Article Text

Abstract
Objective: To investigate the potential role of interleukin (IL) 27 in rheumatoid arthritis (RA) by examining the expression of IL27 in the articular joints of patients with RA and the effect of recombinant IL27 in vivo in a murine model of collagen-induced arthritis (CIA).
Methods: Synovial membranes from patients with RA were examined for the presence of IL27 by immunohistochemistry and by western blot. Mice developing CIA were treated with IL27 and the ensuing disease progression and immunological profile determined. The effect of IL27 on T-cell response in vitro was also ascertained.
Results: IL27 was clearly detected in the RA synovial membranes. Short-term administration of IL27 at the onset of the disease significantly attenuated disease severity compared with untreated controls. Histological examination showed that while untreated mice developed severe cellular infiltration in the joints, synovial hyperplasia and joint erosion, this pathology was profoundly reduced in IL27-treated animals. Treatment of mice with IL27 also decreased the amounts of serum IL6 and collagen-specific IgG2a. Spleen and lymph node cells from the IL27-treated mice produced significantly less interferon γ and IL17 than cells from the control mice when cultured with collagen in vitro.
Conclusion: These results demonstrate that IL27 may be a potential therapeutic agent against RA at the onset of the disease.
Statistics from Altmetric.com
Production of raised levels of proinflammatory cytokines characterises rheumatoid arthritis (RA) synovial inflammation.1 Moreover, successful therapeutic targeting of cytokines in RA, particularly tumour necrosis factor α (TNFα) has demonstrated their critical pathogenic importance. Th1 responses were previously considered to predominate within RA synovial T-cell subsets and to contribute significantly to dysregulated cytokine production.2–4 However, recent studies demonstrate that interleukin (IL)17-producing CD4+ T cells (Th17), and not interferon γ positive (IFNγ+) CD4+ effector Th1 cells, play a predominant pathogenic role in RA.5 6 IL17 is implicated in disease pathogenesis through enhanced expression of TNFα and IL1β by monocytes, synergistic effects on TNFα-induced IL1, IL6 and IL8 synthesis by synovial fibroblasts, and increased neutrophil activation.7 8 IL17 is also directly implicated in cartilage damage and bone erosion.9 10 IL17 neutralisation using soluble receptors or neutralising antibodies suppresses rodent arthritis and manifests in both reduced synovial infiltration and articular destruction.5 11 Moreover, IL17-deficient mice exhibit reduced articular inflammation. One key question arises, how is Th17 regulated in a proinflammatory environment? We report here that IL27 can modulate arthritic inflammation.
IL27, a member of the IL12 family, is a heterodimeric cytokine consisting of an IL12p40-related protein, EBI3 and a unique IL12p35-like protein p28.12 The role of IL27 in the regulation of inflammatory diseases is currently controversial. Early studies showed that mice lacking IL27 receptor (IL27R) had reduced Th1-type responses in a variety of in vitro and in vivo assays, suggesting that IL27 was directly involved in Th-cell differentiation.13 14 Consistent with these reports are the findings that the severity of adjuvant-induced arthritis in rats and experimental autoimmune encephalomyelitis in mice can be abolished by anti-IL27 antibodies.15 16 IL27 neutralisation in one study of rodent adjuvant arthritis suggested suppression of inflammation. In contrast, other studies demonstrated that mice lacking IL27R developed raised Th17 and enhanced central nervous system inflammation when infected with Toxoplasma gondii or used for experimental autoimmune encephalomyelitis, implying that IL27 was an antagonist of Th17 activity.17 18 Consistent with this finding, IL27 was shown to inhibit the development of Th17 in vitro. Thus it was postulated that IL27 can induce Th1-cell differentiation on naïve CD4+ T cells but can also suppress the proinflammatory cytokine production such as IL17.19
We have therefore investigated the potential role for IL27 in inflammatory arthritis. We detected IL27 expression in human RA tissues. Thereafter we investigated the potential functional implications of this expression using the murine collagen-induced arthritis (CIA) model. We found that IL27 could attenuate CIA when administered at the onset of articular disease. Reduced disease development was associated with downregulation of ex vivo IL17 and IL6 synthesis. Our data therefore suggest that IL27 may be an effective therapeutic agent against RA.
MATERIALS AND METHODS
Detection of IL27 in RA synovial tissue
Synovial tissue samples from patients diagnosed with RA were obtained from the tissue bank of the Centre of Rheumatic Diseases, Royal Infirmary, Glasgow. The study was approved by local ethical committees and informed consent was obtained from every patient. For immunohistochemistry, synovial membrane tissues were fixed in formalin for 24 h and then embedded in paraffin. Serial sections (5 µm) were stained with a polyclonal goat anti-human IL27p28 (R&D Systems, Abingdon, UK). Endogenous peroxidase was blocked by applying methanol containing 3% hydrogen peroxide (Sigma-Aldrich, Poole, UK). Non-specific protein binding was blocked with a 20% horse serum/20% human serum phosphate-buffered saline (PBS) solution. The sections were incubated with primary antibody (0.5 μg/ml in 5% horse serum/PBS) overnight. Sections were then incubated for 1 h with biotinylated anti-goat antibody (Vector, Burlingame, California, USA) followed by peroxidase-conjugated avidin–biotin complex (ABC Kit; Vector). They were then counterstained with haematoxylin (Sigma-Aldrich). Negative controls were stained with a normal goat IgG fraction at the same concentration as the primary antibody. For western blot, proteins were extracted from the synovial tissue samples using the Triazol (GIBCO, Paisley, UK) reagent, according to the manufacturer’s instructions. Protein extracts were resuspended with sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) sample buffer (Invitrogen, Paisley, UK). The BCA protein assay (Pierce, Illinois, USA) was used to calculate the concentration of each sample. Protein extracts (40 μg of protein per sample) were separated by SDS-(4–12%) PAGE, under reducing conditions. The Invitrogen western blot kit was used for western blot analysis of the RA synovial membrane protein extracts, according to the manufacturers instructions.
Animals and reagents
Male DBA/1 mice obtained from Harlan Olac (Bicester, UK) were used at 8–10 weeks old and maintained at the Biological Services Facilities, University of Glasgow. All animal experiments conducted in this study were carried out in accordance with the Home Office, UK animal guidelines.
Gene expression and protein purification of IL23-Fc and IL27-Fc
IL23 and IL27 were cloned, expressed and purified as previously described.12 20 A mammalian cell expression vector (pS-L-Fc) was constructed to fuse two proteins, linked with a linker peptide (3×GGGGS), in-frame with the human IgG1 Fc region. Briefly, mouse IL12p40 and EBI3 cDNA PCR fragments were amplified from a lipopolysaccharide-activated J774 cell cDNA library, using primer sequences as follows: IL12p40 sense, 5′- TCA TGG ATC CGT GGG AGC TGG AGA AAG ACG TTT; anti-sense, 5′- CGG CGG ATC CGG ATC GGA CCC TGC AGG GAA CAC; EBI3 sense, 5′- CCC CGG ATC CCA CTG AAA CAG CTC TCG TGG CTC T; anti-sense, 5′- CGG GAT CCC TTA TGG GGT GCA CTT TCT ACT TGC C. A BamHI site was introduced in both pairs of primers. The PCR fragments were then inserted into the BamHI site of the pS-L-Fc vector in the right orientation to form p40-L-Fc and pEBI3-L-Fc vectors.
IL23p19 and IL27p28 PCR fragments were also amplified from the same cDNA library with primer sequences as follows: IL23p19 sense, 5′- TCA GGA ATT CGC TGT GCC TAG GAG TAG CAG TC; anti-sense, 5′- TTA AGC GGC CGC CAC TAA TGG CTC AGT CAG AG; IL27p28 sense, 5′- CCT GGC GGC CGC CAA CAG ACC CCC TGA GCC TTC A; anti-sense, 5′- GCC TGC GGC CGC GGG CAG GGA CAG CAG CAG CAG GTC. The EcoRI and NotI sites were introduced in the 5′ ends of IL23p19 sense and anti-sense primers, respectively. The NotI site was introduced in both IL27p28 sense and anti-sense primers. An IL23p19 PCR fragment was inserted into EcoRI, NotI sites downstream of linker sequence of the p40-L-Fc vector, producing IL23-Fc (p40-L-p19-Fc), while an IL27p28 PCR fragment was inserted into the NotI site of the pEBI3-L-Fc vector to generate IL27-Fc (EBI3-L-p28-Fc) expression construct. The reading frames of IL23-Fc and IL27-Fc were sequenced to confirm that no signal mutations had occurred during PCR amplification. The IL23-Fc and IL27-Fc expression constructs were then transfected into Chinese hamster ovary cells. The permanently transfected cell lines were selected using 700 μg/ml Zeocin for 10 days’ culture. The highest expression clones for both IL23-Fc and IL27-Fc were screened using a specific human IgG1 Fc ELISA with purified rabbit anti-human IgG1 (Sigma) as capture antibody and biotinylated monoclonal anti-human Fc specific antibody as detecting antibody (DakoCytomation, Denmark). Recombinant proteins were purified with a protein A agarose 4B column (Amersham, UK). The molecular weight and purity of recombinant proteins were examined by SDS-PAGE and Coomassie blue gel staining (Invitrogen). IL23 and IL27 were also obtained from R&D Systems. IL23 and IL27 produced in our laboratory had comparable activity to the commercially obtained cytokines.
Induction of CIA and assessment of arthritis
CIA was elicited in mice as previously described.21 Briefly, mice were immunised by intradermal injection of 200 µg of acidified bovine type II collagen (CII; Sigma) emulsified in complete Freund’s adjuvant (Difco, Detroit, Michigan, USA). Mice were boosted intraperitoneally with CII (200 µg in PBS) on day 21. Mice were monitored for signs of arthritis as previously described.21 Scores were assigned based on erythema, swelling or loss of function present in each paw on a scale of 0–3, giving a maximum score of 12 for each mouse. Paw thickness was measured with a dial calliper (Kroeplin, Munich, Germany). For histological assessment, mice were killed and the hind limbs removed, fixed in 10% neutral-buffered formalin and 5 µm sections were stained with H&E or toluidine blue (both Sigma). The quantification of arthritis was by an observer “blinded” to treatment and a score was assigned to each joint based on the degree of inflammation, synovial hyperplasia and erosion, as described previously.22
Administration of IL23 and IL27
To investigate the effect of IL27 in murine CIA, DBA/1 mice were injected intraperitoneally daily from day 21 for 10 days with IL27 (2 µg/mouse). Control mice received similar amounts of IL23 or PBS.
Collagen-specific in vitro culture
Spleens were removed on day 32 after primary immunisation. Single-cell suspensions were prepared and cultured in triplicate at 2×106 cells/ml in RPMI 1640 supplement with 100 IU/ml penicillin, 100 µg/ml streptomycin, 25 mM HEPES buffer and 10% fetal calf serum (all Invitrogen, Paisley, UK) at 37°C in 5% CO2. Cells were cultured with graded concentrations of CII in flat-bottom, 96-well plates (Nunc, Roskilde, Denmark). Supernatants were collected after 72 or 96 h and stored at −20°C until assayed for cytokine concentration. Proliferation assays were performed in parallel cultures in U-bottom, 96-well plates (Nunc) for 96 h and were pulsed with [3H]thymidine (GE Healthcare, Little Chalfont, UK) during the last 6 h of culture. Plates were then harvested and measured for incorporation of radioactivity, as previously described.21
Measurement of cytokines and anti-collagen antibody levels
All cytokines and anticollagen antibody levels were detected by ELISA. The antibody pairs for TNFα, IFNγ, IL5, IL6 and IL10 were obtained from BD Pharmingen (San Diego, California, USA) and assays were performed according to manufacturer’s instructions. Detection limits were as follows: IL5, IL6 and TNFα all at 10 pg/ml; IL10 and IFNγ both at 20 pg/ml. Sensitivity of the assay was 50 pg/ml. Serum anti-collagen II antibody titres of individual serum were detected with biotin-conjugated anti-mouse IgG1 or IgG2a (BD Pharmingen), followed by conjugated avidin peroxidase (Sigma) and developed with tetramethylbenzidine substrate (Kirkegaard and Perry, Gaithersbury, Maryland, USA).
Polarisation of Th17 cells and the effect of IL27 in vitro
CD4+ T cells were purified from spleen and lymph nodes (LN) of naïve mice and then separated into CD25+ and CD25− fractions using magnetic-activated cell sorting (Miltenyi Biotec, Bisley, UK) with purity routinely >95%. Cells were labelled with CFSE (5,6-carboxyfluorescein diacetate, succinimidyl ester; Invitrogen (Molecular Probe), Paisley, UK) and cultured with soluble anti-CD3 antibody and mitomycin-C-treated antigen-presenting cells, transforming growth factor β, IL6 and IL1β, as previously described.23 IL27 (10 or 50 ng/ml) was added at the beginning of culture. Cells were harvested at 72 h and assayed for cell division (CFSE intensity) and intracellular cytokines. Culture supernatants were analysed for cytokine concentrations by ELISA.
Intracellular staining for IL17, IFNγ and Foxp3
Single-cell suspensions were stimulated for 4 h with 50 ng/ml of phorbol myristate acetate (Sigma) and 500 ng/ml of ionomycin (Sigma). GolgiStop (BD Biosciences, Oxford, UK) was added for the final 3 h of culture. Cells were first stained extracellularly with anti-CD4, then fixed and permeabilised with Perm/Fix solution (eBioscience) and, finally, stained intracellularly with anti-IFNγ, anti-IL17 and anti-Foxp3 (all eBioscience). Directly conjugated isotype-matched rat anti-mouse antibodies (eBioscience) were used as controls for non-specific staining.
Statistical analysis
Clinical and histological scores were analysed with the non-parametric Mann–Whitney U test. Differences between cumulative incidences at a given time point were analysed by the χ2 contingency analysis. Cytokine and collagen-specific IgG levels were compared using the Student t test.
RESULTS
Detection of IL27 in the synovium of RA
We first investigated whether IL27 is present in the synovium of patients with RA. Synovial membranes (pannus) from patients with RA were stained with an anti-IL27 antibody or the proteins extracted for western blotting with IL27-specific antibody. Figure 1 shows that histological sections from patients with RA are clearly positively stained for IL27. Furthermore, IL27 (p28) was also clearly detectable in all the synovial tissues tested.

Effect of IL27 on the development of CIA
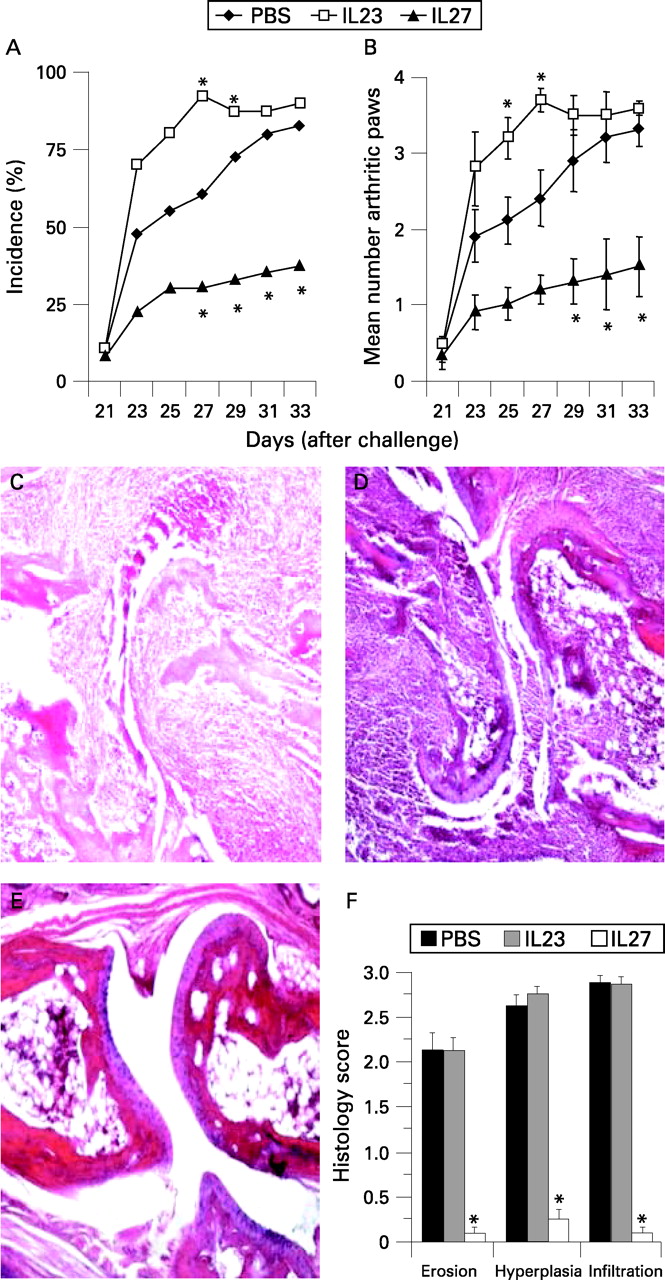
To investigate the potential role of IL27 in arthritis, we employed a murine model of collagen-induced arthritis. DBA/1 mice were primed and challenged with bovine CII, as described in “Materials and methods”. Mice began to show clinical signs of arthritis on day 23 (2 days after challenge on day 21) after priming. Mice were treated intraperitoneally for 10 days with a daily dose of IL27 from day 21, the day of CII challenge. Control mice were similarly injected with IL23 or PBS. As expected,24–26 mice treated with IL23 developed more severe disease than control mice treated with PBS. In contrast, mice treated with IL27 developed significantly reduced incidence (fig 2A) and reduced number of arthritic paws (fig 2B).

Histological analysis showed that mice treated with IL23 or PBS exhibited extensive mononuclear and polymorphonuclear cell infiltration into the joint compartment, synovial hyperplasia and adjacent cartilage, together with bone erosion (figs 2C and D). Each of these parameters was markedly suppressed in the mice treated with IL27 (fig 2E). Figure 2F summarises the histological scores. The effect of IL27 is unlikely to be due to the Fc fragment of the molecule since IL23 (which has a similar construct) exacerbated the disease. Together, these data clearly demonstrate that IL27 potently suppressed the development of CIA and such activity can prevent progression of articular damage. It should, however, be noted that when administered after the onset of disease (day 24), IL27 had little or no effect on the progression of CIA (data not shown).
Serum cytokines and anti-collagen antibody production in vivo
A potential mechanism by which IL27 could suppress CIA is by blocking proinflammatory cytokine production. Serum concentrations from arthritic mice were measured by ELISA at the end of the treatment period (day 33). There was no significant difference in the levels of IL12, TNFα and IFNγ between the IL27 and the control PBS-treated groups (data not shown). In contrast, IL27 treatment led to significant reduction in serum IL6 and IL17 compared with PBS-treated mice (fig 3A). IL4, IL5 and IL10 were not detected (data not shown). CII-specific IgG1 and IgG2a levels were also analysed by ELISA. IL27-treated mice produced significantly lower IgG2a but not IgG1 antibody (fig 3B).

IL27 suppressed CII-specific proinflammatory response in vitro
The immune response was then examined in vitro in the spleen and LN cells obtained at the end of IL27 treatment (day 33). Freshly obtained spleen and LN cells were stained for intracellular cytokines. IL27 treatment resulted in significant reduction in the percentage of cells expressing IL17 and IFNγ (fig 4A). Other cytokines were either undetected or were similar between the two groups of mice (data not shown). The LN cells were then cultured in vitro with graded concentrations of CII and cellular proliferation, and cytokine secreted into culture supernatant was analysed by ELISA. Cells from IL27-treated or PBS-treated groups proliferated equally well in response to CII (data not shown). In contrast, LN cells from IL27-treated mice produced markedly less IL17 than cells from PBS-treated control mice (fig 4B). There was no significant difference in the levels of IFNγ, TNFα or IL6 produced by the two groups (data not shown). Thus there is evidence that IL27 treatment in vivo consistently led to a decrease in the production of IL17 and also IL6 and IFNγ. The number of lymphoid cells in the arthritic paws was too few to allow estimation of the frequency of cytokine-secreting cells (data not shown).

To confirm the effect of IL27 on Th17 development, we polarised Th17 in vitro in the presence of IL27. CD4+, CD4+CD25− and CD4+CD25+ T cells were purified from spleen and LN of mice by magnetic-activated cell sorting. The cells were labelled with CFSE and cultured with anti-CD3 antibody, antigen-presenting cells, transforming growth factor β, IL6 and IL1β, as previously described.23 IL27 was added at the beginning of the culture. Cells were harvested on day 3 for intracellular staining of IL17 and Foxp3. Culture supernatant was also collected for ELISA. Figure 5 shows that IL27 markedly suppressed the development of Th17 from all three populations of CD4+ T cells. IL27 had no significant effect on T-cell proliferation (from CFSE staining) or the percentage of Foxp3+ Treg cells. The relatively low percentage of Foxp3+ cells in the CD4+CD25− population may be due to residual contamination of CD4+CD25+ T cells.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
DISCUSSION
Data reported here demonstrate that IL27 can attenuate CIA when administered at the onset of the disease. The inhibition of disease development by IL27 is likely to be due to a blocking of Th17 differentiation. This contention is borne out by our in vitro experiments, whereby we showed that IL27 markedly inhibited the induction of Th17 from naïve CD4+ T cells. IL27 also had an inhibitory effect on IL6 synthesis. This may be a consequence of the blocking of IL17, which can synergise with other cytokines for IL6 production.7 8
Our data may explain the current controversy surrounding the role of IL27 in various models of inflammatory diseases.15–19 It is generally accepted that Th17 plays a pivotal role in a variety of inflammatory conditions5 6 and that IL27 can suppress the development of Th17.17–19 We have now confirmed and extended this concept by analysing the effect of IL27 on various subsets of CD4+ T cells. IL27 markedly suppressed the differentiation of Th17 in all the three T-cell populations (CD4+, CD4+CD25−, CD4+CD25+) under optimal conditions for Th17 induction in vitro. However, IL27 had no detectable effect on Foxp3+ regulatory T cells in vitro and in vivo (fig 5 and data not shown). In disease models where IL27 had been found to suppress the inflammation,17 18 it is likely that Th17 plays a pivotal role and that the influence of IL27 was involved at the onset of the disease, such as IL27Rko or IL27ko. In contrast, where the effect of IL27 was found to exacerbate disease,15 16 it is most likely that either Th17 was not directly involved or that IL27 was administered at a later stage of disease development. In most of these disease-exacerbating cases, the pathogenesis may be mediated by IL6 or Th1 cytokines such as IFNγ which can be effectively induced by IL27.12–14 The fact that IL27 could have the opposite effect on the inflammatory process depending on the timing of administration may constrain its potential use as an effective therapeutic agent against inflammatory diseases.
We have recently reported that IL35, a member of the IL12 family which also includes IL27, is also an anti-inflammatory cytokine.27 IL35 is a heterodimer which shares EBI3 with IL27, and p35 with IL12. IL35 also strongly attenuated CIA by suppressing Th17 differentiation in vitro and in vivo. There are, however, important differences between IL27 and IL35. While IL35 inhibited the induction of Treg cells, IL27 did not. Furthermore, IL35 suppressed established CIA, IL27 only inhibited CIA when administered at the onset of the disease.
The mechanism by which IL27 influences Th17 has been extensively investigated. IL27 inhibits the generation of Th17 through a STAT1-dependent pathway,17 18 28 although a partial involvement of STAT3 has also been implicated.19 The inductive effect of IL27 on IFNγ may be through its induction of T-bet29 and inhibition of GATA-3,30 the master switches of Th1 and Th2, respectively. Other potential pathways involved in the regulatory role of IL27 on cytokine production are currently being explored. The receptor and the signalling pathway of IL35 are unknown.
It is of interest to note that IL27 is detected at significant levels in the joints of patients with RA (fig 1). It may be that the level of IL27 present was insufficient to suppress the Th17-driven inflammation, or that the late onset of IL27 synthesis was unable to downregulate mature Th17 and established disease.
In conclusion, our results demonstrate that IL27 could be a potential therapeutic agent against arthritic diseases at the onset of the disease but may not be effective against established arthritis.
Acknowledgments
We thank Mr Rod Ferrier and Mr James Reilly for preparation of the histological slides.
REFERENCES
Footnotes
Funding: This work was supported by the Wellcome Trust, Medical Research Council, UK; Arthritis Research Campaign, UK; the Chief Scientist’s Office, Scotland and the European Union. WN was also supported by the Committee of Scientific Research of Poland (grants 4PO 5BO 1319 and 2PO 5BO 8527).
Competing interests: None.
Ethics approval: Approved by local ethical committees.