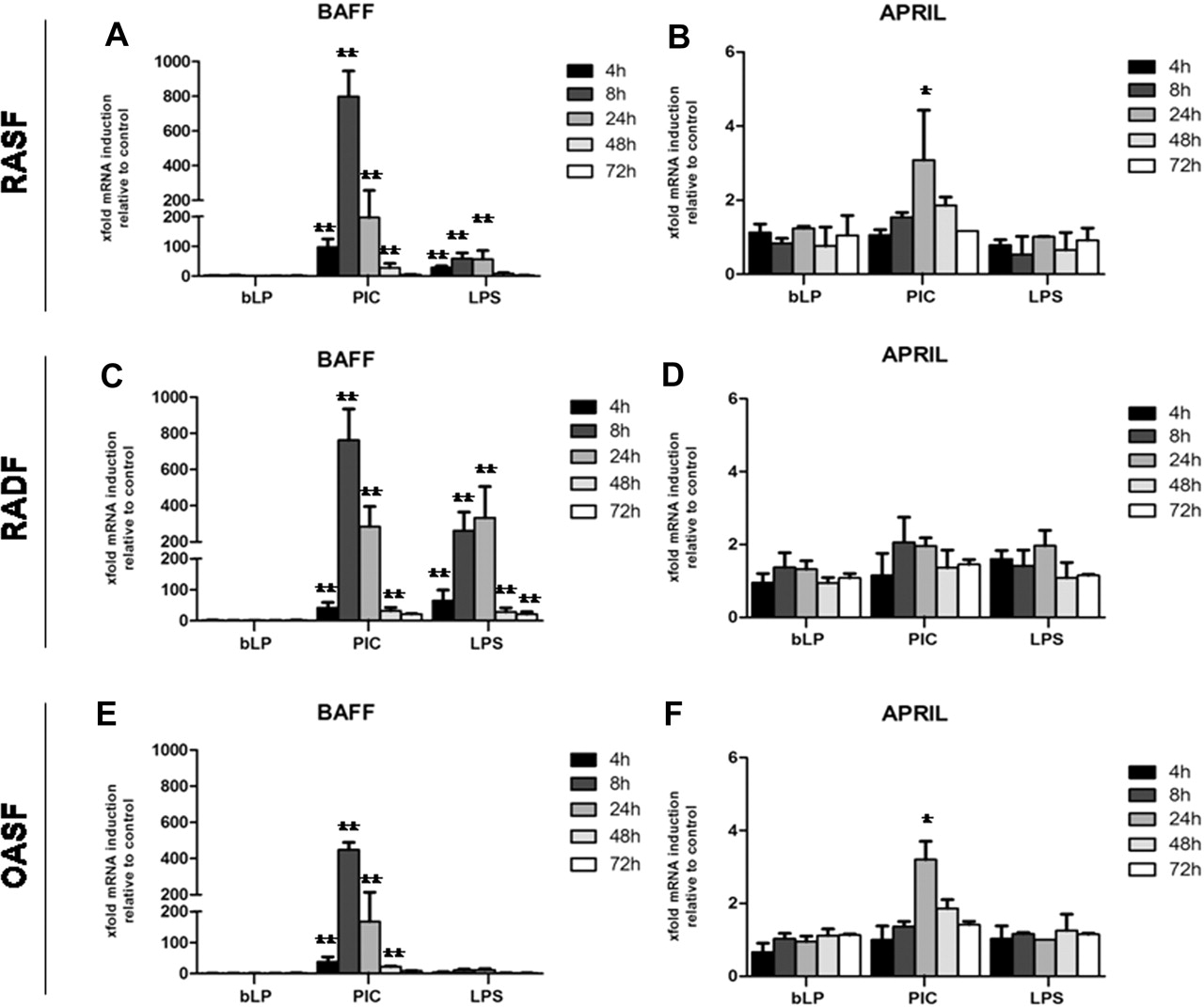
Induction of BAFF and APRIL mRNA in RASF, RADF and OASF in response to TLR ligands. Quantitative Taqman real-time PCR analysis of BAFF and APRIL mRNA expression in response to TLR ligands. RASF (A and B), RADF (C and D) and OASF (E and F) were stimulated with 10 µg/ml PIC (TLR3), 100 ng/ml lipopolysaccharide (TLR4), 300 ng/ml bLP (TLR2) or medium alone for 4, 8, 24, 48 and 72 h. Results are expressed as mean±SEM of the mRNA fold increase calculated at each time point by setting the baseline unstimulated expression as 1. Data were normalised using mammalian 18S as an endogenous control gene (18S). *p<0.05, **p<0.01 compared with unstimulated. OASF, synovial fibroblasts from osteoarthritis patients; PIC, polyinosinic:polycytidylic acid; RADF, rheumatoid arthritis dermal fibroblast; RASF, rheumatoid arthritis synovial fibroblast; TLR, toll-like receptor.
In comparison with BAFF, APRIL was significantly but more modestly modulated by TLR3 stimulation in RASF and OASF (∼threefold increase) (figure 1B, F) with no apparent modulation in RADF (figure 1D), and was characterised by a later peak of induction at 24 h. We finally investigated whether there was a differential expression of TLR3 mRNA in RASF compared with RADF and found no difference among these cells in basal TLR3 expression (online supplementary figure 3).
RASF but not RADF release high levels of BAFF in response to TLR3 stimulation
We next examined whether TLR3-induced BAFF mRNA was mirrored by increased protein expression. As shown in figure 2A, high amounts of soluble BAFF were released in the supernatant of RASF in a time-dependent manner and were significantly increased upon PIC stimulation (figure 2A). Similar levels were observed in the supernatant of OASF (data not shown). Despite strong upregulation of BAFF mRNA, RADF produced only modest amount of soluble BAFF in resting conditions and in response to PIC.

TLR3 stimulation induces intracellular and soluble BAFF in RASF but not RADF. (A) Column-bar graph showing the amount of soluble BAFF released in the cell culture supernatant from RASF and RADF conditioned for 24, 48 and 72 h in the presence or absence of 10 μg/ml PIC. RASF produce significantly higher amount of BAFF both in resting and stimulated conditions than RADF. Results are represented as mean±SEM of minimum three experiments. (B) Representative microphotographs of BAFF immunostaining on RASF and RADF before and after stimulation with PIC. RASF/RADF were grown on coverslips and processed for immunofluorescence as described in the Methods. Nuclei are counterstained in blue (DAPI) and BAFF in red (Alexa-555). Primary RASF/RADF were left untreated (medium only) or stimulated for 24 h with PIC (10 μg/ml). Appropriate isotype control was used to confirm specificity. Images were captured at 20× magnification and with the same exposure time for both conditions. PIC, polyinosinic:polycytidylic acid; RADF, rheumatoid arthritis dermal fibroblast; RASF, rheumatoid arthritis synovial fibroblast; TLR, toll-like receptor.
We next investigated the intracellular localisation of BAFF upon TLR3 stimulation by immunofluorescence. As shown in figure 2B, weak and granular perinuclear BAFF immunoreactivity was detected in untreated permeabilised RASF. Upon incubation with PIC, a strong increase in cytoplasmic BAFF (figure 2B) as well as membrane-bound BAFF (online supplementary figure 4) was observed. Similar data were observed in OASF (data not shown). In contrast, following exposure to PIC, BAFF expression remained barely detectable in RADF. Overall, these data indicate that increased production of BAFF, at basal level or upon TLR3 stimulation, is characteristic of SF but not DF.
RASF but not RADF constitutively produce large amounts of APRIL
We next investigated the ability of RASF and RADF to release APRIL in resting and stimulated conditions. Unlike RADF, RASF produced abundant basal levels of soluble APRIL, reaching ∼900 pg/ml on day 3 (figure 3A) which was only marginally increased upon TLR3 stimulation (figure 3A). However, APRIL was released at a higher rate within the first 24 h in PIC-stimulated RASF. Again, similar results were observed in OASF (data not shown). In contrast to RASF, APRIL levels in RADF were extremely low in basal conditions and remained low following TLR3 activation. Immunofluorescence staining for APRIL demonstrated that while resting RASF expressed an intense perinuclear and cytoplasmic immunoreactivity (figure 3B), likely corresponding to vesicle-sequestered or Golgi-associated APRIL,26 RADF were characterised by a significantly weaker staining. Overall, our findings indicated that RASF, but not RADF, retain the capacity to release constitutively high levels of APRIL, which are modestly increased upon TLR3 stimulation.

Intracellular and soluble APRIL expression in RASF vs RADF. (A) Column-bar graph showing the amount of soluble APRIL released in the cell culture supernatant from RASF and RADF conditioned for 24, 48 and 72 h in the presence or absence of 10 μg/ml PIC. Data are presented as mean±SEM of at least three experiments. (B) Representative microphotographs of APRIL immunostaining on RASF and RADF in resting conditions. RASF/RADF were grown on coverslips and processed for immunofluorescence as described in the Methods. Nuclei are counterstained in blue (DAPI) and APRIL is stained in green (Alexa-488). Appropriate isotype control was used to confirm specificity. Images were captured at 20× magnification and with the same exposure time for both conditions. PIC, polyinosinic:polycytidylic acid; RADF, rheumatoid arthritis dermal fibroblast; RASF, rheumatoid arthritis synovial fibroblast.
TLR3 stimulation of RASF, but not RADF, enhances class-switched immunoglobulin secretion from IgD+ human B lymphocytes
We next examined whether TLR3 stimulation enhanced the capacity of RASF to promote Ig production in B lymphocytes12 and whether this ability was dependent on direct cell–cell interactions and/or soluble factors.
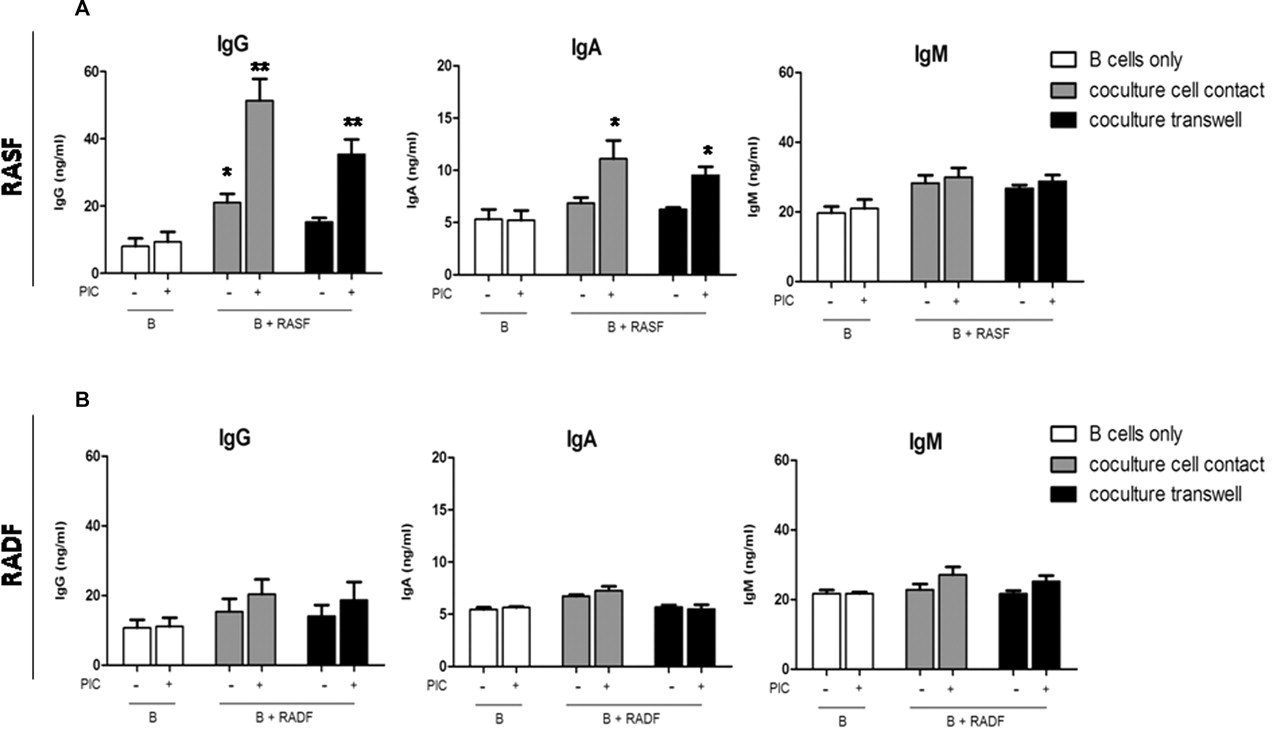
Tonsil IgD+ unswitched B cells at 97% purity (online supplementary figure 1) were co-cultured either in cell–cell contact or in a transwell system with RASF, OASF or RADF. As shown in figure 4, IgD+ B cells alone produced low amounts of IgG and IgA (figure 4A). Conversely, co-culture of B cells with RASF induced a two- to threefold increase in IgG production already at 72 h, which was significantly increased upon TLR3 stimulation (figure 4A). Similar, albeit more modest, induction of IgA was also evident. Paired RADF did not induce IgG or IgA production either in resting or stimulated conditions (figure 4B), while OASF displayed a similar behaviour to RASF (data not shown). Interestingly, CSR to IgG/IgA still occurred (albeit with slightly lower efficiency) when B cells were cultured in transwells (figure 4A), indicating that soluble factors released by RASF are sufficient to promote CSR. Treatment of RASF with PIC minimally modulated IgM production, suggesting that RASF induced functional B cell activation and not simply increased survival/proliferation, as indicated by the increased IgG:IgM ratio after 8 days compared with 24 and 72 h (figure 5D).
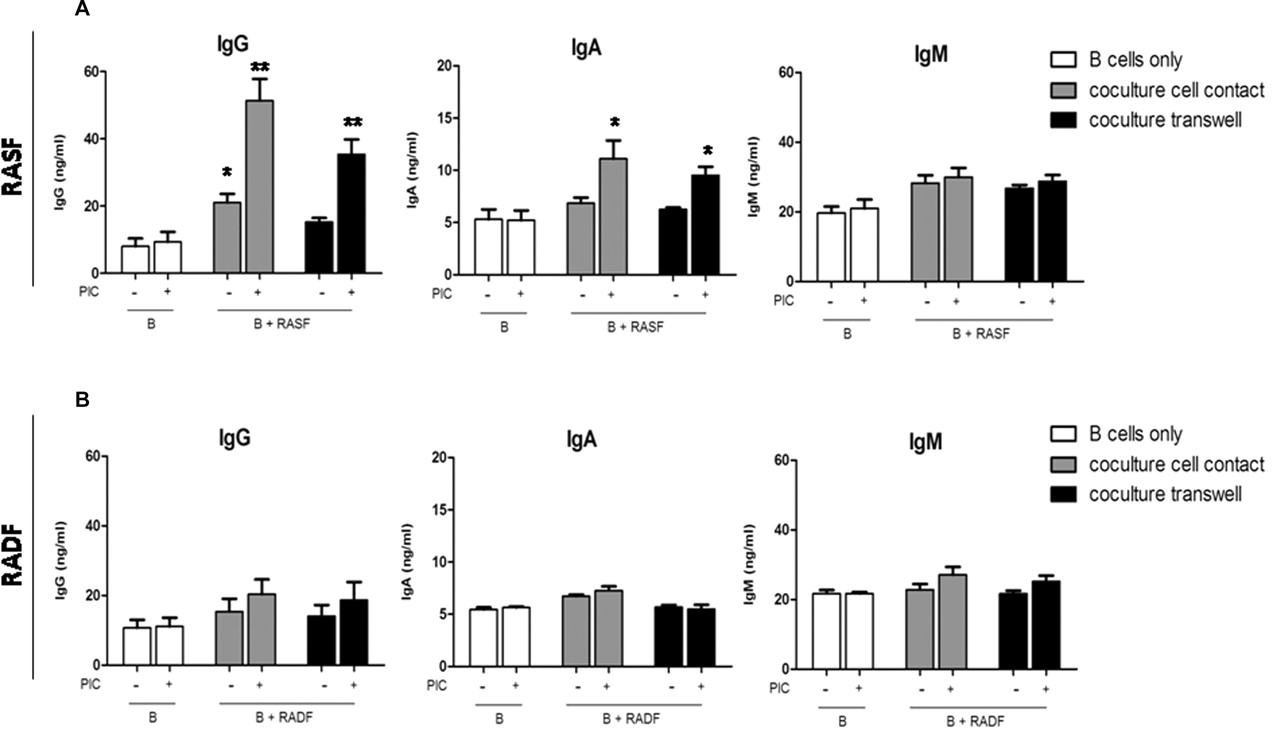
TLR3 stimulation of RASF but not RADF enhances their capacity to promote the production of class-switched antibodies in IgD+ B cells. Purified tonsillar IgD+ B cells were cultured with fibroblasts for 72 h. B cells were cultured alone (open bars) or in co-culture fibroblasts in direct physical contact (grey bars) or a transwell system (black bar). All cultures were incubated with (+) or without (−) 10 μg/ml PIC. IgG, IgA and IgM levels in supernatants were measured by ELISA. Results are presented as mean±SEM of at least three experiments. *p<0.05, **p<0.01 compared with unstimulated. PIC, polyinosinic:polycytidylic acid; RADF, rheumatoid arthritis dermal fibroblast; RASF, rheumatoid arthritis synovial fibroblast; TLR, toll-like receptor.
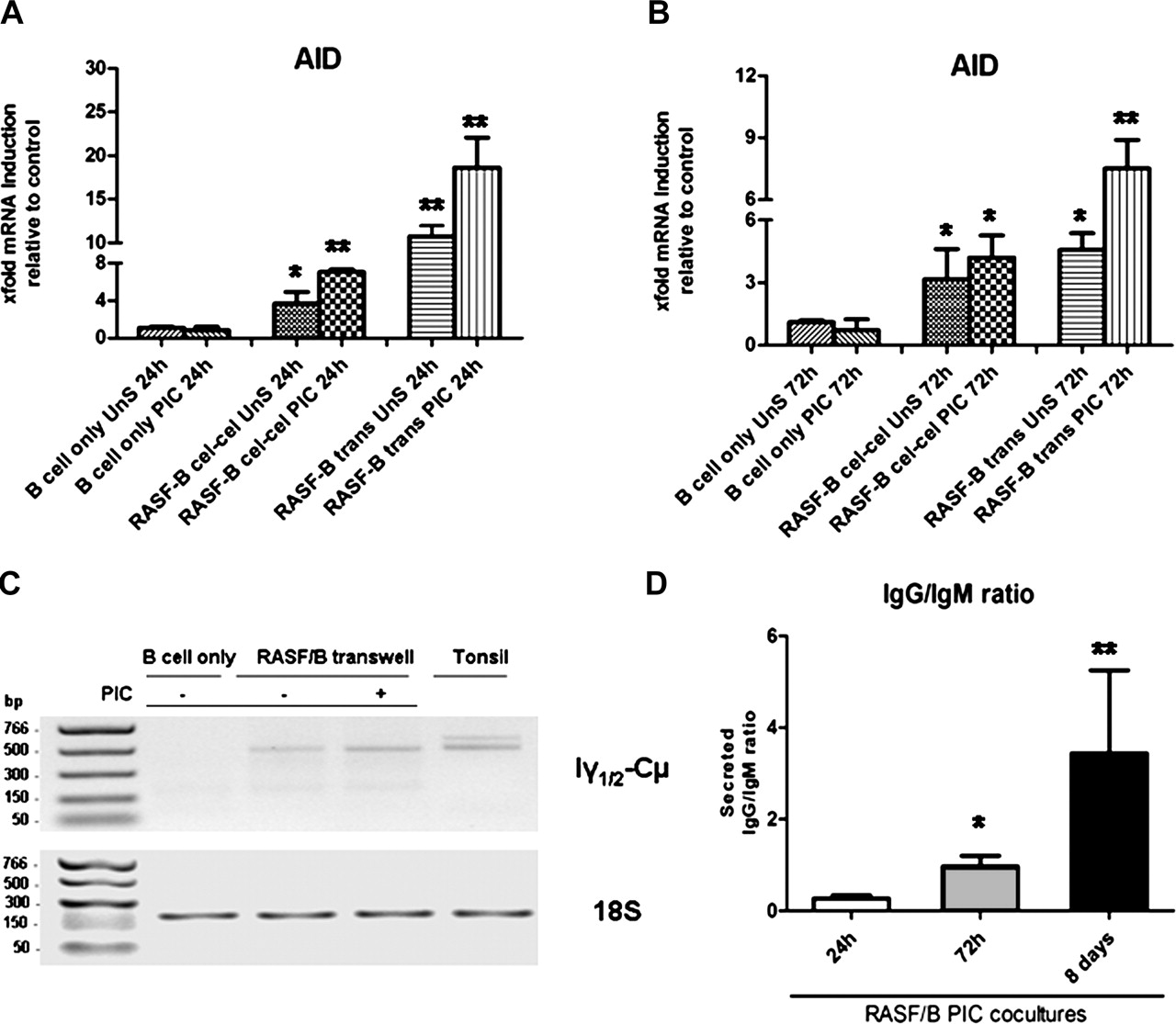
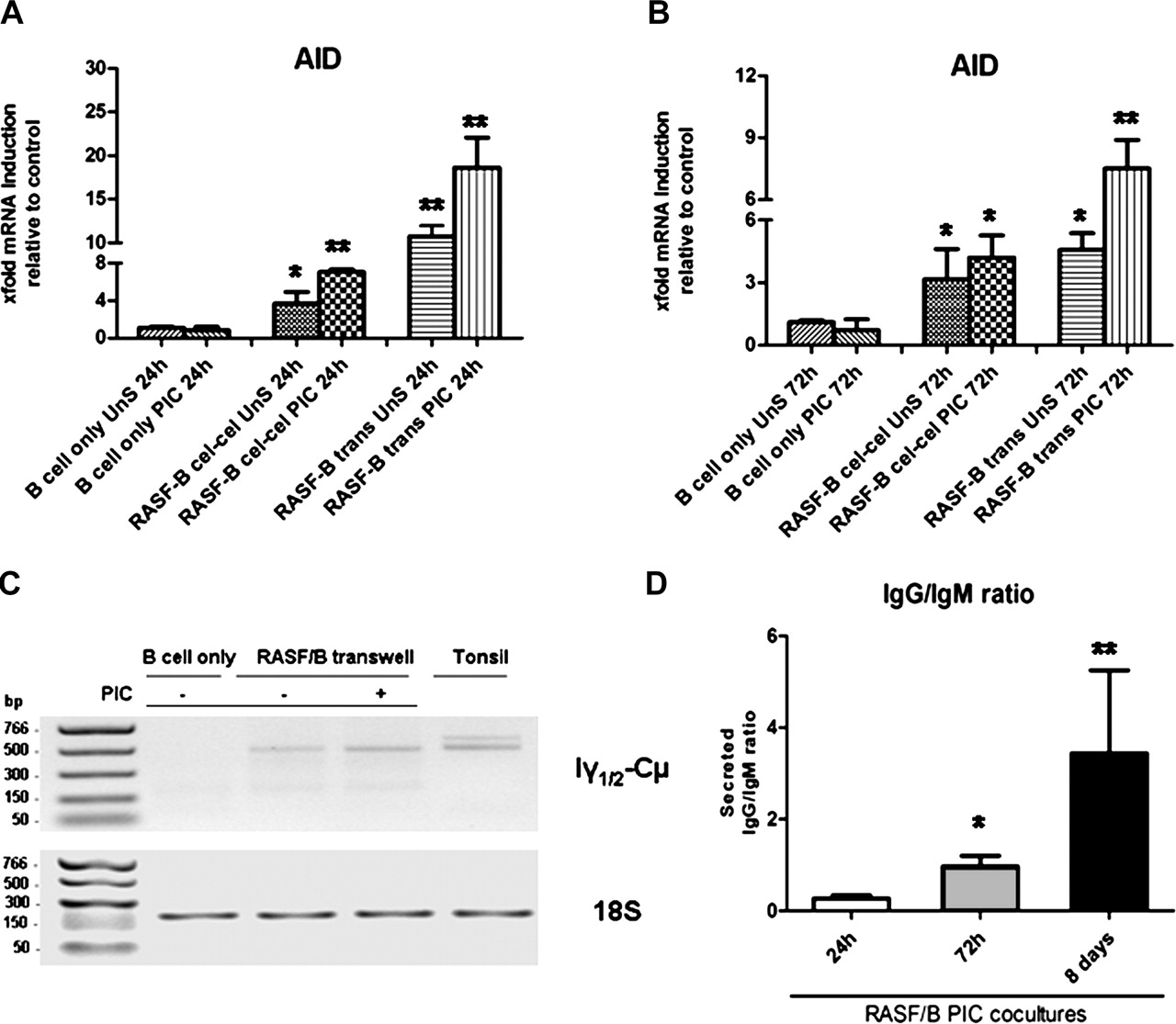
TLR3 stimulation of RASF enhances activation-induced cytidine deaminase (AID) expression and ongoing class-switching in co-cultured IgD+ tonsillar B cells. (A and B) Purified tonsillar IgD+ B cells were cultured with RASF for 24 h (A) and 72 h (B) with 10 μg/ml PIC or left untreated in the presence (transwell) or absence (cell–cell contact) of a cell insert. AID mRNA expression was determined by quantitative real-time PCR normalised to an endogenous control gene (18S) and the fold difference relative to baseline expression was calculated by setting AID expression in B cell alone as 1. *p<0.05, **p<0.01 compared with unstimulated. (C) Representative 1% agarose gel (inverted colour) of Iγ-Cµ circular transcripts (CTs; amplicon 502 bp, upper gel), specific by-products of ongoing class-switching from IgM to IgG. The lower gel represents the endogenous control, mammalian 18S (amplicon 187 bp). CTs were amplified from B cell cDNA using specific primers (Methods). B cells were cultured alone or in transwell with RASF for 72 h in the presence or absence of 10 μg/ml PIC. Tonsil was used as a positive control. (D) Progressive production of IgG over IgM in B cells co-cultured in transwell with TLR3-stimulated RASF after 24 h, 72 h and 8 days, demonstrating. Data are presented as mean IgG to IgM ratio ±SEM of at least three experiments per time point. *p<0.05, **p<0.01 compared with unstimulated. PIC, polyinosinic:polycytidylic acid; RASF, rheumatoid arthritis synovial fibroblast; TLR, toll-like receptor.
TLR3-stimulated RASF induce upregulation of AID and CTs in B cells
A formal demonstration of the capacity of RASF to induce class-switching require evidence of AID expression and the appearance of CTs. RASF strongly induced AID mRNA after 24 and 72 h of co-culture in cell–cell and transwell systems, a capacity further enhanced upon TLR3 stimulation (figure 5A, B). IgD+ B cells alone (figure 5A, B) or cultured with RADF (data not shown) displayed no or barely detectable levels of AID mRNA. The specificity of the AID and endogenous control PCR products were confirmed by agarose gel electrophoresis (online supplementary figure 2C). In order to confirm that AID was functional, we determined the presence of Iγ1/2-Cµ CTs, specific by-products of ongoing CSR from IgM to IgG1/2 in B cells, which disappear 48 h after CSR.27 While no Iγ1/2-Cµ CTs were detected in IgD+ B cells alone at 72 h, co-culture with unstimulated RASF was sufficient to induce CTs expression which was clearly enhanced upon TLR3 stimulation (figure 5C, top gel, lane 4). RASF-dependent induction of Iγ1/2-Cµ CTs and IgG production was similar to that observed using recombinant BAFF and IL4 (online supplementary figure 2).
Overall, these data provide conclusive evidence that RASF directly induce the expression of AID and the molecular machinery required for ongoing CSR in unswitched B cells and that TLR3 stimulation enhances this effect.
Dual blockade of BAFFR and BCMA abrogates TLR3-induced AID expression and class-switching
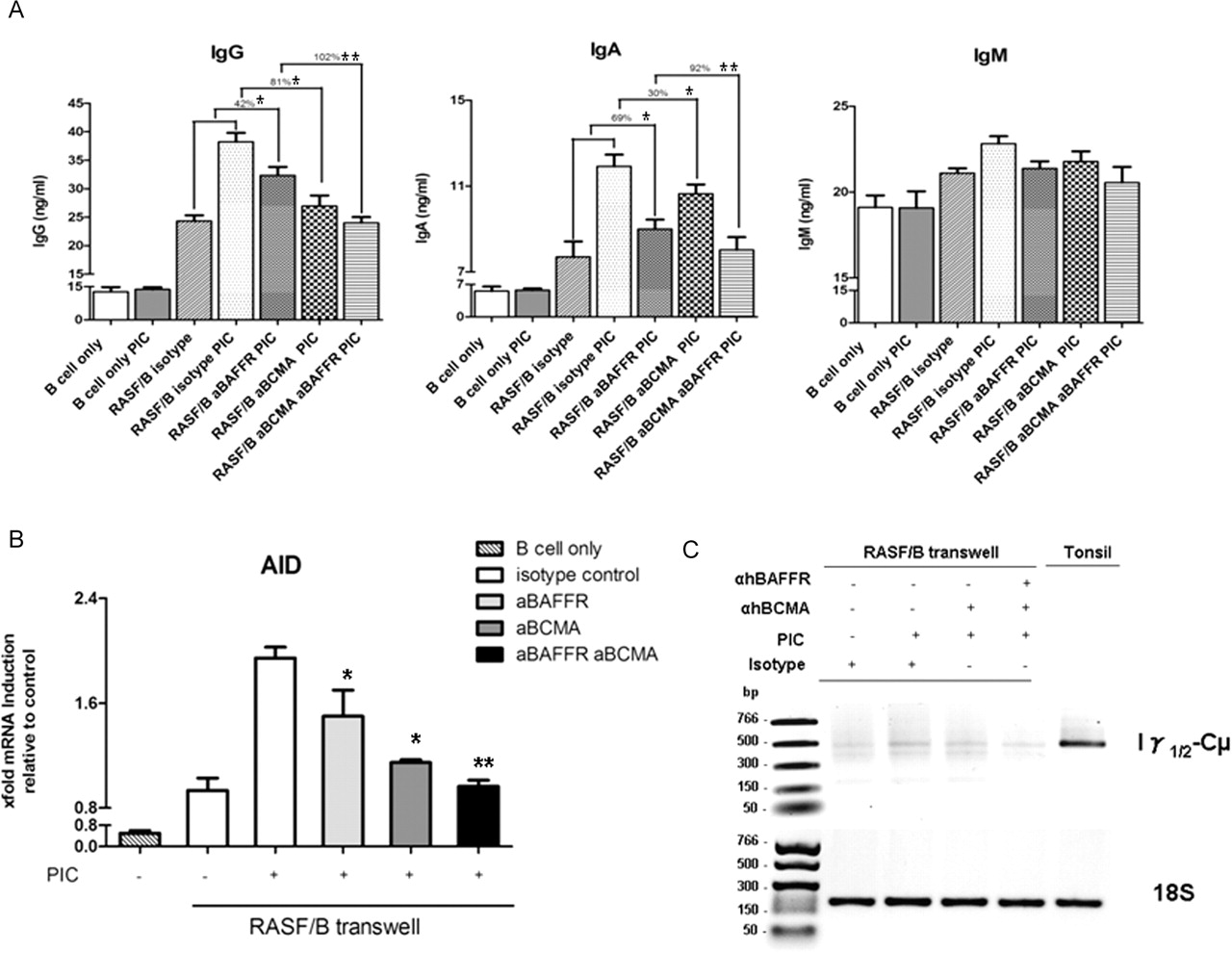
We finally investigated whether the activating capacity of RASF in B cells in response to TLR3 was dependent on BAFF and/or APRIL. By using neutralising antibodies against BAFFR (specific for BAFF) and BCMA (binding both BAFF and APRIL), we showed that in the transwell system individual receptor blocking displayed significant but partial inhibition of TLR3-dependent IgG and IgA production, while the combination of anti-BAFFR/anti-BCMA completely suppressed TLR3-induced IgG and IgA production (figure 6A). In addition, blockade of BAFFR/BCMA downregulated AID expression and the production of Iγ1/2-Cµ CTs (figure 6B, C, respectively). Experiments in cell–cell contact displayed similar results, although the inhibitory effect was slightly smaller (data not shown). Overall, these findings suggest that CSR induced in IgD+ B cells by RASF via TLR3 stimulation is mostly dependent on the production of BAFF and APRIL and can be blocked by receptor interference.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Blockade of BAFFR and/or BCMA reverses TLR3 induced RASF-dependent IgD+ B cell activation. (A) IgG, IgA and IgM production by tonsillar IgD+ B cells cultured in transwell with RASF for 72 h in the presence of 10 μg/ml of either anti-human BCMA or anti-human BAFF-R blocking antibodies or the two combined. Suppression of IgG and IgA production is expressed as the percentage of reduction of the effect induced by TLR3 stimulation. *p<0.05, **p<0.01 compared with PIC stimulated. (B) Modulation of activation-induced cytidine deaminase mRNA expression in B cells co-cultured in transwell with RASF for 72 h after blockade of BAFFR and/or BCMA signalling. Results are expressed as mean±SEM of the mRNA fold increase calculated by setting the baseline expression as 1. *p<0.05, **p<0.01 compared with PIC stimulated. (C) Representative 1% agarose gel (inverted colour) of Iγ-Cµ circular transcripts (CTs, amplicon 502 bp, upper gel) expression in B cells co-cultured in transwell with RASF for 72 h after blockade of BAFFR and/or BCMA signalling. Endogenous control, mammalian 18S (amplicon 187 bp) is shown in the lower gel. PIC, polyinosinic:polycytidylic acid; RASF, rheumatoid arthritis synovial fibroblast; TLR, toll-like receptor.
Discussion
In situ autoreactive B cell activation and autoantibody production within the RA synovium are typical features of RA patients in whom synovitis is characterised by the formation of ELSs. Within RA lymphoid aggregates, B lymphocytes undergo antigen-driven affinity maturation, clonal expansion, Ig isotype switching and plasma cell differentiation.7 28 29 We recently showed that these processes: (1) require the expression of AID, the enzyme responsible for the initiation of somatic hypermutation and CSR of the Ig genes, (2) are self-sustained for several weeks in the SCID/RA chimeric model in the absence of recirculating immune cells and (3) develop in the context of prolonged expression of the B cells survival and proliferating factors BAFF and APRIL.8 The functionality of synovial B cell follicles is dependent on the interaction with networks of stromal cells which express CD215 6 and closely resemble FDCs in secondary lymphoid organs.30 Accordingly, the diffuse form of synovitis, characterised by the absence of B cell aggregates, lacks AID expression and ongoing Ig class-switching.8 Despite strong evidence that ELSs support the production of autoantibodies within the RA synovium, we and others demonstrated that circulating autoantibodies in RA do not associate with the presence of synovial ELSs,8 31 32 suggesting that secondary lymphoid organs are also important sites for autoreactive B cell activation.
Although it is currently unclear whether synovial FDCs differentiate from resident stromal cells,33 fibroblast-like synoviocytes isolated from the RA synovium (RASF) are able to rescue B cells from apoptosis and promote their differentiation into antibody-producing cells.10,–,13 34 35 However, the critical factors regulating these interactions have not been fully investigated. Cell-dependent mechanisms involving adhesion molecules (ie, VCAM-1) and chemokines such as CXCL12 only partially account for the observed B cell activation,10 suggesting a critical contribution of additional factors. In addition, because previous work did not differentiate between naïve, switched and unswitched memory B cells in co-culture experiments, the actual capacity of RASF to induce the molecular machinery required for Ig class-switching in naïve B cells was questionable.
In this work, we provide novel observations that contribute to our understanding of the mechanisms regulating the crosstalk between RASF and B cells within the RA synovium. First, we demonstrated that RASF constitutively produce large amount of BAFF and APRIL, expanding recent evidence of overexpression of these factors at mRNA level in RASF.17 18 36 Furthermore, we showed that TLR3 stimulation induced strong and rapid upregulation of BAFF at both mRNA and protein level while APRIL was more modestly modulated and displayed a delayed kinetic of expression, peaking 24–48 h poststimulation. Interestingly, soluble BAFF levels in resting and TLR3-activated RASF were comparable with those released by dendritic cells and airway epithelial cells, which express biologically functional amounts of BAFF upon TLR3 triggering.37,–,39 Similarly, in this work we provided the first demonstration that RASF constitutively release high levels of soluble APRIL. Although, TLR3 activation did not induce a large increase in the amount of APRIL produced by RASF, APRIL was released at higher rate within the first 24 h, possibly reflecting a role for TLR3 in the early mobilisation of intracellular APRIL. Furthermore, APRIL was not homogeneously expressed among RASF, demonstrating some heterogeneity within the RASF population.
Despite RASF overexpressing TLR4,19 LPS treatment induced only limited levels of BAFF, similar to airway fibroblasts.40 These results are in keeping with previous evidence that TLR4 can even downregulate BAFF mRNA expression in RASF in the presence of interferon γ (IFNγ)21 and confirm that BAFF transcription is under the control of TRIF (Toll/IL-1R domain-containing protein inducing IFNΒ) rather than MyD88-dependent pathways. Importantly, the capacity of synoviocytes to respond to TLR3 by releasing high levels of BAFF/APRIL was not restricted to RASF as OASF displayed similar expression in resting and stimulated conditions. Conversely, RADF produced negligible amounts of BAFF/APRIL and failed to significantly increase protein expression despite similar mRNA responses to TLR3. Overall, this suggests that the overexpression of BAFF and APRIL in SF is dependent on epigenetic abnormalities related to the anatomical localisation rather than disease specificity.22 In addition, the importance of TLR3 is relevant to RA pathogenesis as RASF can be activated in the synovial microenvironment by RNA released by necrotic cells.24 Importantly, because TLR3 signals via the adaptor molecule TRIF in a MyD88-independent pathway, its activation also leads to type I IFN and IFN responsive genes expression. In this regard, increase in IFNΒ, which is known to be upregulated in the RA synovium,41 might contribute to the pathogenesis of RA by exacerbating Th17, as recently suggested.42
RASF were extremely efficient compared with RADF in inducing the prompt and sustained production of class-switched antibodies, primarily IgG and to a lesser extent IgA from unswitched IgD+ B cells. Stimulation of RASF (and OASF), but not RADF, with TLR3 ligands strongly enhanced the production of IgG and IgA in co-cultured B cells, an effect directly dependent on RASF as IgD+ B cells alone did not respond to PIC.
The process of isotype class-switching requires AID and occurs via excision of switch circles following the introduction of double strand DNA breaks in the Ig switch regions.43 Accordingly, we showed that RASF, but not RADF, induced a rapid and sustained de novo expression of AID in co-cultured IgD+ B cells, an effect significantly potentiated by TLR3 simulation. Subsequently, we demonstrated that AID expression induced Iγ-Cμ CTs, which are a sensitive marker of class-switching, being transiently produced for up to 48 h after CSR.27 Thus, increased detection of Iγ-Cμ CTs after over 72 h of co-culture demonstrated that class-switching from IgM to IgG was actively ongoing and that TLR3 stimulation of RASF potentiated CSR in IgD+ B cells. Accordingly, the IgG:IgM ratio was over 10-fold higher 8 days after co-culture in TLR3-stimulated conditions than after 24 h. Of relevance, TLR3-induced RASF-dependent B cells activation was maintained when B cells and RASF were separated via a cell insert, suggesting that soluble factors are sufficient to induce B cell activation. This evidence is of particular importance as BAFF can elicit its biological functions both in a membrane-bound and in a soluble form.44
BAFF and APRIL are critically involved in promoting functional B cell activation, AID expression and Ig class-switching;16 45 thus, we finally questioned whether RASF-derived BAFF and APRIL were required to mediate these processes. We showed that TLR3-dependent IgG and IgA production was completely inhibited after interfering with signalling through both BAFFR and BCMA on the B cell surface. Of relevance, only minimal effects on IgM production were observed, suggesting that reduced B cell activation rather than survival was responsible for the downregulation of IgG/IgA. Accordingly, BAFFR/BCMA blockade also downregulated the expression of AID and the production of Iγ-Cμ CTs. These in vitro data support in vivo observations in the RA/SCID model, in which functional niches of autoreactive B cells required BAFF and APRIL for Ig production,8 AID expression, ongoing class-switching and production of ACPA.8
In summary, in this study we report that: (1) RASF, but not RADF, constitutively release high levels of BAFF and APRIL; (2) BAFF, and to a lesser extent APRIL, are induced by TLR3 ligands; (3) TLR3 stimulation enhances the capacity of RASF to promote AID expression, isotype class-switching and IgG/IgA production in unswitched IgD+ B cells; and (4) these effects can be efficiently blocked by inhibition of BAFF/APRIL signalling through their receptors on the B cell membrane. Overall, this work contributes to elucidate the mechanisms regulating the crosstalk between stromal and B cells in the synovial microenvironment and highlights the importance of the BAFF/APRIL system in sustaining the functionality of synovial niches of autoreactive B cells, with potential implications for biological treatments targeting B cell-related pathways in RA.
Acknowledgments
We would like to thank Dr Taher Taher and Dr Mohey Eldin El Shikh at QMUL for their help with FACS analysis and confocal microscopy, respectively. This work was funded by the Arthritis Research UK project grant 18399 to MB, IM and CP and by FP 7 Masterswitch and IAR Epalinges CH to FB, DK and SG.
References
Supplementary materials
Web Only Data
Files in this Data Supplement:
Footnotes
MB and NWK contributed equally to this work.
-
Funding Arthritis Research UK.
-
Competing interests None.
-
Ethics approval This study was conducted with the approval of the East London & The City Research Ethics Committee 3 (LREC07/Q0605/29).
-
Provenance and peer review Not commissioned; externally peer reviewed.