Article Text

Abstract
Background: Gene therapy of the joint has great potential as a new therapeutic approach for the treatment of rheumatoid arthritis (RA). The vector chosen is of crucial importance for clinical success.
Objective: To investigate the tropism and transduction efficiency in arthritic joints in vivo, and in synovial cells in vitro, using five different serotypes of recombinant adeno-associated virus (rAAV) encoding β-galactosidase or green fluorescent protein genes.
Methods: rAAV was injected into the ankle joints of rats with adjuvant arthritis after the onset of disease. Synovial tissue was examined at different time points for β-galactosidase protein and gene expression by in situ staining and polymerase chain reaction (PCR) analysis, respectively. In addition, the ability of rAAV to transduce primary human fibroblast-like synoviocytes from patients with RA was investigated in vitro.
Results: Intra-articular injection of the rAAV5 serotype resulted in the highest synovial transduction, followed by much lower expression using rAAV2. Expression of the transgene was already detectable 7 days after injection and lasted for at least 4 weeks. Only background staining was seen for serotypes 1, 3, and 4. Importantly, there was a minimal humoral immune response to rAAV5 compared with rAAV2. Additionally, it was found that both rAAV2 and rAAV5 can efficiently transduce human fibroblast-like synoviocytes obtained from patients with RA.
Conclusion: Intra-articular rAAV mediated gene therapy in RA might be improved by using rAAV5 rather than other serotypes.
- AIA, adjuvant induced arthritis
- CMV, cytomegalovirus
- FLS, fibroblast-like synoviocytes
- GC, genomic copies
- gDNA, genomic DNA
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- GFP, green fluorescent protein
- IOD, integrated optical density
- MOD, mean optical density
- PBS, phosphate buffered saline
- PCR, poymerase chain reaction
- RA, rheumatoid arthritis
- rAAV, recombinant adeno-associated virus
- adeno-associated virus
- serotypes
- rheumatoid arthritis
- synovium
- gene therapy
Statistics from Altmetric.com
- AIA, adjuvant induced arthritis
- CMV, cytomegalovirus
- FLS, fibroblast-like synoviocytes
- GC, genomic copies
- gDNA, genomic DNA
- GAPDH, glyceraldehyde-3-phosphate dehydrogenase
- GFP, green fluorescent protein
- IOD, integrated optical density
- MOD, mean optical density
- PBS, phosphate buffered saline
- PCR, poymerase chain reaction
- RA, rheumatoid arthritis
- rAAV, recombinant adeno-associated virus
Rheumatoid arthritis (RA) is a chronic inflammatory disease characterised by persistent joint swelling and progressive destruction of cartilage and bone. Biological agents targeting cytokines, like tumour necrosis factor α or interleukin 1, and their receptors have proved useful as specific treatments for RA.1–,4 However, the need for continuous treatment with multiple injections of a recombinant protein, the relatively high systemic doses necessary to achieve constant therapeutic levels in the joints, and the reported side effects hamper this therapeutic approach.
An alternative approach might be to use intra-articular gene therapy to deliver therapeutic genes directly to the synovium in order to have the protein synthesised at the site of inflammation. Synoviocytes can be readily accessed by intra-articular injection, making them good targets for gene delivery. They are resident cells in the human synovium and have a low mitotic rate.5 Therefore, they are likely to express transduced genes for a considerable length of time.
Viral mediated gene transfer is currently the most efficient system for delivering therapeutic proteins in vivo. Adeno-associated virus (AAV) especially holds great promise as a new vector and appears to be safe.6,7 AAV is a single stranded DNA virus that does not induce a significant immune response and is not associated with disease in humans. The ability of AAV to mediate expression of therapeutic genes has been well established in several animal models of arthritis.8,9,10,11,12 Recombinant AAV2 (rAAV2) has been validated for gene therapy in human clinical trials in cystic fibrosis, α1 antitrypsin deficiency, and haemophilia.13–,15 rAAV vectors are particularly useful in targeting slowly dividing cells and in the treatment of chronic disease because of their potential for site-specific integration into the host genome or formation of stable episomal DNA, both of which result in long term gene expression.16,17
All studies reported so far have been performed using rAAV serotype 2. However, rAAV2 vectors are rather inefficient in transducing some cell types.18–,20 In addition, the prevalence of antibodies against this serotype is high in the human population.6 Many people carry antibodies that can neutralise infection of cells with rAAV2 in vitro, and it is widely assumed that this is also relevant in vivo.21,22 At this moment at least eight naturally occurring serotypes of AAV have been isolated, cloned, and sequenced (AAV1 to AAV8). Although these serotypes are very similar in genetic structure and biological properties, the capsid genes show as little as 50–60% homology in DNA sequence, resulting in viral shells that have a different amino acid composition.6,23 The use of non-type 2 AAV capsids to deliver transgenes to target cells might offer two important advantages over conventional AAV2 vectors: (a) an overall broader host range, and (b) an escape from anti-AAV2 immune responses, allowing vector (re-) administration despite existing immunity against AAV2.
In an effort to compare the role of serotype-specific virion shells on synovial transduction, we cloned the serotype capsid encoding domains into a common vector backbone containing AAV2 replication genes. This strategy allows the packaging of AAV2 inverted terminal repeat vectors into specific virions of each serotype, resulting in hybrid serotype vectors differing only in their capsids. In the study presented here we compared the transduction efficiency of five different recombinant AAV serotypes (rAAV1 to rAAV5) encoding the E coli derived reporter gene, β-galactosidase (β-Gal), to transduce arthritic synovium in vivo. In addition, we examined the transduction efficacy of rAAV containing the enhanced green fluorescent protein (GFP) gene in primary human RA fibroblast-like synoviocytes (FLS).
METHODS
Production of rAAV
All rAAV constructs were derived from AAV2 and driven by the cytomegalovirus (CMV) immediate early promoter. rAAV was produced by co-transfection of 293 HEK cells with 150 μg packaging plasmid (pDG for AAV224 and pDP1, pDP3, pDP4, and pDP5 for AAV1, AAV3, AAV4, and AAV5,25 respectively) and 50 μg vector plasmid containing the LacZ or the GFP gene with the CMV promoter and the bovine growth hormone polyA region between the AAV2 inverted terminal repeat vectors, by the calcium phosphate method. Plasmids were purified with Qiafilter plasmid Giga Kits (Qiagen, Alameda, USA).
Seventy two hours after transfection, the cells were lysed using benzonase (Merck, Whitehouse Station, USA) to a final concentration of 75 U/ml. Crude cell lysates were further purified with iodixanol gradients as described earlier.26 The gradient layer containing the virus was extracted, diluted 10 times with phosphate buffered saline (PBS) and concentrated to about 2 ml with centricon devices (YM-100, Millipore Corporation, Billerica, USA). Stock viral titres ranging between 1011 and 1012 genomic copies (GC)/ml in saline were reached.
Detection of viral genomic copies (GC) by quantitative polymerase chain reaction (Q-PCR)
To determine the viral titre as GC (full viral particles), rAAV samples were first diluted 10-fold in PBS. Samples were incubated with 10 μl of MagneSil BLUE suspension (Promega), and the viral DNA was isolated using the MagnaBot 96 Magnetic Separation Device (Promega) according to the supplier’s instructions.
Dilutions of the purified viral DNA or the genomic DNA (gDNA) isolated from the joints and organs were added to the PCR mix containing 0.5 μmol/l of the CMV forward primer (5′-AATGGGCGGTAGGCGTGTA-3′) (Invitrogen), 0.5 μmol/l of the CMV reverse primer (5′-AGGCGATCTGACGGTTCACTAA-3′) (Invitrogen), and SYBR green PCR master mix buffer (Applied Biosystems, Foster City, USA). PCR reactions were performed using the Abi prism SDS7000 sequence detection system (Applied Biosystems, Foster City, USA).
Animals
Pathogen-free male Lewis rats (150–200 g) were obtained from Harlan Sprague Dawley Inc (Horst, The Netherlands) and were maintained in our central animal facility. The ethical animal care and use committee of the University of Amsterdam approved all experiments.
Local gene transfer
All rats were immunised at the base of the tail with 1 mg of Mycobacterium tuberculosis H37RA (Difco, Detroit, USA) in 0.1 ml mineral oil on day 0.27 Paw swelling was usually seen by days 10–12 and measured daily by water displacement plethysmometry. At day 12, the skin was disinfected with ethanol and 6.1×1010 GC rAAV1–5 containing the gene encoding LacZ (further referred to as rAAV1–5) was injected anterolaterally into the right ankle joint in a total volume of 50 μl saline using a 31-gauge needle on a glass syringe.28 Adenovirus containing the LacZ gene (adjusted to 6.1×1010 GC/animal) served as a positive control and diluent was used as negative control. Rats injected with rAAV and control rats were killed 2 weeks after intra-articular injection by CO2 inhalation, whereas adenovirus-injected rats were killed 2 days after injection (n = 6/group). Serum samples were taken from the vena cava. Hind paws, spleen, heart, lung, liver, and kidney were collected.
To investigate the transgene expression at different time points, a second experiment was performed using a different batch of rAAV. Animals received an intra-articular injection of 1.14×1010 GC rAAV2 or rAAV5 and were killed as described 1, 2, 3, and 4 weeks after injection (n = 3/group). Serum samples were obtained from all groups by tail bleeding before rAAV injection and by vena cava punction as the animal was killed.
Detection of β-galactosidase activity
Joints were fixed in 4% formalin, decalcified using EDTA, and snap frozen in liquid nitrogen. Sections (10 μm) were cut on a cryostat and mounted on glass slides (Star Frost adhesive slides; Knittelgläser, Germany). Subsequently, detection of β-Gal activity was performed by X-Gal staining as described previously.28 After washing with PBS, sections were counterstained with nuclear red. The sections were evaluated by digital image analysis.
Digital image analysis
Six randomly selected fields within each section were chosen for digitising the positive signal. These images were acquired on an Olympus microscope (Olympus, Tokyo, Japan), captured using a Charged Coupled Device video camera (Sony, Tokyo, Japan), and digitised with a PV100 multimedia 16 bit colour video digitiser card. In the resultant colour images the area of positive staining and the mean optical density (MOD) were measured by a macro program as described earlier.29,30 The MOD is proportional to the cellular concentration of protein. The integrated optical density (IOD) is equal to the MOD multiplied by the area of positive staining.
PCR detection of LacZ
Ankle joints (trimmed of skin) and organs were snap frozen in liquid nitrogen, pulverised using a mortar, and homogenised in TRIzol Reagent (100 mg/ml; GibcoBRL Life Technologies, Gaithersburg, USA) using a tissue homogeniser. Total RNA was isolated from the aqueous phase and gDNA was extracted from the phenol-chloroform phase according to the manufacturer’s instructions. gDNA was stored for quantitative PCR analysis. RNA was dissolved in DEPC-water and cDNA was synthesised from 1 μg RNA using Superscript II RT (Invitrogen) according to the manufacturer’s instructions.
For PCR, 10 μl of cDNA solution was amplified using 25 μl of AccuPrime SuperMix I (Invitrogen), 215 mmol of the LacZ forward primer (5′-GCATCGAGCTGGGTAATAAGCGTTGGCAAT-3′), and 215 mmol of the LacZ reverse primer (5′-GACACCAGACCAACTGGTAATGGTAGCGAC-3′) in a total volume of 50 μl. Rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control (forward: 5′-CGGTGTCAACGGATTTGGC-3′, reverse: 5′-CCATGCCAGTGAGCTTCCC-3′). Amplification was then performed in a thermocycler (MJ Research, Inc, Waltham, USA) as follows: 3 minutes at 95°C, followed by 35 cycles at 94°C for 1 minute, 58°C for 90 seconds, and 72°C for 1 minute, respectively, followed by a final extension phase at 72°C for 10 minutes. The PCR products were analysed by standard agarose gel electrophoresis on a 0.9% agarose gel containing ethidium bromide for ultraviolet assisted visualisation of the 622 bp product. For semiquantitative analysis, the gel was digitised and the signal was corrected for background and area. Thereafter, the LacZ/GAPDH ratio was determined using Quantity One software (Biorad, Hercules, USA).
Formation of anti-rAAV neutralising antibodies in serum
Neutralising antibody titres were analysed by assessing the ability of serum antibodies to inhibit the transduction of rAAV into COS cells. Various dilutions of serum (1:200 to 1:51 200) were preincubated with rAAV at 37°C for 1 hour and then added to 80% confluent COS cells. Thereafter, cell cultures were incubated with rAAV in the presence of serum for 20 hours, and LacZ expression was measured by β-Gal staining as described earlier. The antibody titre was represented by the highest dilution that did not inhibit β-Gal expression compared with cells incubated with rAAV alone.
In vitro transduction of human FLS
Small-bore arthroscopy (2.7 mm arthroscope, Storz, Tuttlingen, Germany) was performed under local anaesthesia in three patients with established RA.31,32
The biopsy specimens were enzymatically dispersed and cultured as described previously33 and the FLS obtained were plated on six-well dishes (Flacon, Bedford, USA) at 1×105/well. After incubation for 15 hours, 1×108 GC of rAAV1–5 containing the gene for GFP was added to each well in medium containing 10% fetal calf serum. The cells were cultured for 72 hours and marker gene expression was evaluated by fluorescent microscopy and flow cytometric analysis. Three independent primary FLS cell lines from three patients were used for these experiments.
Flow cytometry
Transduced FLS were trypsinised, resuspended in Dulbecco’s modified Eagle’s medium containing 10% fetal calf serum, and washed in PBS containing 0.01% (wt/vol) NaN3 and 0.5% (wt/vol) bovine serum albumin. Analysis of GFP gene expression was performed using a FACS Calibur flow cytometer and CellQuest software (BD Biosciences, Franklin Lakes, USA).
Statistics
Where applicable, differences in means between groups were determined by Kruskal-Wallis test, followed by a Mann-Whitney U rank sum test. p<0.05 was considered significant. All analyses were done using SPSS version 11.5 (SPSS, Chicago, USA).
RESULTS
Comparative efficiency of five rAAV serotypes
To compare the transduction efficiency of rAAV serotypes in the joints, rAAV1–5 were injected into the right ankle joints of rats (n = 6/group) on day 12 after adjuvant immunisation. Joints were harvested 2 weeks after injection and stained in situ to measure β-Gal expression. Staining was quantified by digital image analysis. The strongest expression of β-Gal in synovial tissue was detected in arthritic joints injected with rAAV5, followed by a much lower expression using rAAV2 (fig 1B⇓). Strikingly, rAAV transduction resulted in a greater penetration into the synovial tissue than with adenovirus, where β-Gal expression was limited to the intimal lining layer. No staining could be seen in the control joints (fig 1A⇓). No expression above background staining was seen for serotypes 1, 3, and 4. These findings were confirmed by quantitative PCR. In gDNA isolated from crushed joints, the highest number of viral GC was detected using rAAV5 (1.9×105 GC/μg gDNA) (fig 2⇓). Semiquantitative PCR showed the presence of LacZ mRNA in all injected joints, but not in spleen, liver, heart, lung, kidney, or uninjected joints (data not shown).

β-Gal expression in rat synovial tissue 2 weeks after intra-articular injection of different rAAV serotypes. Joints were snap frozen and sections were cut and stained in situ for β-Gal activity and counterstained with nuclear red. Representative cryosections of right ankle joints of rats with adjuvant induced arthritis (AIA), injected with 6.1×1010 GC rAAV, Ad.LacZ as a positive control or saline as a negative control are shown (magnification ×100, ×400). The lining layer is indicated by arrows (A). Tissue sections were quantified for β-Gal expression by computer assisted digital image analysis (B). Values are expressed as mean (SD) cumulative IOD/mm2. *p<0.05 as compared with the control group, Mann-Whitney U test.

Viral genomic copies in the right ankle joints of rats injected with different rAAV serotypes. Two weeks after intra-articular rAAV injection, gDNA was isolated from crushed ankle joints and analysed for viral GC by quantitative PCR. Specific primers against the CMV promoter of the transgene were used. No viral DNA was found in control or left joints. Values are expressed as mean (SD) log GC/μg DNA. *p<0.05.
Duration of transgene expression
To monitor the transgene expression over time, rAAV2 or rAAV5 were injected into arthritic joints of rats and killed 1, 2, 3, or 4 weeks after injection of the vector. For both serotypes transgene expression was detectable 1 week after injection with marked expression for up to 4 weeks, but displayed different expression kinetics. rAAV5 resulted in earlier β-Gal expression than rAAV2, with higher levels at all time points (fig 3⇓). This was confirmed by quantitative PCR analysis. The number of GC detected in the joints after rAAV5 injection was higher at all time points than with rAAV2 (table 1⇓).
Detection of viral genomic copies in injected joints after intra-articular injection of rAAV2 or rAAV5
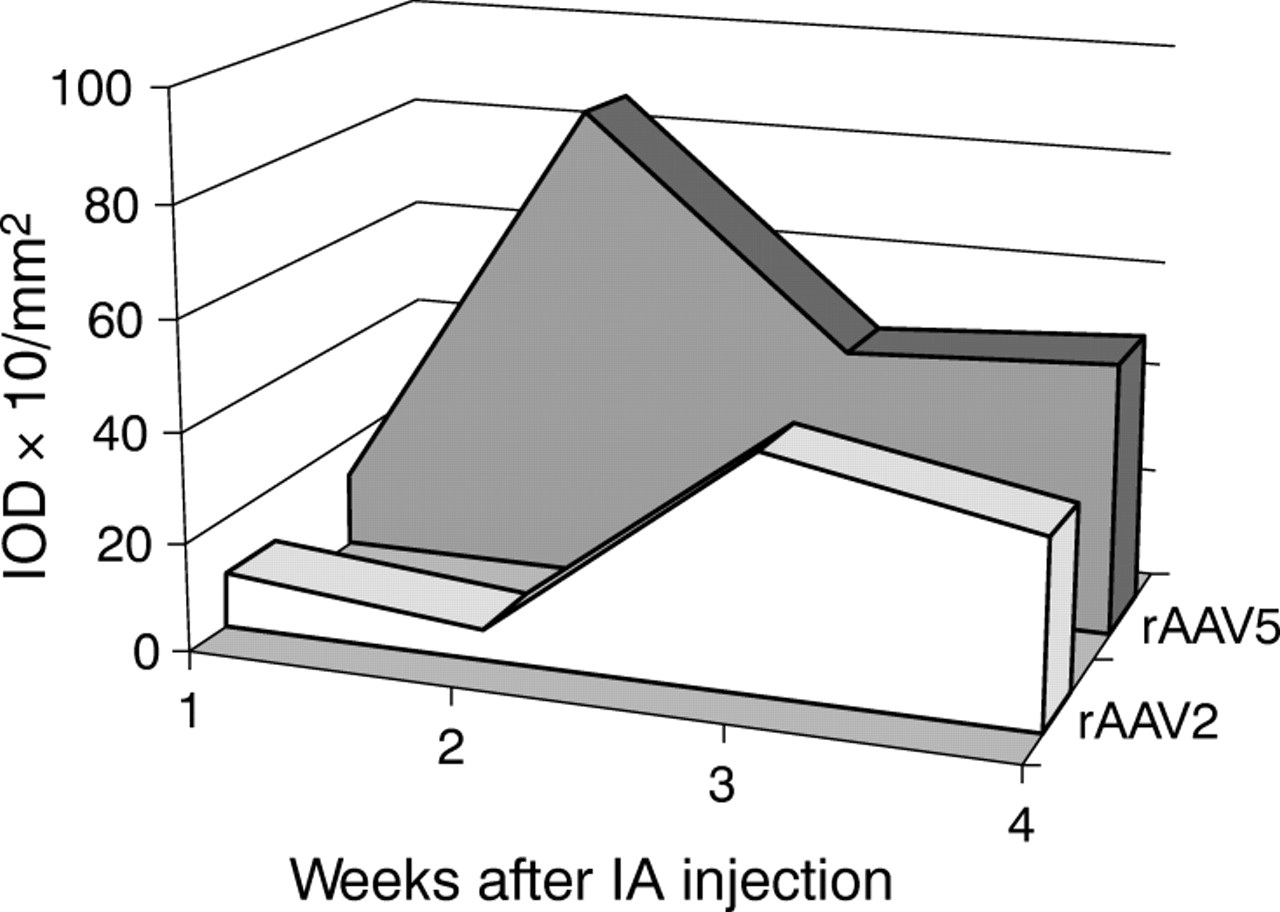
β-Gal expression in rat synovial tissue 1, 2, 3, and 4 weeks after intra-articular (IA) injection of 1.14×1010 GC rAAV serotype 2 or 5, quantified by digital image analysis. Injected joints were snap frozen and cryosections were stained in situ for β-Gal activity. Staining was evaluated by computer assisted analysis and expressed as IOD×10/mm2.
To test if this increased amount of viral GC also results in an increased transcription of the transgene, PCR analysis was carried out using specific primers for LacZ. Figure 4⇓ demonstrates that in rAAV2 as well as in rAAV5 injected animals, high levels of LacZ mRNA were found after 1 week. This rapidly declined in the rAAV2 injected animals, whereas in the rAAV5 injected animals the level of LacZ mRNA remained about 50% of the levels found after 1 week.

LacZ mRNA in rat ankle joints after intra-articular injection of rAAV2 and rAAV5. Total mRNA was extracted from crushed ankle joints and cDNA was synthesised. PCR was performed using specific primers for the LacZ gene. GAPDH was used as internal control. To correct for differences in GAPDH mRNA, LacZ/GAPDH ratios are displayed as measured by computer assisted gel quantification. The uninjected left joints as well as the control joints did not show LacZ expression.
A limited (n = 3/group) biodistribution study was performed, and no viral DNA or LacZ mRNA was found in the uninjected left joints at any time point. A relatively low amount of GC was found only in the kidney and liver after 1 week, but not at later time points, suggesting minimal, temporary dissemination of the vector (data not shown). In the spleen, liver, heart, lung, kidney, and lymph nodes, no LacZ mRNA was detected at any time point (data not shown).
Formation of rAAV antibodies
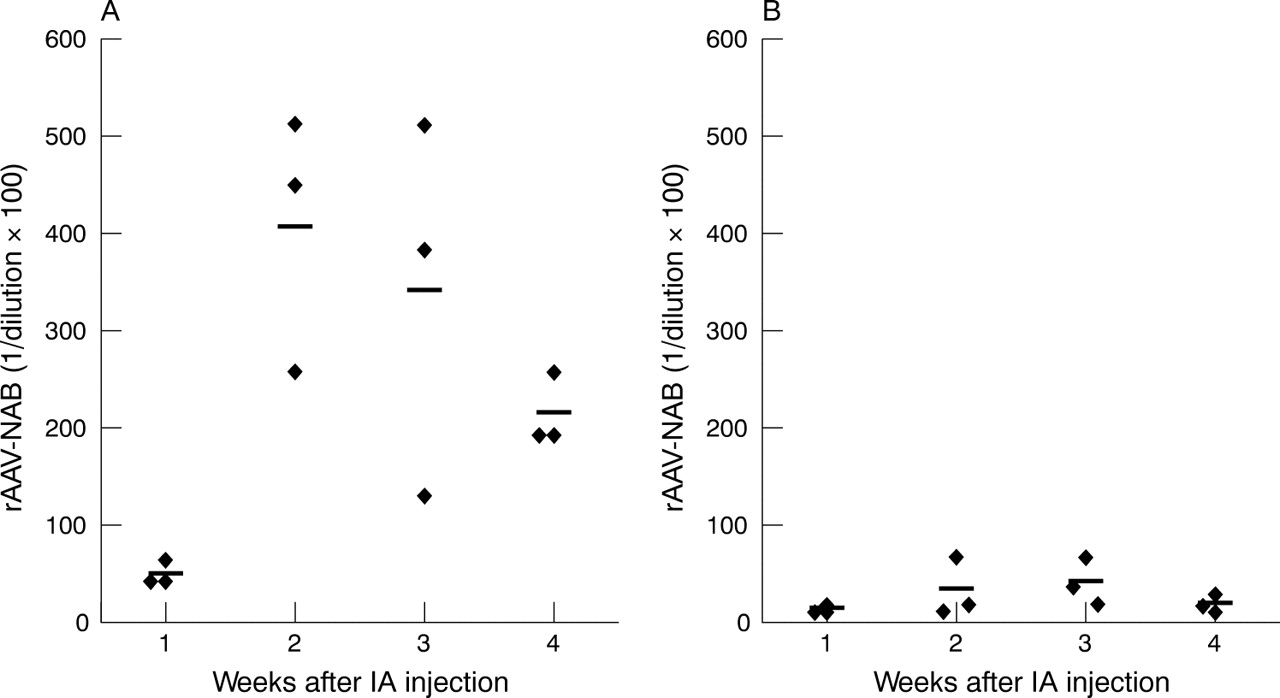
To detect a possible humoral immune response against the rAAV capsid proteins after intra-articular injection, we performed a neutralising antibody assay. The presence of neutralising antibodies before and after rAAV injection was determined in the serum of rats injected with rAAV2 and rAAV5.
Before injection, no antibodies were found in any of the samples. One week after injection, neutralising antibodies were detected, peaking at 2 weeks and slowly decreasing after 3 weeks. Although this tendency was seen for both serotypes, rAAV2 injection obviously induced higher neutralising antibody titres in serum than rAAV5, which induced titres only slightly above background (fig 5⇓). Importantly, no cross reactivity was found for the two serotypes (data not shown).

Neutralising antibody (NAB) titres in serum after intra-articular (IA) injection of rAAV2 or rAAV5. Arthritic rats were injected with 1.14×1010 GC rAAV2 (A) or rAAV5 (B) into the right ankle joints. Serum samples were obtained 1, 2, 3, and 4 weeks after injection. Titres were calculated as the highest dilution which shows no decrease in number of β-Gal positive cells compared with wells incubated with rAAV.LacZ alone.
Transduction of human FLS
Having shown that rAAV2 and rAAV5 can transduce rat synovium, we investigated the potential of both serotypes to transduce primary human FLS obtained from patients with RA. For this purpose we used rAAV vectors expressing GFP. FLS were incubated with the different rAAV serotypes and transgene expression was visualised after 72 hours by fluorescent microscopy (fig 6⇓). Both serotypes could transduce human FLS with high efficiency, although no clear difference was seen between rAAV2 and rAAV5. To quantify the percentage of GFP expressing cells, FLS were transduced using five rAAV serotypes as described and GFP expression was evaluated by FACS analysis (fig 7⇓). Both rAAV2 and rAAV5 resulted in significant numbers of GFP expressing cells (45% and 40%, respectively) compared with control (fig 7A⇓). Compared with rAAV1, 3, and 4, rAAV2 and rAAV5 transduced significantly more FLS (fig 7B⇓), p<0.05.

Transduction of FLS with rAAV in vitro. Human FLS isolated from synovial biopsy specimens from patients with RA were incubated with rAAV2.GFP or rAAV5.GFP. After 48 hours of incubation, the cells were fixed and transgene expression was determined by fluorescent microscopy. Representative pictures of three independent experiments using cells from three different patients are shown. Left: fluorescent microscopy, right: phase contrast microscopy.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Flow cytometric analysis of GFP expression in rAAV transduced FLS. Human FLS were incubated with rAAV2.GFP or rAAV5.GFP at a multiplicity of infection of 100. After 72 hours, the cells were washed and analysed for GFP expression by FACS. Flu-1 plotted against FSC-H showed 45% (left panel, rAAV2) and 40% (middle panel, rAAV5) GFP positive cells, compared with 3% in control cells (right panel), p<0.05 (A). Differences between rAAV2 and rAAV5 were not significantly different. In a histogram comparing GFP expression of control cells with all five rAAV serotypes, only differences between rAAV2 or rAAV5 and control or rAAV1, 3, or 4 proved significant using the Mann-Whitney U test (B) (p<0.05). Representative histogram of three independent experiments is shown.
DISCUSSION
rAAV vectors have gained much attention owing to their ability to mediate efficient transduction of both dividing and non-dividing cells and their ability to induce long term gene expression in the absence of toxicity. Over the past decade, rAAV vectors derived predominantly from serotype 2 were generated and investigated in vivo. The recent discovery of AAVs other than serotype 2 has enabled the generation of pseudotyped rAAV vectors. Until now, no studies of gene transfer to the inflamed joint have been carried out with non-serotype-2 vectors. Therefore, we compared the transduction efficiencies of titre-matched rAAVs derived from serotypes 1–5, in synovial tissue of rats with AIA. The efficiency of gene expression was evaluated by enzymatic staining and by determining the amount of GC present in the joints. In this comparative study we found that direct injection of rAAV5 into the ankle joints of rats with AIA resulted in the highest synovial transduction as demonstrated by the expression level of β-Gal, followed by a much lower expression using rAAV2. Analysis of the gDNA isolated from the joints confirmed these results.
In a second experiment we investigated the transgene expression of rAAV2 and rAAV5 at different time points. Both serotypes demonstrated marked expression over time, which was already present 1 week after injection. Of importance, β-Gal protein expression using rAAV5 was higher at all time points, confirmed by higher levels of mRNA encoding β-Gal. The fact that the gDNA profile did not parallel β-Gal expression could be explained, in part, by the relative stability of the β-Gal protein, resulting in prolonged presence of the protein. Although β-Gal is a foreign protein and possibly immunogenic, we did not observe differences in paw swelling or cellularity between rAAV injected and control animals (data not shown).
rAAV2 has previously been shown to mediate long term gene transfer in a number of tissues, including synovial tissue.8,9,10,34,35,36 The feasibility of direct in vivo gene transfer to rat and murine arthritic joints has also been demonstrated. In most of the studies, rAAV2 genome persisted within joints for at least 100–200 days.8,11,36 One study reported long term expression after rAAV2 gene transfer to the murine synovium for up to 7 months.9 The expression pattern of rAAV2 after intra-articular injection in mice has been variable, varying from synovial lining cells36 to chondrocytes,11 and myocytes.8,9 In our study, morphological analysis suggested that the cells expressing β-Gal were mainly synovial fibroblasts. Because these cells are thought to stay in the synovial tissue for long periods of time, this would allow long term transgene expression in the joint, with minimal spreading of transduced cells to peripheral tissues. In addition to the intimal lining layer, cells in the synovial sublining were transduced as well, maximising the potential of therapeutic protein production throughout the synovial tissue.
Serotype-specific affinities for cell surface attachment and internalisation receptors may, at least in part, account for the difference in the level of transgene expression between the different serotypes. Heparan sulphate proteoglycans,37 integrins,38 and fibroblast growth factor receptor 139 are important receptors for rAAV2. Recently, N-linked sialic acid40 and platelet derived growth factor receptor41 were identified as co-receptors for AAV5, and their expression in vivo correlates well with the transduction profile of rAAV5.41 In addition, trafficking of rAAV5 to the nucleus or its uncoating might be more efficient than for the other serotypes, explaining the higher number of GC and gene expression observed with rAAV5.
Having shown the feasibility of direct in vivo gene transfer to rat synovium, we investigated the transduction efficiency of rAAV to human FLS derived from patients with RA, which is crucial for using this vector for local human gene therapy. FLS have the highest residence in the human synovium and have a low mitotic rate.5,42 Therefore, FLS are likely to express transduced genes for a considerable length of time, even if the transgene is located episomally. In contrast with rAAV1, 3, and 4, synovial fibroblasts isolated from the joints of patients with RA could be transduced with high efficiency in vitro using both rAAV2 and rAAV5. No significant differences were found between these two serotypes, suggesting that factors other than differences in affinity for cell surface receptors contribute to the different tropism seen in vivo. The transduction of human macrophages was most efficient using rAAV5, but did result in only 2% GFP positive cells (data not shown).
Some studies investigating the effect of repeated administration of rAAV indicate that an immune response generated after the first administration may prevent re-treatment.43 We found that local rAAV2 injection in the joint clearly induces more neutralising antibodies in rat serum than rAAV5, which induced neutralising antibody levels only slightly above background. No antibodies were detected before injection of the vectors. This indicates that a marked humoral immune response is mounted against rAAV2, while the capsid of rAAV5 is less immunogenic. Importantly, no cross reactivity was seen between antibodies raised against rAAV2 and rAAV5, suggesting that re-administration could be applied using different serotypes. It should be noted that an estimated 80% of the human population carry antibodies to the capsid proteins of wild-type AAV2, and 30–70% demonstrate the presence of neutralising anti-capsid antibodies.21,44 In contrast, neutralising antibodies against AAV5 are rare. Moreover, endogenous sequences of AAV5 suggesting previous infection with this serotype could not be detected in a large diverse group of people.45 Together, these observations are important for the choice of an rAAV serotype vector for clinical applications, favouring the use of rAAV5.
In summary, our results show that rAAV5 is more efficient in transducing rat synovial tissue in vivo, and both rAAV2 and rAAV5 can efficiently transduce human RA synoviocytes in vitro. In addition, we showed little to no humoral immune response to rAAV5 in rat AIA. Of the five rAAV serotypes tested, rAAV5 appears to be an excellent candidate vector for local gene therapy in patients with RA, allowing long term expression of the transgene limited to the synovial compartment.
Acknowledgments
We thank Dr Marc R Kok for his contributions and critical review of the manuscript.
REFERENCES
Footnotes
Published Online First 5 May 2005