Article Text

Abstract
CTLA-4 is a regulator of co-stimulation and inhibits the activation of T cells through interfering with the interaction of CD80/86 on antigen-presenting cells with CD28 on T cells. CTLA-4 binds to the surface of antigen-presenting cells, such as dendritic cells and monocytes through CD80/86. Monocytes can differentiate in osteoclasts, the primary bone resorbing cells. Herein, we investigated whether the binding of CTLA-4 affects the differentiation of monocytes into osteoclasts in vitro and vivo. We show that CTLA-4 dose-dependently inhibits RANKL- as well as tumour necrosis factor (TNF)-mediated osteoclastogenesis in vitro without the presence of T cells. Furthermore, CTLA-4 was effective in inhibiting TNF-induced osteoclast formation in a non-T cell dependent TNF-induced model of arthritis as well as the formation of inflammatory bone erosion in vivo. These data suggest that CTLA-4 is an anti-osteoclastogenic molecule that directly binds osteoclast precursor cells and inhibits their differentiation. These findings are an attractive explanation for the anti-erosive effect of abatacept, a CTLA-4 immunoglobulin fusion protein used for the treatment of rheumatoid arthritis.
Statistics from Altmetric.com
CTLA-4 (CD152) is a surface protein on T lymphocytes, which negatively regulates the co-stimulation process between antigen-presenting cells (APCs) and T cells.1–3 Co-stimulation is the second essential signal for T cell activation apart antigen presentation through the T cell receptor. Lack of the co-stimulation does not allow T cell activation and rather promotes the inverse reaction, T cell anergy.4 CTLA-4 competes for the binding of CD28 on T cells with the co-stimulation molecules B7-1 (CD80) and B7-2 (CD86) on APCs. Owing to a more than 10-fold higher affinity for B7-1 and B7-2, CTLA-4 disrupts the co-stimulation signal for T cells and prevents a persistent and pathological T cell activation. This regulatory effect has provided the rationale for using CTLA-4 as an immunomodulatory drug. Administration of CTLA-4 thus affects a variety of T cell dependent animal models of autoimmune disease, such as experimental autoimmune encephalomyelitis, collagen-induced arthritis and diabetes mellitus.5–7 Moreover, CTLA-4 inhibits the signs and symptoms of human rheumatoid arthritis and has recently been approved as a therapy for rheumatoid arthritis refractory to other disease-modifying drugs.8–10
Current concepts suggest that the inhibition of co-stimulation of T cells is the key mode of action of CTLA-4. However, additional modes of action could be based on the fact that CTLA-4 directly binds to APCs and modulates their function. Thus, CTLA-4 might not only indirectly affect T cells through disrupting their interaction with APCs, but may also directly affect its primary cellular target, which is the cell of the monocytic lineage expressing its ligands B7-1 (CD80) and B7-2 (CD86). Macrophages are not only one of the major cytokine-producing cells at sites of inflammation but they also differentiate into bone-resorbing osteoclasts if these sites are closely located to mineralised tissue such as bone.11 In experimental arthritis as well as human rheumatoid arthritis, formation of osteoclasts within the inflamed joint is a key step for inflammatory bone erosion and joint damage.12–14 CTLA-4 binding to monocytes/macrophages could thus affect their differentiation into osteoclasts and directly inhibit inflammatory bone loss in addition to affecting T cell function.
MATERIALS AND METHODS
Fluorescence-activated cell sorter (FACS) analysis
For assessing the binding of CTLA-4 (kindly provided by Bristol Myers Squibb, New York, USA) to osteoclast precursor cells, murine peripheral blood mononuclear cells were labelled with an antibody against CD11b (BD Biosciences, Heidelberg, Germany) as well as fluorescein isothiocyanate (FITC)-conjugated CTLA-4. FITC conjugation of murine and human CTLA-4 immunoglobulin was done by using the FluoroTag FITC conjugation kit (purchased from Sigma, Munich, Germany) according to the manufacturer’s instructions. After passage through a gel filtration column to remove unbound FITC, the protein was diluted to 1 mg/ml with phosphate-buffered saline. Binding of human FITC-labelled CTLA-4 was evaluated on human peripheral blood mononuclear cells. FITC labelling of human CTLA-4 immunoglobulin was done according to the method described above. Human cells were additionally stained with anti-CD14 antibody (R&D, Wiesbaden-Nordenstadt, Germany) and an isotype control, FACS analysis was performed according to standard settings.
Osteoclast differentiation and bone resorption assay
Murine osteoclasts were generated as previously described.15 In brief, spleen cells from 6-week-old wild-type mice and human tumour necrosis factor (TNF) transgenic (hTNFtg) mice were cultured overnight in αMEM containing 10% fetal bovine serum and 30 ng/ml macrophage colony stimulating factor (M-CSF). After 24 h, non-adherent cells were harvested and subjected to gradient purification, seeded in 48-well plates (1×106 cells/ml) and supplemented with 30 ng/ml M-CSF and 50 ng/ml RANKL (both R&D). In the TNFα-stimulated osteoclast cultures, soluble murine TNFα was added in a concentration of 10 ng/ml (R&D). CTLA-4 immunoglobulin was added at various doses ranging from 1 to 100 μg/ml. Tartrate-resistant acid phosphatase (TRAP) staining was performed after 5 days and osteoclasts were identified by the presence of three or more nuclei and purple colour. Osteoclast precursors were identified as TRAP-positive cells with one or two nuclei. For bone resorption assay a similar protocol was applied but cells were cultured on a BioCoat Osteologic Bone Cell Culture System (from BD) for 6 days as previously described,15 allowing to visualise resorption pits. Number and area of resorption pits was counted.
Human osteoclast assay
Human peripheral blood mononuclear cells were harvested and CD14 expressing cells were positively selected via a magnetic cell sorter. Monocytes were then cultured in αMEM medium containing 10% fetal calf serum, supplemented with 25 ng/ml M-CSF and 50 ng/ml RANKL (both from Peprotech, Hamburg, Germany) as well as 5 ng/ml transforming growth factor-β1 (from R&D) over 17 days. Human CTLA-4 immunoglobulin was added at the same concentrations as in the murine culture system. Medium, differentiation factors and CTLA-4 immunoglobulin were changed every 72 h.
Animal procedures
Murine CTLA-4 immunoglobulin (5 mg/kg) was daily injected into the peritoneal cavity of 6-week-old hTNFtg mice (strain Tg197; genetic background: C57BL/6;16). Control groups receiving phosphate-buffered saline or infliximab (10 mg/kg three times weekly). Treatment lasted for 4 weeks and each group comprised 10 animals. The local ethics committee approved all animal procedures.
Clinical assessments
Scoring of clinical signs of arthritis and measurement of body weight was done in all mice once weekly starting from 5 weeks of age. Arthritis was evaluated in a blinded manner as described previously.15 Briefly, joint swelling was examined in all four paws, and a clinical score of 0–3 was assigned (0 = no swelling, 1 = mild, 2 = moderate, 3 = severe swelling of the toes and ankle). In addition, grip strength was examined in each paw, using a 3 mm diameter wire, and was scored on a scale of 0 to −4 (0 = normal grip strength, −1 = mildly reduced, −2 = moderately reduced, −3 = severely reduced, −4 = no grip strength).
Joint histology
Hind paws were fixed overnight in 4.0% formalin and then decalcified in 14% EDTA (Sigma, St Louis, Missouri, USA) until the bones were pliable. Serial paraffin sections (2 μm) of hind paws were stained with haematoxylin and eosin for the assessment of synovial inflammation and bone erosion, toluidine blue for proteoglycan loss of articular cartilage, and TRAP for detection of osteoclasts. TRAP staining was performed using a leucocyte acid phosphatase staining kit (Sigma-Aldrich, St Louis, Missouri, USA). Synovial inflammation, bone erosions, osteoclast numbers and cartilage destruction were quantified with the use of a Zeiss Axioskop 2 microscope (Zeiss, Marburg, Germany) equipped with a digital camera and image analysis system (Osteomeasure, OsteoMetrics, Decatur, Georgia, USA), as described previously.15 The area of inflammation was quantified on haematoxylin and eosin stained sections. Total scores were calculated as the sum of the areas of inflammation in all tarsal joints in each mouse. Erosions were quantified in the same haematoxylin and eosin stained sections. Osteoclasts (three or more nuclei, TRAP-positive) were assessed in TRAP-stained serial sections. For proteoglycan content analysis, toluidine blue sections were used. Negatively stained cartilage appears as light blue as opposed to proteoglycan-rich dark blue stained cartilage. The area of negatively stained and total cartilage was measured, calculated and given as the percentage cartilage that contains no proteoglycan.
Immunohistochemistry
For immunohistochemistry, deparaffinised, ethanol-dehydrated tissue sections were stained with monoclonal antibodies against surface markers of neutrophil granulocytes (MCA771G; clone 7-4; Serotec, Raleigh, North Carolina, USA), macrophages (anti-F4/80; clone Cl:A3-1; Serotec), T lymphocytes (anti-CD3; CD3-12; Novocastra, Newcastle upon Tyne, UK) and B lymphocytes (anti-B220; clone RA3-6B2; Pharmingen, Leiden, the Netherlands) as well as polyclonal antibody against interleukin 1 (R&D, Minneapolis, Minnesota, USA). Sections were pre-treated with proteinase K (Roche, Mannheim, Germany; 0.05% for 5 min at 37°C) for detection of neutrophils, macrophages and T lymphocytes, or heat (95°C, 20 min) for interleukin 1 or were left without pre-treatment for staining of B lymphocytes. All above-mentioned procedures were followed by blocking of endogenous peroxidase and subsequent incubation with a biotinylated species-specific anti-IgG secondary antibody. Sections were incubated with the appropriate ABC-complex using 3,3'-diaminobenzidine (Sigma-Aldrich) as chromogen, resulting in brown staining of antigen-expressing cells.
RESULTS
CTLA-4 binds to murine and osteoclast precursors
To address whether CTLA-4 binds to monocytic osteoclast precursors we performed FACS analyses with labelled murine CTLA-4 (fig 1A). Whereas virtually all CD11b-negative cells did not show binding of CTLA-4, CD11b-positive cells, which serve as osteoclast precursors17 showed a strong binding to labelled CTLA-4. Similarly, FITC-labelled human CTLA-4 (abatacept) showed strong binding to human CD14-positive monocytes. Thus, more than 60% of peripheral monocytes showed a positive binding of CTLA-4 in the FACS analysis (fig 1B).

CTLA-4 blocks osteoclast formation in vitro
We next investigated whether CTLA-4 affects the differentiation of osteoclast from monocyte precursors in vitro. Monocytes differentiate into osteoclasts when stimulated with M-CSF and RANKL. Addition of CTLA-4 dose-dependently decreases osteoclast formation. Thus the number of TRAP-positive mononuclear osteoclast precursors as well as the number of multinucleated osteoclasts was significantly lower when CTLA-4 was added, indicating that CTLA-4 directly inhibits osteoclast formation (fig 2A). TNF is a potent stimulator of osteoclast formation and monocytes from hTNFtg mice thus show an increased potential to differentiate into osteoclasts. Addition of CTLA-4 to monocytes from hTNFtg mice also dose-dependently blocked their differentiation into the osteoclast lineage, suggesting that CTLA-4 also affects inflammatory osteoclast formation (fig 2B). In line, CTLA-4 also dose-dependently blocked osteoclast formation, when cultures were stimulated by exogenous TNF-α (fig 2C).

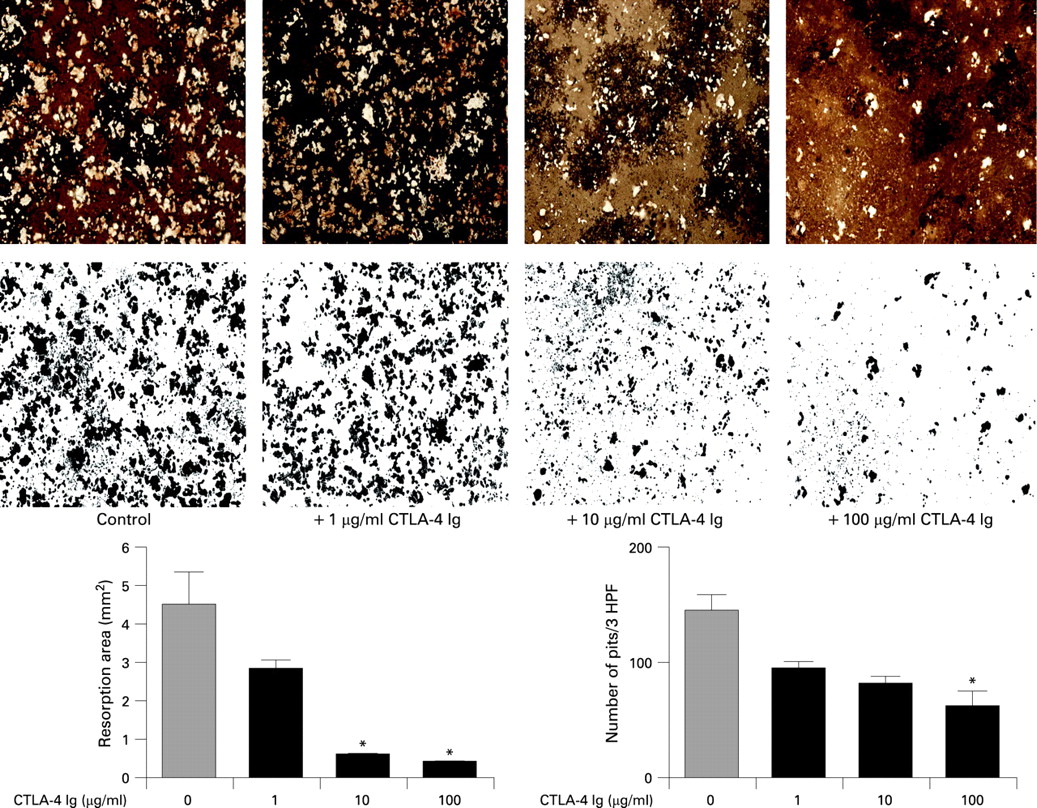
CTLA-4 blocks bone resorption in vitro
To assess the functional ability of the cultivated osteoclasts and to depict their inhibited effect on bone resorption we quantified the area of bone resorption and the number of resorption holes in the Osteologic Bone Cell Culture System. A significant dose-dependent reduction in the area and number of resorption holes was found, when CTLA-4 was added to the cultures, suggesting that the impact of CTLA-4 on osteoclast differentiation also has functional consequences by reducing bone resorption (fig 3). Thus, bone resorption area was strongly reduced by CTLA-4 to only about 10% of the size without any addition of the compound, suggesting a highly effective blockade of bone resorption.
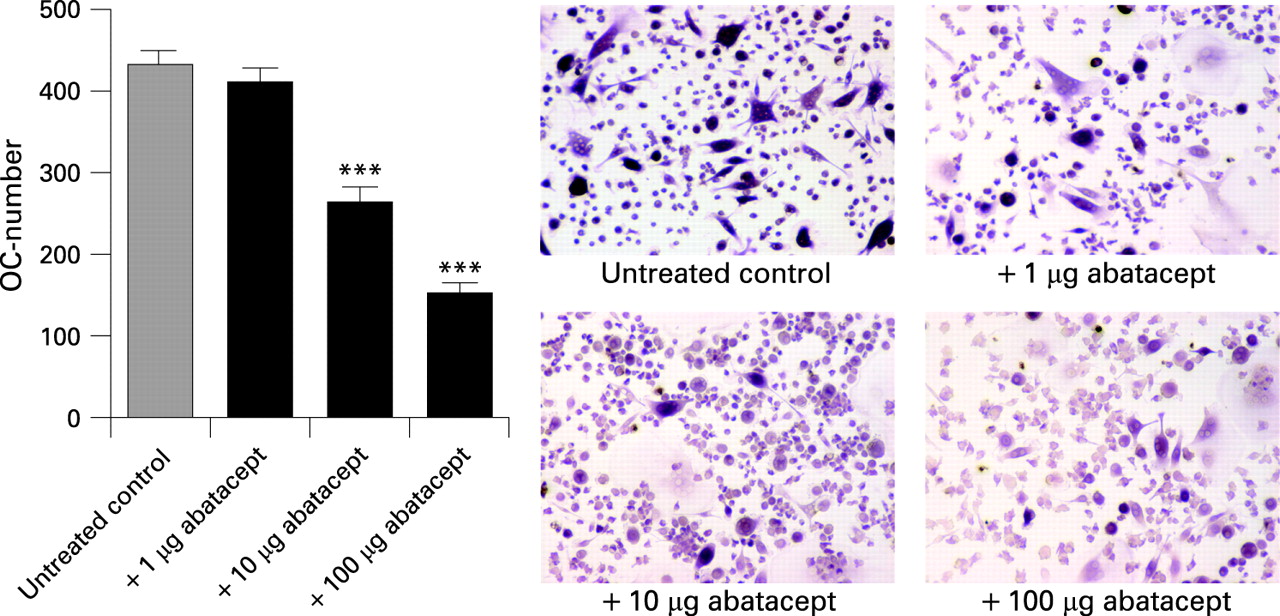
CTLA-4 blocks human osteoclast formation
We next addressed, whether CTLA-4 also inhibits the formation of human osteoclasts by impairing their differentiation from precursor cells. Human osteoclasts can be differentiated from peripheral blood mononuclear cells by addition of M-CSF and RANKL (fig 4). This procedure yields formation of TRAP-positive multinucleated osteoclasts. Addition of human CTLA-4 was again effective in inhibiting the differentiation of osteoclasts in a dose-dependent manner, indicating that the direct inhibitory effect of human CTLA-4 holds true for the formation of human osteoclasts.
Tumour necrosis factor-mediated joint inflammation is not affected by CTLA-4
To investigate whether CTLA-4 directly affects osteoclast formation in vivo, we examined the effects of co-stimulation blockade with CTLA-4 immunoglobulin in hTNFtg mice, an experimental model of arthritis. This model is considered as T cell independent and shows a highly destructive phenotype, which is based on enhanced osteoclast formation in the joint due to overexpression of TNF. Taking into consideration that inflammatory osteoclast formation in vivo may indirectly depend on the severity and cellular composition of synovial inflammation, we found it important to address whether CTLA-4 affects synovial inflammation. We used several methods to address the effects of CTLA-4 on TNF-mediated synovial inflammation, which included: (a) the clinical signs of arthritis; (b) the histological signs of synovial inflammation; (c) the immune phenotyping of cells invading the inflamed joint; (d) cytokine expression in the synovial membrane; as well as (e) autoantibody formation during arthritis. There was no difference among CTLA-4 and vehicle treatment with respect to clinical signs of arthritis, such as paw swelling, grip strength and body weight (fig 5A–C). Thus paw swelling steadily increased in both groups, whereas grip strength of paws as well as body weight continuously decreased. Moreover, histological scores for inflammation were similar among the two groups (fig 5D). Thus, synovial hyperplasia, influx of inflammatory cells and pannus formation was equally pronounced in hTNFtg mice treated with CTLA-4 and vehicle. Distribution of macrophages, neutrophils, T cells and B cells were similar among the two groups (fig 5E). Whereas macrophages, neutrophils and a few T cells accumulated in the inflamed synovial membrane, B cell aggregates were found in the bone marrow adjacent to inflamed joints.

CTLA-4 blocks inflammatory osteoclast formation in vivo
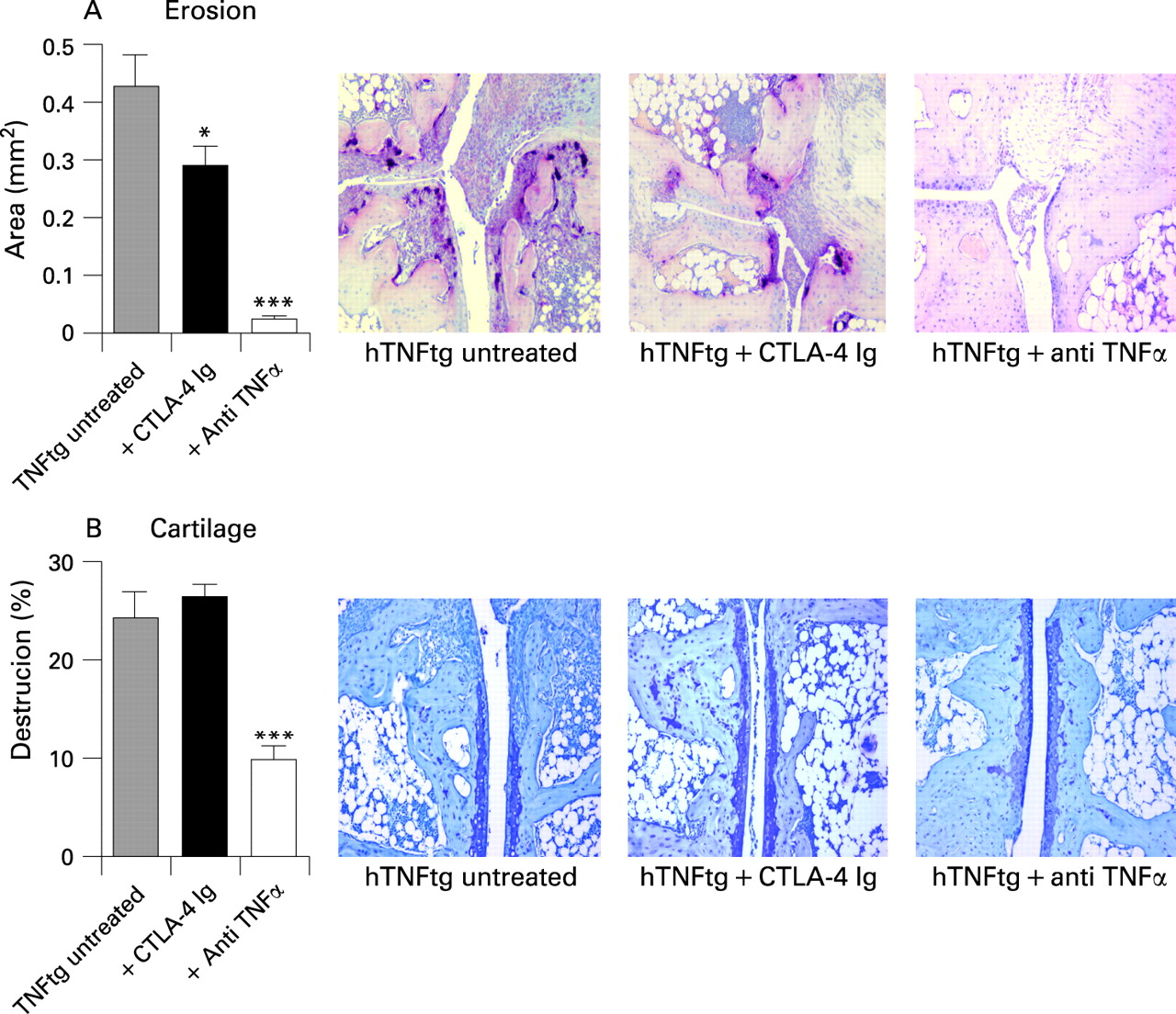
We next addressed that CTLA-4 might impair inflammatory osteoclast formation in vivo, which is followed by less pronounced bone damage of the inflamed joint. When investigating the number of osteoclasts formed in the joints of hTNFtg mice treated with CTLA-4 we found significantly fewer osteoclasts compared with vehicle-treated mice. Furthermore, the formation of osteoclast precursor cells was also impaired upon CTLA-4 treatment. When assessing bone resorption it was evident that CTLA-4 inhibited bone erosions in hTNFtg mice, which is well in line with its inhibitory function on osteoclast formation (fig 6A). In contrast cartilage proteoglycan loss, which does not depend on osteoclasts and is closely linked to synovial inflammation, was unchanged (fig 6B).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
CTLA-4 is a key molecular regulator of co-stimulation that prevents a sustained interaction between CD28 on the surface of T cells and CD80/CD86 on the surface of APCs. This allows fine tuning of T cell–APC interaction, which is required for the proper functioning of the immune system. Deficiency of CTL-4 results in autoimmune inflammatory disease in animals as well as in humans.18 Current concepts have highlighted the role of CTLA-4 as a regulator of T cell activation and have favoured a more or less unidirectional view with the APC as the donor of the signal and the T cell as the recipient. Immunomodulatory therapy with CTLA-4 immunoglobulin is thus preferentially considered as an intervention, which targets the activation of T lymphocytes.
Recent concepts, one of them being reverse signalling, however, have refined the general concepts of cell–cell interaction and have outlined that these interactions can be rather bidirectional, which means that the “ligand”-expressing cell undergoes a functional change upon engaging with the receptor.19 In the case of CTLA-4 this would mean that binding of this molecule leads to a functional change of the APC. Indeed, it has been shown that regulatory T cells, which express CTLA-4 “retrogradely”, influence APCs, such as dendritic cells, by affecting tryptophan catabolism.20 At a molecular level, CTLA-4 activates tryptophan hydroxylase 1 in APCs, which is the key enzyme for degradation of tryptophan. Similar mechanisms may be considered to explain the inhibitory effect of CLTA-4 on osteoclast differentiation, although tryptophan hydroxylase 1 appears not to be the molecular mechanism in osteoclasts as we were not able to find a major change in its expression in osteoclast precursor cells challenged by CTLA-4.
Although osteoclasts are not considered as APCs they share some similarities with them. Similar to dendritic cells and macrophages, osteoclasts derive from monocytic precursor cells and differentiation into the various lineages is dependent on the cytokine respective milieu. Thus, granulocyte M-CSF and interleukin 4, for instance, support differentiation of dendritic cells but suppress osteoclast formation; however, other cytokines such as M-CSF and RANKL are essential for osteoclast differentiation.21–24 Osteoclasts, such as dendritic cells, are interaction partners for T cells and provide signals that are instrumental for their differentiation. Molecular receptor–ligand partners such as RANKL-RANK are shared in T cell–dendritic cell interaction as well as T cell–osteoclast interactions. Considering these similarities we hypothesised that CTLA-4 could influence cells from the osteoclast lineage.
In our experiments CTLA-4 emerges as a potent negative regulator for osteoclasts in vitro and in vivo. Osteoclasts can be raised from mononuclear cells just by the addition of M-CSF and RANKL and without the presence of T cells. The inhibitory effect of exogenous CTLA-4 on to the osteoclast lineage was a direct effect that did not require the presence of T cells in the culture system. This suggests that CTLA-4 directly blocks osteoclast differentiation independent of T cells. Moreover, this inhibitory effect also applies for inflammatory osteoclastogenesis, as osteoclast differentiation was also impaired in the presence of TNF-α. The effect of CTLA-4 on osteoclast also hold true in vivo, showing an inhibition of osteoclast formation in arthritic joints, which is accompanied by less severe bone destruction. As T cells are not considered as a major trigger of arthritis in the hTNFtg mouse model and only a few T lymphocytes are found in the synovium of these mice, the effect of CTLA-4 on osteoclast formation appears to be a direct one affecting the differentiation process of monocytic cells into osteoclasts rather than indirectly inhibiting T cell activation.
Treatment with abatacept, a recombinant fusion protein of CTLA-4 and the Fc portion of immunoglobulin has proven efficacy in controlling the signs and symptoms of rheumatoid arthritis.6–8 The mode of function of abatacept is the inhibition of T cell co-stimulation and T cell activation. However, it should be considered that the primary cellular target of abatacept are mononuclear APCs that express its ligands, CD80 and CD86. Direct effects on mononuclear effects and negative regulation of osteoclast differentiation is an elegant explanation for the bone-sparing effect of abatacept, which has been demonstrated in clinical studies in patients with rheumatoid arthritis, showing retardation of structural joint damage.9 10
Acknowledgments
We thank Birgit Türk for excellent technical assistance and George Kollias (Alexander Fleming Biomedical Sciences Research Center, Vari, Greece) for hTNFtg mice. This work was supported by the START prize of the Austrian Science Fund (to GS), the Interdisciplinary Center for Clinical Research at the University of Erlangen (IZKF project C5; to GS and JZ) and Deutsche Forschungsgemeinschaft Grant FOR 661 (to GS).
REFERENCES
Footnotes
Competing interests: None.